Signální dráha TGF beta - TGF beta signaling pathway
The signální dráha transformujícího růstového faktoru beta (TGFB) podílí se na mnoha buněčných procesech v dospělém organismu i ve vývoji embryo počítaje v to růst buněk, diferenciace buněk, apoptóza, buněčná homeostáza a další buněčné funkce. Navzdory širokému spektru buněčných procesů, které signální dráha TGFβ reguluje, je proces relativně jednoduchý. Nadrodina TGFβ ligandy se vážou na receptor typu II, který rekrutuje a fosforyláty receptor typu I. Receptor typu I poté fosforyluje SMAD regulované receptorem (R-SMAD ), který nyní může vázat coSMAD SMAD4. Komplexy R-SMAD / coSMAD se hromadí v jádru, kde působí transkripční faktory a podílet se na regulaci exprese cílového genu.
Mechanismus
Ligandová vazba
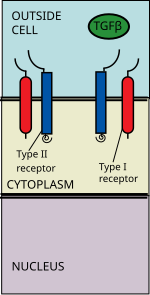
Nadrodina ligandů TGF beta zahrnuje: Kostní morfogenetické proteiny (BMP), Faktory růstu a diferenciace (GDF), Anti-Müllerian hormon (AMH), Activin, Nodal a TGFβ je[1] Signalizace začíná navázáním ligandu TGF beta nadrodiny na receptor TGF beta typu II. Receptor typu II je kináza receptoru serinu / threoninu, která katalyzuje the fosforylace receptoru typu I. Každá třída ligandu se váže na specifický receptor typu II.[2] U savců je známo sedm receptorů typu I a pět receptorů typu II.[3]
Existují tři aktiviny: Aktivin A, Aktivin B. a Activin AB. Aktiviny se účastní embryogeneze a osteogeneze. Mnoho také regulují hormony počítaje v to hypofýza, gonadal a hypotalamus hormony stejně jako inzulín. Jsou taky nervová buňka faktory přežití.
BMP se váží na kostní morfogenetický proteinový receptor typu-2 (BMPR2). Podílejí se na mnoha buněčných funkcích včetně osteogeneze, diferenciace buněk, specifikace přední / zadní osy, růst a homeostáza.
Rodina beta TGF zahrnuje: TGFβ1, TGFβ2, TGFβ3. Stejně jako BMP se beta-TGF účastní embryogeneze a buněčné diferenciace, ale také apoptózy a dalších funkcí. Oni se vážou TGF-beta receptor typu 2 (TGFBR2).
Nodal váže se na receptor aktivinu A, typ IIB ACVR2B. Poté může buď tvořit receptorový komplex s receptorem aktivinu A, typ IB (ACVR1B ) nebo s receptorem aktivinu A, typ IC (ACVR1C ).[3]
Když dojde k vazbě receptor-ligand lokálním působením, je to klasifikováno jako parakrinní signalizace.
Nábor a fosforylace receptorů

TGF beta ligand se váže na dimer receptoru typu II, který rekrutuje dimer receptoru typu I a tvoří s ligandem hetero-tetramerní komplex.[4] Tyto receptory jsou receptory serin / threonin kinázy. Mají cystein bohatý extracelulární doména, a transmembránová doména a a cytoplazmatický doména bohatá na serin / threonin. The GS doména receptoru typu I sestává ze série asi třiceti serin -glycin opakování.[5] Vazba ligandu rodiny TGF beta způsobuje rotaci receptorů tak, že jejich cytoplazmatické kinázové domény jsou uspořádány v katalyticky příznivé orientaci. Receptor typu II fosforyláty serin zbytky receptoru typu I, který aktivuje protein.
Fosforylace SMAD

Existuje pět receptorem regulovaných SMAD: SMAD1, SMAD2, SMAD3, SMAD5, a SMAD9 (někdy označované jako SMAD8). Existují v zásadě dvě intracelulární cesty R-SMAD. TGF beta, aktiviny, uzliny a některé GDF jsou zprostředkovány SMAD2 a SMAD3, zatímco BMP, AMH a několik GDF jsou zprostředkovány SMAD1, SMAD5 a SMAD9. Vazba R-SMAD na receptor typu I je zprostředkována proteinem obsahujícím zinek s dvojitým prstem FYVE. Dva takové proteiny, které zprostředkovávají cestu TGF beta, zahrnují SARA (kotva SMAD pro aktivaci receptoru) a HGS (substrát tyrosinkinázy regulovaný růstovým faktorem hepatocytů).
SARA je přítomna brzy endozom který tím, že klatrinem zprostředkovaná endocytóza, internalizuje receptorový komplex.[6] SARA rekrutuje a R-SMAD. SARA umožňuje vazbu R-SMAD na oblast L45 receptoru typu I.[7] SARA orientuje R-SMAD tak, aby na něm zůstal serinový zbytek C-konec čelí katalytické oblasti receptoru typu I. Receptor typu I. fosforyláty serinový zbytek R-SMAD. Fosforylace indukuje konformační změnu v MH2 doména R-SMAD a jeho následná disociace z receptorového komplexu a SARA.[8]
Vazba CoSMAD

Fosforylovaný RSMAD má vysokou afinitu k coSMAD (např. SMAD4 ) a tvoří komplex s jedním. Fosfátová skupina nepůsobí jako dokovací místo pro coSMAD, spíše fosforylace otevírá úsek aminokyseliny umožňující interakci.
Transkripce

Fosforylovaný komplex RSMAD / coSMAD vstupuje do jádra, kde se váže na promotory / kofaktory transkripce a způsobuje transkripci DNA.
Kostní morfogenetické proteiny způsobují transkripci mRNA zahrnutý do něčeho, zůčastnit se čeho osteogeneze, neurogeneze, a ventrální mezoderm Specifikace.
TGF betas způsobují transkripci mRNA zapojených do apoptóza, extracelulární matrix neogeneze a imunosuprese. Rovněž se podílí na G1 zatčení v buněčný cyklus.
Aktivin způsobuje transkripci mRNA zapojených do gonadal růst, diferenciace embryí a tvorba placenty.
Nodal způsobuje transkripci mRNA zapojených do specifikace levé a pravé osy, mezoderm a endoderm indukce.
Regulace dráhy
Signální dráha TGF beta je zapojena do široké škály buněčných procesů a následně je velmi silně regulována. Existuje celá řada mechanismů, kde je dráha modulována buď pozitivně, nebo negativně: Existují agonisté ligandů a R-SMAD; existují návnady receptory; a R-SMAD a receptory jsou ubikvitinovaný.
Agonisté / antagonisté ligandu
Oba chordin a člun ze dřeva jsou antagonisté BMP. Váží BMP a brání vazbě ligandu na receptor.[9] Bylo prokázáno, že Chordin a Noggin dorzalizují mezoderm. Oba se nacházejí v hřbetním rtu Xenopus a převést jinak pokožka specifikovaná tkáň do nervové tkáně (viz neurulace ). Noggin hraje klíčovou roli ve vzorcích chrupavek a kostí. Myši Noggin - / - mají přebytečnou chrupavku a chybí jim tvorba kloubů.[9]
Členové rodiny proteinů DAN také antagonizují členy rodiny TGF beta. Obsahují Cerberus, DAN, a Skřítek. Tyto proteiny obsahují devět konzervovaných cysteiny které mohou tvořit disulfidové můstky. Předpokládá se, že DAN antagonizuje GDF5, GDF6 a GDF7.
Follistatin inhibuje aktivin, na který se váže. Přímo to ovlivňuje folikuly stimulující hormon (FSH) sekrece. Follistatin se také podílí na rakovině prostaty, kde mu mutace v genu mohou bránit v působení na aktivin, který má antiproliferativní vlastnosti.[9]
Levičák je regulátor TGFβ a podílí se na vzorování os během embryogeneze. Je také členem nadrodiny proteinů TGF. Je asymetricky exprimován na levé straně myších embryí a následně hraje roli ve specifikaci zleva doprava. Levice působí tak, že brání fosforylaci R-SMAD. Činí tak prostřednictvím konstitutivně aktivního receptoru TGFp typu I a procesem po jeho aktivaci.[10]
Byli také identifikováni antagonisté na bázi drog, jako je SB431542,[11] který selektivně inhibuje ALK4, ALK5 a ALK7.
Regulace receptoru
The receptor transformujícího růstového faktoru 3 (TGFBR3) je dosud nejhojnější z TGF-β receptorů,[12] nemá žádnou známou signální doménu.[13] Může však sloužit ke zvýšení vazby ligandů TGF beta na receptory TGF beta typu II vazbou TGFp a jeho prezentací na TGFBR2. Jeden z navazujících cílů TGF β signalizace, GIPCse váže na svou PDZ doménu, což brání její proteosomální degradaci, která následně zvyšuje aktivitu TGFp. Může také sloužit jako inhibin coreceptor do AktivinRII.[9]
BMP a inhibitor vázaný na membránu aktivinu (BAMBI), má podobnou extracelulární doménu jako receptory typu I. Postrádá intracelulární doménu serin / threonin proteinkinázy, a proto je pseudoreceptor. Váže se na receptor typu I a brání jeho aktivaci. Slouží jako negativní regulátor signalizace TGF beta a může omezit expresi tgf-beta během embryogeneze. Pro svou expresi vyžaduje signalizaci BMP
FKBP12 váže GS oblast receptoru typu I a brání fosforylaci receptoru receptory typu II. Předpokládá se, že FKBP12 a jeho homology pomáhají zabránit aktivaci receptoru typu I v nepřítomnosti ligandů, protože vazba ligandu způsobuje jeho disociaci.
Regulace R-SMAD
Úloha inhibičních SMAD
Rodina SMAD doplňují další dva, SMAD inhibiční SMAD (I-SMADS), SMAD6 a SMAD7. Hrají klíčovou roli v regulaci TGF beta signalizace a účastní se negativní zpětné vazby. Stejně jako ostatní SMAD mají doménu MH1 a MH2. SMAD7 soutěží s jinými R-SMAD s receptorem typu I a zabraňuje jejich fosforylaci.[9][14] Nachází se v jádře a při aktivaci TGF beta receptoru se translokuje do cytoplazmy, kde se váže na receptor typu I. SMAD6 váže SMAD4 a brání tak navázání dalších R-SMAD na coSMAD. Úrovně I-SMAD se zvyšují se signalizací TGF beta, což naznačuje, že jsou cílovými cíli signalizace TGF-beta.
Ubikvitinace R-SMAD
E3 ubikvitinový protein ligázy SMURF1 a SMURF2 regulovat úrovně SMAD. Přijímají ubikvitin z konjugačního enzymu E2, kde přenášejí ubikvitin na RSMAD, což způsobuje jejich ubikvitinaci a následné proteosomální degradace. SMURF1 se váže na SMAD1 a SMAD5 zatímco SMURF2 se váže SMAD1, SMAD2, SMAD3, SMAD6 a SMAD7. To[je zapotřebí objasnění ] zvyšuje inhibiční účinek SMAD7 a snižuje transkripční aktivity SMAD2.
Souhrnná tabulka
TGF-β ligandy H. sapiens zvýrazněno šedě, z D.melanogaster ligandy v růžové barvě C.elegans žlutě.
| Ligand superrodiny TGF-p | Inhibitory ligandů | Receptor typu II | Receptor typu I. | R-SMAD | coSMAD | I-SMAD |
|---|---|---|---|---|---|---|
| Aktivin A | Follistatin | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| GDF1 | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | SMAD7 | |
| GDF11 | ACVR2B | ACVR1B (ALK4), TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | SMAD7 | |
| BMP2-8 | Člun ze dřeva, Chordin, DAN | BMPR2 | BMPR1A (ALK3), BMPR1B (ALK6) | SMAD1 SMAD5, SMAD8 | SMAD4 | SMAD6, SMAD7 |
| Nodal | Levičák | ACVR2B | ACVR1B (ALK4), ACVR1C (ALK7) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| TGFβs | LTBP1, THBS1, Decorin | TGFpRII | TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| Dpp | Pramice[nutná disambiguation ] | Tkv | Šílený | Medea | ||
| Šroub | Pramice[nutná disambiguation ] | Saxofon[nutná disambiguation ] | Šílený | Medea | ||
| myoglianin | Vtip[nutná disambiguation ] | Pavián[nutná disambiguation ] | dSmad2 | Medea | ||
| dActivin | Vtip[nutná disambiguation ], Pramice[nutná disambiguation ] | Pavián[nutná disambiguation ] | dSmad2 | Medea | ||
| Gbb | Vtip, Pramice[nutná disambiguation ] | Tkv, Saxofon[nutná disambiguation ] | Šílený | Medea | ||
| Daf-7 | Daf-4 | Daf-1 | Daf-8, Daf-14 | Daf-3 | ||
| Dbl-1 | Daf-4 | SMA-6 | SMA-2, SMA-3, SMA-4 | SMA-4 |
externí odkazy
- Kyoto Encyclopedia of Genes and Genomes -Signální dráha TGF beta mapa
- Netpath - Vybraný zdroj drah přenosu signálu u lidí
Reference
- ^ „Prosite Documentation PDOC00223“. Archivovány od originál dne 25. 05. 2011. Citováno 2006-07-01.
- ^ Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). Molekulární biologie buňky. New York, NY: Garland Science. ISBN 978-0-8153-3218-3.
- ^ A b Munir S, Xu G, Wu Y, Yang B, Lala PK, Peng C (červenec 2004). „Nodal a ALK7 inhibují proliferaci a indukují apoptózu v lidských trofoblastových buňkách“. The Journal of Biological Chemistry. 279 (30): 31277–86. doi:10,1074 / jbc.M400641200. PMID 15150278.
- ^ Wrana JL, Attisano L, Cárcamo J, Zentella A, Doody J, Laiho M a kol. (Prosinec 1992). „Signály TGF beta prostřednictvím komplexu heteromerních proteinových kinázových receptorů“. Buňka. 71 (6): 1003–14. doi:10.1016 / 0092-8674 (92) 90395-S. PMID 1333888.
- ^ „Položka Pfam TGF_beta_GS“. Citováno 2006-07-01.
- ^ Runyan CE, Schnaper HW, Poncelet AC (březen 2005). „Role internalizace v transformaci asociace Smad2 vyvolané růstovým faktorem beta1 s kotvou Smad pro aktivaci receptoru (SARA) a signalizací závislou na Smad2 v lidských mezangiálních buňkách“. The Journal of Biological Chemistry. 280 (9): 8300–8. doi:10,1074 / jbc.M407939200. PMID 15613484.
- ^ Moustakas A (září 2002). "Signální síť Smad". Journal of Cell Science. 115 (Pt 17): 3355–6. PMID 12154066.
- ^ Souchelnytskyi S, Rönnstrand L, Heldin CH, ten Dijke P (2001). "Fosforylace signálních proteinů Smad receptorovými serin / threonin kinázami". Protokoly proteinkinázy. Metody v molekulární biologii. 124. 107–20. doi:10.1385/1-59259-059-4:107. ISBN 1-59259-059-4. PMID 11100470.
- ^ A b C d E Massagué J, Chen YG (březen 2000). „Řízení signalizace TGF-beta“. Geny a vývoj. 14 (6): 627–44. PMID 10733523.
- ^ Ulloa L, Tabibzadeh S (červen 2001). „Lefty inhibuje receptorem regulovanou fosforylaci Smad vyvolanou aktivovaným receptorem transformujícího růstového faktoru-beta“. The Journal of Biological Chemistry. 276 (24): 21397–404. doi:10,1074 / jbc.M010783200. PMID 11278746.
- ^ Laping NJ, Grygielko E, Mathur A, Butter S, Bomberger J, Tweed C a kol. (Červenec 2002). „Inhibice extracelulární matrice indukované transformujícím růstovým faktorem (TGF) -beta1 novým inhibitorem aktivity kinázy receptoru TGF-beta typu I: SB-431542“. Molekulární farmakologie. 62 (1): 58–64. doi:10,1124 / mol. 62.1.58. PMID 12065755.
- ^ Blobe GC, Liu X, Fang SJ, How T, Lodish HF (říjen 2001). „Nový mechanismus pro regulaci signalizace transformujícího růstového faktoru beta (TGF-beta). Funkční modulace exprese receptoru TGF-beta typu III prostřednictvím interakce s proteinem domény PDZ, GIPC“. The Journal of Biological Chemistry. 276 (43): 39608–17. doi:10,1074 / jbc.M106831200. PMID 11546783.
- ^ Online Mendelian Inheritance in Man (OMIM): TRANSFORMACE RŮSTU FAKTOR-BETA RECEPTOR, TYP III; TGFBR3 - 600742
- ^ Itoh F, Asao H, Sugamura K, Heldin CH, ten Dijke P, Itoh S (srpen 2001). „Podpora signalizace kostního morfogenetického proteinu prostřednictvím negativní regulace inhibičních Smadů“. Časopis EMBO. 20 (15): 4132–42. doi:10.1093 / emboj / 20.15.4132. PMC 149146. PMID 11483516.