Decapentaplegic - Decapentaplegic
| Decapentaplegic | |||||||
|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||
| Organismus | |||||||
| Symbol | Dpp | ||||||
| UniProt | P07713 | ||||||
| |||||||
Decapentaplegic (Dpp) je klíč morfogen podílející se na vývoji ovocné mušky Drosophila melanogaster a je prvním validovaným secernovaným morfogenem.[1] Je známo, že je to nezbytné pro správné vytváření vzorů a vývoj na počátku Drosophila embryo a patnáct imaginární disky, což jsou tkáně, které se stanou končetinami a dalšími orgány a strukturami u dospělé mouchy. Rovněž bylo navrženo, že Dpp hraje roli v regulaci růstu a velikosti tkání. Mouchy s mutacemi v decapentaplegic nedokážou správně vytvořit tyto struktury, proto název (dekapenta-, patnáct, -plegický, paralýza). Dpp je homolog Drosophila obratlovců kostní morfogenetické proteiny (BMP), které jsou členy Nadrodina TGF-β, třída proteinů, které jsou často spojeny s jejich vlastní specifickou signální cestou. Studie Dpp u Drosophila vedly k lepšímu pochopení funkce a důležitosti jejich homologů u obratlovců, jako jsou lidé.
Funkce v Drosophila
Dpp je klasický morfogen, což znamená, že je přítomen v gradientu prostorové koncentrace v tkáních, kde se nachází, a jeho přítomnost jako gradientu mu dává funkční význam v tom, jak ovlivňuje vývoj. Nejvíce studovanými tkáněmi, ve kterých se Dpp nachází, jsou časné embryo a imaginární disky křídla, které později tvoří křídla mouchy. Během embryonálního vývoje je Dpp rovnoměrně exprimován na hřbetní straně embrya a vytváří ostrý koncentrační gradient.[2] V imaginálních discích je Dpp silně vyjádřen v úzkém pruhu buněk uprostřed disku, kde tkáň označuje hranici mezi přední a zadní stranou. Dpp difunduje z tohoto pruhu směrem k okrajům tkáně a vytváří gradient podle očekávání morfogenu. Ačkoli se však buňky v doméně Dpp v embryu neproliferují, buňky v disku imaginárního křídla se silně množí, což způsobuje růst tkáně.[1] I když je tvorba gradientu v časném embryu dobře známa, zůstává, jak se formuje gradient Dpp morfogenu v imaginárním disku křídla, kontroverzní.
Role a formace v embryonálním vývoji
Na začátku blastoderm fázi je signalizace Dpp rovnoměrná a nízká podél hřbetní strany. Ostrý signální profil se objevuje na hřbetní středové čáře embrya během cellularizace, přičemž vysoké hladiny Dpp specifikují extraembryonální amnioserosa a nízké úrovně specifikují dorzální ektoderm.[3] Signalizace DPP také zahrnuje a Pozitivní zpětná vazba mechanismus, který podporuje budoucí vazbu Dpp.[4] Morfogenní gradient v embryích je stanoven známým aktivním transportním mechanismem.[5] Tvorba gradientu závisí na inhibitorech BMP Krátká gastrulace (Sog) a Twisted gastrulace (Tsg) a další extracelulární proteiny jako Tolloid (Tld) a Screw (Scw).[6][7][8] Sog je produkován ve ventrálně-laterální oblasti embrya (kolmo k gradientu Dpp) a tvoří gradient inhibující BMP, který zabraňuje vazbě Dpp na jeho receptor.[9] Sog a Tsg tvoří komplex s Dpp a jsou aktivně transportovány směrem k hřbetní střední linii (uprostřed embrya), sledující koncentrační gradient Sog. Tld, a metaloproteáza, uvolňuje Dpp z komplexu zprostředkováním zpracování Sog, aktivací signalizace Dpp ve střední linii.[10] Po gastrulace embrya, gradient Dpp indukuje srdeční a viscerální mezoderm formace.[11]
Signalizační cesta
Dpp, stejně jako jeho homology obratlovců, je signální molekula. V Drosophile je receptor pro Dpp tvořen dvěma proteiny, Thickveins (Tkv) a Punt.[12] Stejně jako samotný Dpp jsou Tkv a Punt velmi podobné homologům u jiných druhů. Když buňka přijme signál Dpp, receptory jsou schopné aktivovat nazývaný intracelulární protein matky proti Dpp (šílený) fosforylací. Počáteční objev šílenců v Drosophile připravil půdu pro pozdější experimenty, které identifikovaly respondenta na signalizaci TGF-β u obratlovců, tzv. SMAD.[13] Activated Mad je schopen vázat se na DNA a chovat se jako transkripční faktor ovlivnit expresi různých genů v reakci na signalizaci Dpp. Geny aktivované signalizací Dpp zahrnují optomotorickou slepotu (omb) a spalt a aktivita těchto genů se často používá jako indikátory signalizace Dpp v experimentech. Další gen s komplikovanější regulační interakcí s Dpp je brinker. Brinker je transkripční faktor, který potlačuje aktivační cíle Dpp, takže aby bylo možné tyto geny zapnout, musí Dpp potlačit Brinker a také aktivovat ostatní cíle.[14]
Role v Imaginal Wing Disc
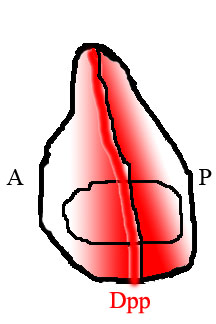
V křídle mouchy jsou zadní a přední poloviny tkáně osídleny různými druhy buněk, které exprimují různé geny. Buňky v zadní, ale ne v přední části, exprimují transkripční faktor Engrailed (En). Jedním z genů aktivovaných En je ježek (hh), signalizační faktor. Hedgehog signalizace dává pokyn sousedním buňkám k expresi Dpp, ale Dpp výraz je také potlačen En. Výsledkem je, že Dpp je produkován pouze v úzkém pruhu buněk bezprostředně sousedících, ale ne v zadní polovině tkáně.[15] Dpp produkovaný na tomto předním / zadním okraji pak difunduje ven k okrajům tkáně a vytváří gradient prostorové koncentrace.
Čtením jejich polohy podél gradientu Dpp jsou buňky v křídle schopny určit svou polohu vzhledem k přední / zadní hranici a podle toho se chovají a vyvíjejí.
Je možné, že tkáně nevytváří ve skutečnosti difúze a gradient Dpp, nýbrž buňky, které přijímají signál Dpp, instruují své sousedy, co mají být, a tyto buňky zase signalizují své sousedy v kaskádě přes tkáň. Bylo vyvráceno několik experimentů, které vyvracely tuto hypotézu a prokázaly, že za vzorování je ve skutečnosti gradient skutečných molekul Dpp.
Existují mutantní formy Dpp receptoru Tkv, které se chovají, jako by přijímaly vysoké množství Dpp signálu i v nepřítomnosti Dpp. Buňky, které obsahují tento mutantní receptor, se chovají, jako by byly v prostředí s vysokým Dpp, jako je oblast v blízkosti pruhu buněk produkujících Dpp. Generováním malých skvrn těchto buněk v různých částech tkáně křídla byli vyšetřovatelé schopni rozlišit, jak Dpp působí na vzor tkáně. Pokud buňky, které přijímají signál Dpp, instruují své sousedy v kaskádě, pak by se měla objevit další centra vzorkování tkáně v místech mutantních buněk, které, jak se zdá, přijímají signalizaci vysokého Dpp, ale samy neprodukují žádný Dpp. Pokud je však nutná fyzická přítomnost Dpp, pak by buňky v blízkosti mutantů neměly být vůbec ovlivněny. Experimenty zjistily, že druhý případ je pravdivý, což naznačuje, že Dpp se chová jako morfogen.[16]
Běžným způsobem, jak posoudit rozdíly ve vzorcích tkání v létajícím křídle, je podívat se na vzorec žil v křídle. U mušek, kde je narušena schopnost Dpp difundovat přes tkáň, je umístění žil posunuto od polohy u normálních mušek a křídlo je obecně menší.[17]
Dpp byl také navržen jako regulátor růstu a velikosti tkáně, klasický problém ve vývoji. Společným problémem pro organismy s mnohobuněčnými orgány, které musí růst z počáteční velikosti, je vědět, kdy přestat růst po dosažení odpovídající velikosti. Protože Dpp je přítomen v gradientu, je možné, že sklon gradientu může být měřítkem, kterým tkáň určuje, jak velká je. Pokud je množství Dpp u zdroje pevné a množství na okraji tkáně je nulové, pak se strmost gradientu sníží, jak se zvětší velikost tkáně a vzdálenost mezi zdrojem a okrajem. Experimenty, kde je v tkáni křídla indukován uměle strmý gradient Dpp, vedly k významně zvýšenému množení buněk, což podpořilo hypotézu strmosti.[18]
Tvorba gradientu Dpp na disku Imaginal Wing
Tvar gradientu Dpp je určen čtyřmi kinetickými parametry ligandu, které jsou ovlivněny biologickými parametry:[19][20]
- Efektivní koeficient difúze, který je závislý na extracelulární difúzi, rychlostech intracelulárního přenosu a kinetika vazby / uvolňování receptoru.
- Účinné rychlosti extracelulární a intracelulární degradace.
- Rychlost produkce v závislosti na produkční cestě Dpp.
- Imobilní zlomek (parametr spojený s metodou používanou k měření kinetiky Dpp, FRAP ).
Je důležité si uvědomit, že jeden biologický parametr může ovlivnit více kinetických parametrů. Například hladiny receptoru ovlivní jak difúzní koeficient, tak rychlost degradace.[21]
Mechanismus, kterým se vytváří gradient Dpp, je však stále kontroverzní a nebylo navrženo ani prokázáno žádné úplné vysvětlení. Čtyři hlavní kategorie teorií za vznikem gradientu jsou volná difúze, omezená difúze, transcytóza, a cytonéma - asistovaná doprava.
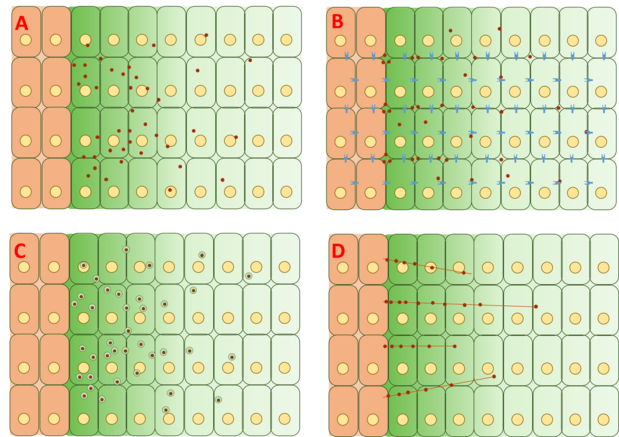
Model volné / omezené difúze
Model volné difúze předpokládá, že Dpp volně difunduje extracelulární matricí a degraduje se prostřednictvím degradačních událostí zprostředkovaných receptorem. Testy FRAP argumentovaly proti tomuto modelu tím, že poznamenávaly, že difúze GFP-Dpp neodpovídá očekávání molekuly podobné velikosti.[20] Jiní však tvrdí, že pozorované rozdíly v difúzi by mohl vysvětlit pomalý krok omezující rychlost dále za procesem, jako je pomalá imobilizace a / nebo pomalá degradace samotného Dpp.[22] Jednotlivé molekuly Dpp byly sledovány pomocí fluorescenční korelační spektroskopie (FCS), což ukazuje, že 65% molekul Dpp difunduje rychle (v souladu s modelem volné difúze) a 35% difunduje pomalu (v souladu s Dpp vázaným na receptory nebo glypicans ).
Model omezené difúze zahrnuje účinky geometrie balení buněk a interakce s extracelulární matricí prostřednictvím vazebných událostí s receptory, jako jsou Tkv a heparinsulfát proteoglykany dally a dally-like.[23][24]
Model transcytózy
Transcytotický model předpokládá, že Dpp bude transportován opakovanými cykly intracelulárního receptorem zprostředkovaného endocytózy, přičemž závažnost gradientu je stanovena endocytotickým tříděním Dpp směrem k recyklaci buňkami vs degradací. Tento model byl zpočátku založen na počátečním pozorování, že Dpp se nemohl akumulovat napříč klony, kam se volal kritický protein dynamin nezbytné pro endocytózu byly mutovány do shibire (shi) fenotyp.[25] Jiné experimenty však ukázaly, že se Dpp dokázal akumulovat znovu shi klony, které zpochybňují model transcytózy.[26] Revize teorie stojící za modelem navrhuje, že endocytóza není pro pohyb Dpp nezbytná, ale je zapojena do signalizace Dpp. Dpp se nepodařilo přesunout mezi buňkami se zmutovanými dally a dally-like, dva heparinsulfát proteoglykany (HSPG) běžně se vyskytující v extracelulární matrici. Výsledkem je, že tyto výsledky naznačují, že Dpp se pohybuje po povrchu buňky prostřednictvím omezené extracelulární difúze dally a dally-like, ale samotný transport Dpp se nespoléhá na transcytózu.[27]
Cytonémem zprostředkovaný dopravní model
The cytonéma zprostředkovaný model naznačuje, že Dpp je přímo transportován do cílových buněk prostřednictvím aktin -na základě filopodia nazývané cytonémy, které sahají od apikálního povrchu buněk reagujících na Dpp ke zdrojovým buňkám produkujícím Dpp.[28] Tyto cytonémy byly pozorovány, ale závislost gradientu Dpp na cytonémech nebyla u imaginárních disků křídla definitivně prokázána. Je však známo, že Dpp je vyžadován a dostatečný k rozšíření a udržení cytonémů. Experimenty analyzující dynamiku mezi Dpp a cytonémy byly provedeny v primordiu vzduchového vaku, kde bylo zjištěno, že signalizace Dpp má funkční spojení s cytonemy. Tyto experimenty však nebyly replikovány v imaginárních discích křídel.
Role v měkkýšů
Dpp se také vyskytuje u měkkýšů, kde hraje klíčovou roli při tvorbě skořápky kontrolou tvaru lastury. U mlžů se vyjadřuje, dokud protokol neskončí s požadovaným tvarem, poté přestane jeho výraz.[29] Je také spojován s tvorbou ulity u plžů,[30] s asymetrickou distribucí, která může být spojena s jejich navíjením: růst pláště se zdá být inhibován kde Dpp je vyjádřen.[31]
Reference
- ^ A b Matsuda S, Harmansa S, Affolter M (únor 2016). "BMP morfogenní přechody u much". Hodnocení cytokinů a růstových faktorů. 27: 119–27. doi:10.1016 / j.cytogfr.2015.11.003. PMID 26684043.
- ^ O'Connor MB, Umulis D, Othmer HG, Blair SS (leden 2006). „Tvarování gradientů morfogenu BMP v křídle embrya a kukly Drosophila“. Rozvoj. 133 (2): 183–93. doi:10.1242 / dev.02214. PMC 6469686. PMID 16368928.
- ^ Wharton KA, Ray RP, Gelbart WM (únor 1993). "Pro specifikaci prvků dorzálního vzoru v embryu Drosophila je nezbytný gradient aktivity dekapentaplegiky." Rozvoj. 117 (2): 807–22. PMID 8330541.
- ^ Wang YC, Ferguson EL (březen 2005). „Prostorová bistabilita interakcí Dpp-receptor během dorzální-ventrální tvorby Drosophila“. Příroda. 434 (7030): 229–34. doi:10.1038 / nature03318. PMID 15759004. S2CID 4415152.
- ^ Ferguson EL, Anderson KV (říjen 1992). „Decapentaplegic působí jako morfogen, který organizuje dorzálně-ventrální obrazec v embryu Drosophila“. Buňka. 71 (3): 451–61. doi:10.1016 / 0092-8674 (92) 90514-D. PMID 1423606. S2CID 40423615.
- ^ Arora K, Levine MS, O'Connor MB (listopad 1994). „Šnekový gen kóduje všudypřítomně exprimovaného člena rodiny TGF-beta potřebnou pro specifikaci osudu dorzálních buněk v embryu Drosophila.“. Geny a vývoj. 8 (21): 2588–601. doi:10,1101 / gad.8.21.2588. PMID 7958918.
- ^ Francois V, Solloway M, O'Neill JW, Emery J, Bier E (listopad 1994). „Dorzálně-ventrální vzorování embrya Drosophila závisí na domnělém negativním růstovém faktoru kódovaném krátkým gastrulačním genem“. Geny a vývoj. 8 (21): 2602–16. doi:10,1101 / gad.8.21.2602. PMID 7958919.
- ^ Ross JJ, Shimmi O, Vilmos P, Petryk A, Kim H, Gaudenz K, Hermanson S, Ekker SC, O'Connor MB, Marsh JL (březen 2001). „Kroucená gastrulace je konzervativní extracelulární antagonista BMP“. Příroda. 410 (6827): 479–83. doi:10.1038/35068578. PMID 11260716. S2CID 24986331.
- ^ Srinivasan S, Rashka KE, Bier E (leden 2002). "Vytvoření gradientu morfogenů Sog v embryu Drosophila". Vývojová buňka. 2 (1): 91–101. doi:10.1016 / S1534-5807 (01) 00097-1. PMID 11782317.
- ^ Marqués G, Musacchio M, Shimell MJ, Wünnenberg-Stapleton K, Cho KW, O'Connor MB (říjen 1997). „Produkce gradientu aktivity DPP v časném embryu Drosophila prostřednictvím protichůdných účinků proteinů SOG a TLD“. Buňka. 91 (3): 417–26. doi:10.1016 / S0092-8674 (00) 80425-0. PMID 9363950. S2CID 16613162.
- ^ Frasch M (březen 1995). "Indukce viscerálního a srdečního mezodermu ektodermálním Dpp v časném embryu Drosophila". Příroda. 374 (6521): 464–7. doi:10.1038 / 374464a0. PMID 7700357. S2CID 4330159.
- ^ Nellen D, Affolter M, Basler K (červenec 1994). "Receptorové serin / threoninkinázy zapojené do kontroly vzoru těla Drosophila pomocí decapentaplegic" (PDF). Buňka. 78 (2): 225–37. doi:10.1016/0092-8674(94)90293-3. PMID 8044837. S2CID 13467898.
- ^ Sekelsky JJ, Newfeld SJ, Raftery LA, Chartoff EH, Gelbart WM (březen 1995). "Genetická charakterizace a klonování matek proti dpp, gen potřebný pro dekapentaplegickou funkci u Drosophila melanogaster". Genetika. 139 (3): 1347–58. PMC 1206461. PMID 7768443.
- ^ Campbell G, Tomlinson A (únor 1999). "Transdukce gradientu morfogenů Dpp v křídle Drosophila: regulace cílů Dpp pomocí brinker". Buňka. 96 (4): 553–62. doi:10.1016 / S0092-8674 (00) 80659-5. PMID 10052457. S2CID 16296766.
- ^ Zecca M, Basler K, Struhl G (srpen 1995). "Postupné organizační činnosti engrailed, ježek a decapentaplegic v křídle Drosophila". Rozvoj. 121 (8): 2265–78. PMID 7671794.
- ^ Affolter M, Basler K (září 2007). "Decapentaplegický morfogenní gradient: od formování vzorů po regulaci růstu". Recenze přírody. Genetika. 8 (9): 663–74. doi:10.1038 / nrg2166. PMID 17703237. S2CID 24005278.
- ^ Crickmore MA, Mann RS (leden 2007). „Hoxova kontrola mobility morfogenů a vývoje orgánů regulací glypikanové exprese“. Rozvoj. 134 (2): 327–34. doi:10,1242 / dev.02737. PMID 17166918.
- ^ Rogulja D, Irvine KD (listopad 2005). "Regulace buněčné proliferace morfogenním gradientem". Buňka. 123 (3): 449–61. doi:10.1016 / j.cell.2005.08.030. PMID 16269336. S2CID 18881009.
- ^ Bollenbach T, Pantazis P, Kicheva A, Bökel C, González-Gaitán M, Jülicher F (březen 2008). "Přesnost gradientu Dpp". Rozvoj. 135 (6): 1137–46. doi:10.1242 / dev.012062. PMID 18296653.
- ^ A b Kicheva A, Pantazis P, Bollenbach T, Kalaidzidis Y, Bittig T, Jülicher F, González-Gaitán M (leden 2007). „Kinetika tvorby morfogenního gradientu“ (PDF). Věda. 315 (5811): 521–5. doi:10.1126 / science.1135774. PMID 17255514. S2CID 2096679.
- ^ Crickmore MA, Mann RS (červenec 2006). "Hox kontrola velikosti orgánu regulací produkce a mobility morfogenů". Věda. 313 (5783): 63–8. doi:10.1126 / science.1128650. PMC 2628481. PMID 16741075.
- ^ Zhou S, Lo WC, Suhalim JL, Digman MA, Gratton E, Nie Q, Lander AD (duben 2012). „Volná extracelulární difúze vytváří gradient morfogenů Dpp křídlového disku Drosophila“. Aktuální biologie. 22 (8): 668–75. doi:10.1016 / j.cub.2012.02.065. PMC 3338872. PMID 22445299.
- ^ Müller P, Rogers KW, Yu SR, Brand M, Schier AF (duben 2013). "Morphogen transport". Rozvoj. 140 (8): 1621–38. doi:10,1242 / dev.083519. PMC 3621481. PMID 23533171.
- ^ Lecuit T, Cohen SM (prosinec 1998). „Úrovně Dpp receptorů přispívají k formování gradientu morfogenů Dpp v imaginárním disku křídla Drosophila“. Rozvoj. 125 (24): 4901–7. PMID 9811574.
- ^ Entchev EV, Schwabedissen A, González-Gaitán M (prosinec 2000). "Gradientní tvorba TGF-beta homologu Dpp". Buňka. 103 (6): 981–91. doi:10.1016 / S0092-8674 (00) 00200-2. PMID 11136982. S2CID 6100358.
- ^ Schwank G, Dalessi S, Yang SF, Yagi R, de Lachapelle AM, Affolter M, Bergmann S, Basler K (červenec 2011). "Tvorba gradientu morfogenů Dpp na velké vzdálenosti". PLOS Biology. 9 (7): e1001111. doi:10.1371 / journal.pbio.1001111. PMC 3144185. PMID 21814489.
- ^ Belenkaya TY, Han C, Yan D, Opoka RJ, Khodoun M, Liu H, Lin X (říjen 2004). „Pohyb morfogenů Drosophila Dpp je nezávislý na endocytóze zprostředkované dynaminy, ale je regulován glypikanovými členy heparansulfátových proteoglykanů“. Buňka. 119 (2): 231–44. doi:10.1016 / j.cell.2004.09.031. PMID 15479640. S2CID 10575655.
- ^ Roy S, Huang H, Liu S, Kornberg TB (únor 2014). "Cytonémem zprostředkovaný kontaktně závislý transport signálního proteinu dekapentaplegického Drosophila". Věda. 343 (6173): 1244624. doi:10.1126 / science.1244624. PMC 4336149. PMID 24385607.
- ^ Kin K, Kakoi S, Wada H (květen 2009). „Nová role dpp při tvarování mlžů odhalená v konzervativním vývojovém programu pro měkkýše“. Vývojová biologie. 329 (1): 152–66. doi:10.1016 / j.ydbio.2009.01.021. PMID 19382296.
- ^ Iijima M, Takeuchi T, Sarashina I, Endo K (květen 2008). "Expresní vzorce engrailed a dpp v plži Lymnaea stagnalis". Vývojové geny a evoluce. 218 (5): 237–51. doi:10.1007 / s00427-008-0217-0. PMID 18443822. S2CID 1045678.
- ^ Kurita Y, Deguchi R, Wada H (prosinec 2009). "Časný vývoj a štěpení vzoru japonské fialové slávky, Septifer virgatus". Zoologická věda. 26 (12): 814–20. doi:10,2108 / zsj.26.814. PMID 19968468. S2CID 25868365.
externí odkazy
- Drosophila decapentaplegic - Interaktivní létání
- dekapentaplegický + protein, + Drosophila v americké národní lékařské knihovně Lékařské předměty (Pletivo)