Prostaglandin-endoperoxid syntáza 2 - Prostaglandin-endoperoxide synthase 2






Prostaglandin-endoperoxid syntáza 2 (prostaglandin G / H syntáza a cyklooxygenáza) (The HUGO oficiální symbol je PTGS2; HGNC ID, HGNC: 9605), také známý jako cyklooxygenáza-2 nebo COX-2, je enzym že u lidí je kódován PTGS2 gen.[5] U lidí je to jeden ze dvou cyklooxygenázy. Podílí se na přeměně kyselina arachidonová na prostaglandin H2, důležitý předchůdce prostacyklin, který je vyjádřen v zánět.
Funkce
PTGS2 (COX-2), převádí kyselina arachidonová (AA) na prostaglandin endoperoxid H2. PGHS jsou cíle pro NSAID a specifické inhibitory PTGS2 (COX-2) zvané koxiby. PGHS-2 je sekvenční homodimer. Každý monomer enzymu má a peroxidáza a PTGS (COX) Aktivní stránky. Enzymy PTGS (COX) katalyzují přeměnu kyselina arachidonová na prostaglandiny ve dvou krocích. Nejprve se vodík oddělí od uhlíku 13 kyseliny arachidonové a poté se pomocí PTGS2 (COX-2) přidají dvě molekuly kyslíku, čímž se získá PGG2. Druhý, PGG2 je snížena na PGH2 v aktivním místě peroxidázy. Syntetizovaný PGH2 se převádí na prostaglandiny (PGD2, PGE2, PGF2α ), prostacyklin (PGI2) nebo tromboxan A2 tkáňově specifickými izomerázami. (obrázek 2)[6]
Při metabolizaci kyseliny arachidonové primárně na PGG2 přeměňuje COX-2 také tuto mastnou kyselinu na malá množství racemické směsi 15-Hydroxyikosatetraenové kyseliny (tj. 15-HETE) složené z ~ 22% 15 (R) -HETE a ~ 78% 15 (S) -HETE stereoizomery stejně jako malé množství 11 (R) -HETE.[7] Dva 15-HETE stereoizomery mají vnitřní biologické aktivity, ale co je důležitější, mohou být dále metabolizovány na hlavní třídu látek, lipoxiny. Dále aspirin ošetřený COX-2 metabolizuje kyselinu arachidonovou téměř výlučně na 15 (R) -HETE, který produkt lze dále metabolizovat na epilipoxiny.[8] Lipoxiny a epi-lipoxiny jsou silnými protizánětlivými látkami a mohou přispívat k celkové činnosti obou COX stejně jako k aspirinu.
COX-2 je přirozeně inhibován kalcitriol (aktivní forma vitaminu D).[9][10]
Mechanismus

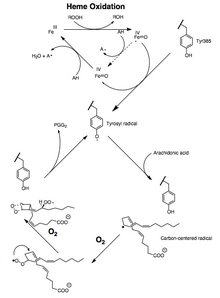
Aktivita peroxidázy i PTGS se během katalýzy inaktivují mechanismy založenými na procesech prvního řádu, což znamená, že aktivity peroxidázy PGHS-2 nebo PTGS klesnou na nulu během 1–2 minut, a to i za přítomnosti dostatečných substrátů.[12][13][14]
Konverzi kyseliny arachidonové na PGG2 lze ukázat jako sérii radikální reakce analogický s polynenasyceným mastné kyseliny autoxidace.[15] 13-pro (S) -vodík se oddělí a dioxygen zachytí pentadienyl radikál na uhlíku 11. 11-Peroxylový radikál cyklizuje na uhlíku 9 a radikál se středem na uhlík generovaný na C-8 cyklizuje na uhlíku 12, čímž vytváří endoperoxid. The allylický generovaný radikál je zachycen dioxygenem na uhlíku 15 za vzniku 15- (S) -peroxylového radikálu; tento radikál se pak redukuje na PGG2 . To je podporováno následujícími důkazy: 1) významný kinetický izotopový efekt je pozorována abstrakce 13-pro (S) -vodíku; 2) radikály se středem na uhlík jsou zachyceny během katalýza;[16] 3) malé množství oxidace produkty se tvoří díky zachycování kyslíku meziproduktu allylového radikálu v pozicích 13 a 15.[17][18]
Další mechanismus, ve kterém je 13-pro (S) -vodík deprotonovaný a karbanion je oxidovaný do a radikální je teoreticky možné. Okysličení kyseliny 10,10-difluorarachidonové na 11- (S) -hydroxyeikosa-5,8,12,14-tetraenovou kyselinu však není v souladu s vytvořením karbaniontového meziproduktu, protože by eliminovalo fluorid za vzniku konjugovaného dienu.[19] Nepřítomnost produktů obsahujících endoperoxid odvozených od 10,10-difluorarachidonové kyseliny naznačuje, že je důležitá karbokace C-10 v PGG2 syntéza.[20] Kationtový mechanismus však vyžaduje, aby tvorba endoperoxidu nastala před odstraněním 13-pro (S) -vodíku. To není v souladu s výsledky izotopových experimentů kyselina arachidonová okysličování.[21]
Struktura

PTGS2 (COX-2) existuje jako homodimer, každý monomer s molekulovou hmotností přibližně 70 kDa. Terciární a kvartérní struktury enzymů PTGS1 (COX-1) a PTGS2 (COX-2) jsou téměř identické. Každá podjednotka má tři různé strukturální domény: krátkou N-terminál epidermální růstový faktor (EGF ) doména; an α-šroubovice skupina vázající membránu; a a C-terminál katalytická doména. PTGS (COX, které lze zaměnit s „cytochrom oxidáza ") enzymy jsou monotopický membránové proteiny; doména vázající membránu se skládá z řady amfipatický α šroubovice s několika hydrofobní aminokyseliny vystaven membránové monovrstvě. PTGS1 (COX-1) a PTGS2 (COX-2) jsou bifunkční enzymy, které provádějí dvě po sobě jdoucí chemické reakce v prostorově odlišných, ale mechanicky spojený aktivní stránky. Oba cyklooxygenáza a peroxidáza aktivní místa jsou umístěna v katalytické doméně, která tvoří přibližně 80% proteinu. Katalytická doména je homologní na savčí peroxidázy, jako je myeloperoxidáza.[23][24]
Bylo zjištěno, že lidský PTGS2 (COX-2) funguje jako konformační heterodimer mající katalytický monomer (E-cat) a alosterický monomer (E-slitina). Heme se váže pouze na peroxidáza místo E-kočky, zatímco substráty, stejně jako jisté inhibitory (např. celecoxib ), navažte COX místo E-cat. E-kočka je regulována E-slitinou způsobem závislým na tom, jaký ligand je vázán na E-slitinu. Podklad a bez substrátu mastné kyseliny (FA) a některé inhibitory PTGS (COX) (např. naproxen ) přednostně se váží na místo PTGS (COX) E-allo. Kyselina arachidonová se může vázat na E-cat a E-allo, ale afinita AA pro E-allo je 25krát vyšší než pro Ecat. Kyselina palmitová, účinný stimulátor huPGHS-2 váže pouze E-slitinu v kokrystalech kyseliny palmitové / myší PGHS-2. Nesubstrátové FA mohou potencovat nebo utlumit Inhibitory PTGS (COX) v závislosti na mastné kyseliny a zda inhibitor váže E-cat nebo E-allo. Studie naznačují, že koncentrace a složení směsi volných mastných kyselin v prostředí, ve kterém PGHS-2 funguje v buňkách, označované také jako tón FA, je klíčovým faktorem regulujícím aktivitu PGHS-2 a jeho reakci na PTGS ( COX) inhibitory.[22]
Klinický význam

PTGS2 (COX-2) je za normálních podmínek ve většině buněk nevyjádřený, ale během něj se nacházejí zvýšené hladiny zánět. PTGS1 (COX-1) je konstitutivně exprimován v mnoha tkáních a je převládající formou v žaludeční sliznici a v ledvinách. Inhibice PTGS1 (COX-1) snižuje bazální produkce cytoprotektivní PGE2 a PGI2 v žaludek, které mohou přispět k žaludeční vředy. Protože PTGS2 (COX-2) je obecně exprimován pouze v buňkách, kde prostaglandiny jsou upregulovaní (např. během zánětu), u kandidátů na léky, kteří selektivně inhibují PTGS2 (COX-2), bylo podezření, že vykazují méně vedlejší efekty[24] ale ukázalo se, že podstatně zvyšuje riziko kardiovaskulárních příhod, jako je infarkt a mrtvice. Protichůdné účinky mohou vysvětlovat dva různé mechanismy. Nízkodávkovaný aspirin chrání před infarkty a mrtvicí blokováním PTGS1 (COX-1) před tvorbou prostaglandinu zvaného tromboxan A2. Lepí krevní destičky k sobě a podporuje srážení; jeho inhibice pomáhá předcházet srdečním onemocněním. Na druhou stranu je PTGS2 (COX-2) důležitějším zdrojem prostaglandinů, zejména prostacyklinu, který se nachází ve výstelce cév. Prostacyklin uvolňuje nebo uvolňuje destičky, takže selektivní inhibitory COX-2 (koxiby) zvyšují riziko kardiovaskulárních příhod v důsledku srážení.[26]
Nesteroidní protizánětlivé léky (NSAID) inhibují prostaglandin produkce společností PTGS1 (COX-1) a PTGS2 (COX-2). NSAID selektivní pro inhibici PTGS2 (COX-2) jsou méně pravděpodobné, než způsobí tradiční léky gastrointestinální nepříznivé účinky, ale může způsobit kardiovaskulární události, jako např srdeční selhání, infarkt myokardu, a mrtvice. Studie s člověkem farmakologie a genetika geneticky manipulováno hlodavci a další zvířecí modely a randomizované studie naznačují, že je to způsobeno potlačením kardioprotektiva závislého na PTGS2 (COX-2) prostaglandiny, prostacyklin zejména.[27]
Exprese PTGS2 (COX-2) je u mnoha druhů rakoviny upregulována. Nadměrná exprese PTGS2 (COX-2) spolu se zvýšenou angiogenezí a expresí SLC2A1 (GLUT-1) je významně spojena s karcinomy žlučníku.[28] Produkt PTGS2 (COX-2), PGH2 je převeden prostaglandin E2 syntáza do PGE2, což může stimulovat progresi rakoviny. V důsledku toho může mít inhibice PTGS2 (COX-2) výhodu v prevenci a léčbě těchto typů rakoviny.[29][30]
Exprese COX-2 byla nalezena v lidských idiopatických epiretinálních membránách.[31] Blokování cyklooxygenáz lornoxicam v akutním stadiu zánětu snížila frekvenci tvorby membrány o 43% v rozptýlit model PVR a o 31% v EU concanavalin jeden. Lornoxikam nejen normalizoval expresi cyklooxygenáz v obou modelech PVR, ale také neutralizoval změny sítnice a cévnatka tloušťka způsobená injekcí prozánětlivých látek. Tato fakta podtrhují význam cyklooxygenáz a prostaglandinů při vývoji PVR.[32]
Upregulace genu PTGS2 byla také spojena s několika stadii lidské reprodukce. Přítomnost genu se nachází v choriový talíř, v amnion epitel, syncytiotrofoblasty, klkové fibroblasty, chorionické trofoblasty, amniotické trofoblasty, stejně jako bazální destička placenty, v desítkové buňky a extravilózní cytotrofoblasty. Během procesu chorioamnionitida / deciduitis, upregulace PTGS2 v EU amnion a choriodecidua je jedním ze tří omezených účinků zánětu v děloha. Zvýšená exprese genu PTGS2 v plodové membrány je spojen s přítomností zánětu, který způsobuje expresi genu prostaglandinu v děloze a imunolokalizaci prostaglandin proteiny dráhy v choriových trofoblastových buňkách a přilehlých decidua nebo choriodecidua. PTGS2 je spojen se zánětlivým systémem a byl pozorován u zánětlivých leukocyty. Bylo poznamenáno, že existuje pozitivní korelace s expresí PTGS2 v amnionu během spontánního porodu a bylo zjištěno, že má zvýšenou expresi s gestačním věkem po přítomnosti porodu, aniž by byla pozorována změna amnionu a choriodecidua během předčasného nebo termínovaného porodu. Dodatečně, oxytocin stimuluje expresi PTGS2 v buňky myometria.[33]
Bylo prokázáno, že nositelé mutantní alely PTGS2 5939C v čínské populaci Han mají vyšší riziko rakovina žaludku. Kromě toho bylo nalezeno spojení mezi Helicobacter pylori infekce a přítomnost alely 5939C.[34]
Interakce
Bylo prokázáno, že PTGS2 komunikovat s Caveolin 1.[35]
Dějiny
PTGS2 (COX-2) byl objeven v roce 1991 Daniel Simmons laboratoř[36] na univerzitě Brighama Younga.
Viz také
- Kyselina arachidonová
- Cyklooxygenáza
- Cyklooxygenáza 1
- NSAID
- Objev a vývoj selektivních inhibitorů COX-2
- Selektivní inhibitor COX-2
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000073756 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000032487 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Hla T, Neilson K (srpen 1992). "Lidská cyklooxygenáza-2 cDNA". Proc. Natl. Acad. Sci. USA. 89 (16): 7384–8. Bibcode:1992PNAS ... 89.7384H. doi:10.1073 / pnas.89.16.7384. PMC 49714. PMID 1380156.
- ^ O'Banion MK (1999). „Cyklooxygenáza-2: molekulární biologie, farmakologie a neurobiologie“. Crit Rev Neurobiol. 13 (1): 45–82. doi:10.1615 / critrevneurobiol.v13.i1.30. PMID 10223523.
- ^ Mulugeta S, Suzuki T, Hernandez NT, Griesser M, Boeglin WE, Schneider C (2010). "Identifikace a absolutní konfigurace dihydroxy-arachidonových kyselin vytvořených okysličením 5S-HETE nativním a aspirin-acetylovaným COX-2". J. Lipid Res. 51 (3): 575–85. doi:10.1194 / ml. M001719. PMC 2817587. PMID 19752399.
- ^ Serhan CN (2005). „Lipoxiny a 15-epi-lipoxiny spouštěné aspirinem jsou prvními lipidovými mediátory endogenního protizánětlivého účinku a rozlišení.“ Prostaglandiny Leukot. Podstatné. Mastné kyseliny. 73 (3–4): 141–62. doi:10.1016 / j.plefa.2005.05.002. PMID 16005201.
- ^ Wang Q1, He Y, Shen Y, Zhang Q, Chen D, Zuo C, Qin J, Wang H, Wang J, Yu Y. (2014). „Vitamin D inhibuje expresi COX-2 a zánětlivou reakci zaměřením na člena nadrodiny thioesterázy 4“. J Biol Chem. 289 (17): 11681–11694. doi:10.1074 / jbc.M113.517581. PMC 4002078. PMID 24619416.CS1 maint: používá parametr autoři (odkaz)
- ^ Kassi E1, Adamopoulos C, Basdra EK, Papavassiliou AG. (2013). „Role vitaminu D při ateroskleróze“. Oběh. 128 (23): 2517–2531. doi:10.1161 / CIRCULATIONAHA.113.002654. PMID 24297817.CS1 maint: používá parametr autoři (odkaz)
- ^ PDB: 3OLT
- ^ Smith WL, Garavito RM, DeWitt DL (prosinec 1996). "Prostaglandin endoperoxid H syntázy (cyklooxygenázy) -1 a -2". J. Biol. Chem. 271 (52): 33157–60. doi:10.1074 / jbc.271.52.33157. PMID 8969167.
- ^ Wu G, Wei C, Kulmacz RJ, Osawa Y, Tsai AL (duben 1999). „Mechanistická studie auto-inaktivace aktivity peroxidázy v prostaglandin H syntáze-1“. J. Biol. Chem. 274 (14): 9231–7. doi:10.1074 / jbc.274.14.9231. PMID 10092596.
- ^ Callan OH, So OY, Swinney DC (únor 1996). „Kinetické faktory, které určují afinitu a selektivitu pro inhibici pomalé vazby lidské prostaglandin H syntázy 1 a 2 indomethacinem a flurbiprofenem“. J. Biol. Chem. 271 (7): 3548–54. doi:10.1074 / jbc.271.7.3548. PMID 8631960.
- ^ Porter NA (1986). "Mechanismy pro autoxidaci polynenasycených lipidů". Účty chemického výzkumu. 19 (9): 262–8. doi:10.1021 / ar00129a001.
- ^ Mason RP, Kalyanaraman B, Tainer BE, Eling TE (červen 1980). "Meziprodukt volných radikálů zaměřený na uhlík v oxidaci prostaglandin syntetázy pomocí kyseliny arachidonové. Zachytávání spinů a studie absorpce kyslíku". J. Biol. Chem. 255 (11): 5019–22. PMID 6246094.
- ^ Hecker M, Ullrich V, Fischer C, Meese CO (listopad 1987). "Identifikace nových metabolitů kyseliny arachidonové vytvořených prostaglandin H syntázou". Eur. J. Biochem. 169 (1): 113–23. doi:10.1111 / j.1432-1033.1987.tb13587.x. PMID 3119336.
- ^ Xiao G, Tsai AL, Palmer G, Boyar WC, Marshall PJ, Kulmacz RJ (únor 1997). „Analýza hydroperoxidem indukovaných tyrosylových radikálů a aktivity lipoxygenázy v lidské prostaglandin H-syntáze-2 ošetřené aspirinem“. Biochemie. 36 (7): 1836–45. doi:10.1021 / bi962476u. PMID 9048568.
- ^ Kwok PY, Muellner FW, Fried J (červen 1987). „Enzymatické konverze kyseliny 10,10-difluorarachidonové s PGH syntázou a sójovou lipoxygenázou“. Journal of the American Chemical Society. 109 (12): 3692–3698. doi:10.1021 / ja00246a028.
- ^ Dean AM, Dean FM (květen 1999). „Karbokace při syntéze prostaglandinů cyklooxygenázou PGH syntázy? Radikální odklon!“. Protein Sci. 8 (5): 1087–98. doi:10.1110 / ps.8.5.1087. PMC 2144324. PMID 10338019.
- ^ Hamberg M, Samuelsson B (listopad 1967). "O mechanismu biosyntézy prostaglandinů E-1 a F-1-alfa". J. Biol. Chem. 242 (22): 5336–43. PMID 6070851.
- ^ A b Dong L, Vecchio AJ, Sharma NP, Jurban BJ, Malkowski MG, Smith WL (květen 2011). "Lidská cyklooxygenáza-2 je sekvenční homodimer, který funguje jako konformační heterodimer". J. Biol. Chem. 286 (21): 19035–46. doi:10.1074 / jbc.M111.231969. PMC 3099718. PMID 21467029.
- ^ Picot D, Loll PJ, Garavito RM (leden 1994). „Rentgenová krystalová struktura membránového proteinu prostaglandin H2 syntáza-1“. Příroda. 367 (6460): 243–9. Bibcode:1994Natur.367..243P. doi:10.1038 / 367243a0. PMID 8121489.
- ^ A b Kurumbail RG, Kiefer JR, Marnett LJ (prosinec 2001). "Cyklooxygenázové enzymy: katalýza a inhibice". Curr. Opin. Struct. Biol. 11 (6): 752–60. doi:10.1016 / S0959-440X (01) 00277-9. PMID 11751058.
- ^ PDB: 3PGH
- ^ Ruan, C. H .; Takže S.P .; Ruan, K.H. (2011). „Indukovatelný COX-2 dominuje nad COX-1 v biosyntéze prostacyklinu: Mechanismy rizika inhibitoru COX-2 pro srdeční choroby“. Humanitní vědy. 88 (1–2): 24–30. doi:10.1016 / j.lfs.2010.10.017. PMC 3046773. PMID 21035466.
- ^ Wang D, Patel VV, Ricciotti E, Zhou R, Levin MD, Gao E, Yu Z, Ferrari VA, Lu MM, Xu J, Zhang H, Hui Y, Cheng Y, Petrenko N, Yu Y, FitzGerald GA (květen 2009 ). „Kardiomyocyt cyklooxygenáza-2 ovlivňuje srdeční rytmus a funkci“. Proc. Natl. Acad. Sci. USA. 106 (18): 7548–52. Bibcode:2009PNAS..106,7548 W.. doi:10.1073 / pnas.0805806106. PMC 2670242. PMID 19376970.
- ^ Legan M (srpen 2010). „Cyklooxygenáza-2, p53 a transportér glukózy-1 jako prediktory malignity ve vývoji karcinomů žlučníku“. Bosn J Basic Med Sci. 10 (3): 192–6. doi:10.17305 / bjbms.2010.2684. PMC 5504494. PMID 20846124.
- ^ EntrezGene 5743
- ^ Menter DG, Schilsky RL, DuBois RN (březen 2010). „Cyklooxygenáza-2 a léčba rakoviny: pochopení rizika by mělo stát za odměnu“. Clin. Cancer Res. 16 (5): 1384–90. doi:10.1158 / 1078-0432.CCR-09-0788. PMC 4307592. PMID 20179228.
- ^ KASE, SATORU; SAITO, WATARU; OHNO, SHIGEAKI; ISHIDA, SUSUMU (2010). "Exprese cyklo-oxygenázy-2 v lidské idiopatické epiretinální membráně". Sítnice. 30 (5): 719–723. doi:10.1097 / iae.0b013e3181c59698. PMID 19996819.
- ^ Tichonovič, Marina V .; Erdiakov, Aleksei K .; Gavrilova, Svetlana A. (2017-06-21). „Nesteroidní protizánětlivá terapie potlačuje rozvoj proliferativní vitreoretinopatie účinněji než steroidní“. Mezinárodní oftalmologie. 38 (4): 1365–1378. doi:10.1007 / s10792-017-0594-3. ISSN 0165-5701. PMID 28639085.
- ^ Phillips, Robert J a kol. "Exprese genu prostaglandinové dráhy v lidské placentě, amnionu a choriodeciduě je odlišně ovlivněna předčasným porodem a zánětem dělohy." BMC těhotenství a porod sv. 14 241. 22. července 2014, doi: 10,1186 / 1471-2393-14-241
- ^ Li Y, He W, Liu T, Zhang Q (prosinec 2010). „Nová varianta genu pro cyklooxygenázu-2 v čínské populaci Han je spojena se zvýšeným rizikem karcinomu žaludku“. Mol Diagn Ther. 14 (6): 351–5. doi:10.1007 / bf03256392. PMID 21275453.
- ^ Liou JY, Deng WG, Gilroy DW, Shyue SK, Wu KK (září 2001). „Kolokalizace a interakce cyklooxygenázy-2 s kaveolinem-1 v lidských fibroblastech“. J. Biol. Chem. 276 (37): 34975–82. doi:10,1074 / jbc.M105946200. PMID 11432874.
- ^ Xie WL, Chipman JG, Robertson DL, Erikson RL, Simmons DL (duben 1991). "Exprese genu reagujícího na mitogen kódující prostaglandin syntázu je regulována sestřihem mRNA". Proc. Natl. Acad. Sci. USA. 88 (7): 2692–6. Bibcode:1991PNAS ... 88,2692X. doi:10.1073 / pnas.88.7.2692. PMC 51304. PMID 1849272.[je zapotřebí lepší zdroj ]
Další čtení
- Richards JA, Petrel TA, Brueggemeier RW (únor 2002). "Signální dráhy regulující aromatázu a cyklooxygenázy v normálních a maligních buňkách prsu". J. Steroid Biochem. Mol. Biol. 80 (2): 203–12. doi:10.1016 / S0960-0760 (01) 00187-X. PMID 11897504.
- Wu T, Wu H, Wang J, Wang J (2011). "Exprese a buněčná lokalizace cyklooxygenáz a syntáz prostaglandinu E v hemoragickém mozku". J Neuroinflamation. 8: 22. doi:10.1186/1742-2094-8-22. PMC 3062590. PMID 21385433.
- Koki AT, Khan NK, Woerner BM, Seibert K, Harmon JL, Dannenberg AJ, Soslow RA, Masferrer JL (leden 2002). „Charakterizace cyklooxygenázy-2 (COX-2) během tumorigeneze u lidských epiteliálních karcinomů: důkazy o možném klinickém využití inhibitorů COX-2 u epiteliálních karcinomů“. Prostaglandiny Leukot. Podstatné. Mastné kyseliny. 66 (1): 13–8. doi:10.1054 / plef.2001.0335. PMID 12051953.
- Saukkonen K, Rintahaka J, Sivula A, Buskens CJ, Van Rees BP, Rio MC, Haglund C, Van Lanschot JJ, Offerhaus GJ, Ristimaki A (říjen 2003). „Cyclooxygenase-2 and gastric carcinogenesis“. APMIS. 111 (10): 915–25. doi:10.1034 / j.1600-0463.2003.1111001.x. PMID 14616542.
- Sinicrope FA, Gill S (2004). "Role cyklooxygenázy-2 u kolorektálního karcinomu". Metastáza rakoviny Rev. 23 (1–2): 63–75. doi:10.1023 / A: 1025863029529. PMID 15000150.
- Jain S, Khuri FR, Shin DM (2004). „Prevence rakoviny hlavy a krku: současný stav a vyhlídky do budoucna“. Curr Probl Cancer. 28 (5): 265–86. doi:10.1016 / j.currproblcancer.2004.05.003. PMID 15375804.
- Saba N, Jain S, Khuri F (2004). "Chemoprevence u rakoviny plic". Curr Probl Cancer. 28 (5): 287–306. doi:10.1016 / j.currproblcancer.2004.05.005. PMID 15375805.
- Cardillo I, Spugnini EP, Verdina A, Galati R, Citro G, Baldi A (říjen 2005). "Cox a mezoteliom: přehled". Histol. Histopathol. 20 (4): 1267–74. PMID 16136507.
- Brueggemeier RW, Díaz-Cruz ES (březen 2006). „Vztah mezi aromatázou a cyklooxygenázami u rakoviny prsu: potenciál nových terapeutických přístupů“. Minerva Endocrinol. 31 (1): 13–26. PMID 16498361.
- Fujimura T, Ohta T, Oyama K, Miyashita T, Miwa K (březen 2006). „Role cyklooxygenázy-2 v karcinogenezi rakoviny gastrointestinálního traktu: přehled a zpráva o osobních zkušenostech“. Svět J. Gastroenterol. 12 (9): 1336–45. doi:10,3748 / wjg.v12.i9.1336. PMC 4124307. PMID 16552798.
- Bingham S, Beswick PJ, Blum DE, Gray NM, Chessell IP (říjen 2006). "Role cylooxygenase dráhy v nocicepci a bolesti". Semin. Cell Dev. Biol. 17 (5): 544–54. doi:10.1016 / j.semcdb.2006.09.001. PMID 17071117.
- Minghetti L, Pocchiari M (2007). Cyklooxygenáza-2, prostaglandin E2 a mikrogliální aktivace u prionových onemocnění. Int. Rev. Neurobiol. Mezinárodní přehled neurobiologie. 82. str. 265–75. doi:10.1016 / S0074-7742 (07) 82014-9. ISBN 9780123739896. PMID 17678966.
externí odkazy
- Nextbio
- NSAID a kardiovaskulární riziko vysvětleno podle studií z Perelman School of Medicine
- Wolfe MM (prosinec 2004). „Rofecoxib, Merck a FDA“. N. Engl. J. Med. 351 (27): 2875–8, odpověď autora 2875–8. doi:10.1056 / NEJM200412303512719. PMID 15625749.