Fosfatáza lehkého řetězce myosinu - Myosin-light-chain phosphatase
| Myosinová fosfatáza s lehkým řetězcem | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Struktura komplexu mezi PP1 a částí MYPT1, generovaná od 1s70[1] | |||||||||
| Identifikátory | |||||||||
| EC číslo | 3.1.3.53 | ||||||||
| Číslo CAS | 86417-96-1 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Myosin fosfatáza s lehkým řetězcem, běžněji nazývané myosin fosfatáza (ES 3.1.3.53 ), je enzym (konkrétně a serin / threonin-specifický protein fosfatáza ) že defosforyláty regulační lehký řetězec myosin II. K této defosforylační reakci dochází v tkáň hladkého svalstva a zahajuje relaxační proces svalových buněk. Myosin fosfatáza tedy odbourává svalová kontrakce proces zahájen myosin kináza lehkého řetězce. Enzym se skládá ze tří podjednotek: katalytické oblasti (proteinová fosfatáza 1 nebo PP1) myosinová vazebná podjednotka (MYPT1) a třetí podjednotka (M20) neznámé funkce. Katalytická oblast používá dva mangan ionty jako katalyzátory k defosforylaci lehkých řetězců na myosinu, což způsobuje konformační změnu myosinu a uvolňuje sval. Enzym je vysoce konzervovaný[1] a nachází se ve tkáni hladkého svalstva všech organismů. I když je známo, že myosin fosfatáza je regulována rho-asociované protein kinázy, existuje aktuální debata o tom, zda jiné molekuly, jako např kyselina arachidonová a tábor, také regulují enzym.[2]
Funkce
Hladká svalová tkáň je většinou vyrobena z aktin a myosin,[3] dva proteiny, které spolu interagují a vytvářejí svalovou kontrakci a relaxaci. Myosin II, také známý jako konvenční myosin, má dva těžké řetězce, které se skládají z domén hlavy a ocasu, a čtyři lehké řetězce (dva na hlavu), které se vážou na těžké řetězce v oblasti „krku“. Když se sval potřebuje stahovat, vápník ionty proudí do cytosol z sarkoplazmatické retikulum, kde aktivují kalmodulin, který se zase aktivuje myosin kináza lehkého řetězce (MLC kináza). MLC kináza fosforyluje lehký řetězec myosinu (MLC20) na zbytku Ser-19. Tato fosforylace způsobuje aktivaci konformační změny v myosinu crossbridge cyklistika a způsobení kontrakce svalu. Protože myosin prochází konformační změnou, sval zůstane kontrahovaný, i když jsou koncentrace vápníku a aktivované MLC kinázy uvedeny na normální hladinu. Aby došlo k uvolnění svalu, musí být změněna konformační změna.[4]
Když se myosin fosfatáza váže na myosin, odstraňuje fosfátová skupina. Bez skupiny se myosin vrátí do své původní konformace, ve které nemůže interagovat s aktinem a udržet sval napnutý, takže se sval uvolní. Sval zůstane v této uvolněné poloze, dokud nebude myosin fosforylován MLC kinázou a neprovede konformační změnu.
Struktura
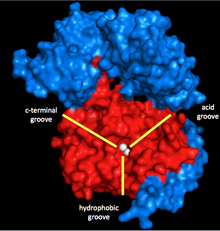
Myosin fosfatáza je vyrobena ze tří podjednotek. Katalytická podjednotka, PP1, je jednou z nejdůležitějších Ser / Thr fosfatáz v eukaryotické buňky, protože hraje roli v glykogen metabolismus, intracelulární transport, syntéza proteinů a buněčné dělení stejně jako kontrakce hladkého svalstva.[5] Protože je to tak důležité pro základní buněčné funkce a protože v buňkách je mnohem méně proteinových fosfatáz než kináz,[6] Struktura a funkce PP1 je vysoce konzervovaná (ačkoli specifická izoforma používaná v myosin fosfatáze je 8 izoforma, PP1δ).[7] PP1 pracuje s použitím dvou manganových iontů jako katalyzátorů pro defosforylaci (viz níže).
Kolem těchto iontů je rozštěp ve tvaru Y se třemi drážkami: hydrofobní, kyselý a C-koncový. Pokud PP1 není vázán na žádnou jinou podjednotku, není to nijak zvlášť specifické. Když se však váže na druhou podjednotku myosin fosfatázy, MYPT1 (MW ~ 130 kDa), tato katalytická štěrbina změní konfiguraci. To má za následek dramatický nárůst specificity myosinu.[1] Je tedy jasné, že MYPT1 má velkou regulační moc nad PP1 a myosin fosfatázou, a to i bez přítomnosti dalších aktivátorů nebo inhibitorů.
Třetí podjednotka, M20 (nezaměňovat s MLC20, kritická regulační podjednotka myosinu), je nejmenší a nejtajemnější podjednotkou. V současné době je o M20 známo jen málo, kromě toho, že to není nutné pro katalýzu, protože odstranění podjednotky nemá vliv na obrat nebo selektivitu.[1] Zatímco někteří věří, že by to mohlo mít regulační funkci, zatím nebylo nic stanoveno.[2]
Mechanismus
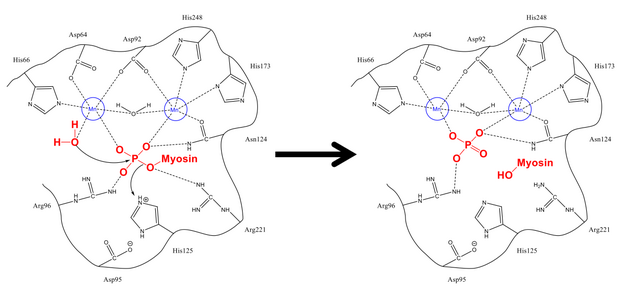
Mechanismus odstraňování fosfátu ze Ser-19 je velmi podobný jiným defosforylačním reakcím v buňce, jako je aktivace glykogen syntáza. Myosinová regulační podjednotka MLC20 se váže na hydrofobní i kyselé drážky PP1 a MYPT1, regulačního místa na myosin fosfatáze.[1][8] Jakmile jsou ve správné konfiguraci, oba fyosforylovali serin a volná molekula vody jsou stabilizovány zbytky vodíkové vazby v aktivním místě, stejně jako kladně nabité ionty (které silně interagují s negativní fosfátovou skupinou). His-125 (na myosin fosfatáze) daruje proton k Ser-19 MLC20) a molekula vody útočí na atom fosforu. Po zamíchání protonů ke stabilizaci (k čemuž dochází rychle ve srovnání s útokem na fosfor) se vytvoří fosfát a alkohol a oba opouštějí aktivní místo.
Regulace a lidské zdraví
Regulační dráhy MLC kinázy jsou dobře zavedené, ale až do konce 80. let se předpokládalo, že myosin fosfatáza není regulovaná a kontrakce / relaxace zcela závisí na aktivitě MLC kinázy.[2] Od 80. let 20. století však byl objeven a důkladně zkoumán inhibiční účinek proteinové kinázy spojené s rho.[11] RhoA GTP se aktivuje Rho-kináza, který fosforyluje MYPT1 na dvou hlavních inhibičních místech, Thr-696 a Thr-866.[12][13] To plně prokazuje hodnotu MYPT1, a to nejen pro zvýšení reakční rychlosti a specificity, ale také pro výrazné zpomalení reakce. Kdy však telokin je přidán, účinně ruší účinek Rho-kinázy, i když nedefosforyluje MYPT1.[12]
Jedna další navrhovaná regulační strategie zahrnuje kyselinu arachidonovou. Když se přidá kyselina arachidonová do napjaté svalové tkáně, kyselina snižuje rychlost defosforylace (a tedy relaxace) myosinu. Není však jasné, jak kyselina arachidonová funguje jako inhibitor.[4] Dvě konkurenční teorie jsou, že buď kyselina arachidonová působí jako spolu-posel ve výše uvedené kaskádě rho-kinázy, nebo že se váže na c-terminál MYPT1.[4]
Když regulační systémy myosin fosfatázy začnou selhávat, mohou to mít závažné zdravotní důsledky. Vzhledem k tomu, že hladký sval se nachází v dýchacích, oběhových a reprodukčních systémech člověka (a také na jiných místech), pokud se hladký sval již nemůže uvolnit kvůli chybné regulaci, pak existuje celá řada problémů od astma, hypertenze, a erektilní dysfunkce může mít za následek.[4][14]
Viz také
Reference
- ^ A b C d E Terrak, Mohammed; Kerff, Frederic; et al. (17. června 2004). „Strukturální základ regulace proteinové fosfatázy 1“. Příroda. 429 (6993): 780–4. Bibcode:2004 Natur.429..780T. doi:10.1038 / nature02582. PMID 15164081.
- ^ A b C Hartshorne, DJ; Ito, M (květen 1998). „Fosfatáza lehkého řetězce myosinu: složení podjednotek, interakce a regulace“. J Muscle Res Cell Motil. 19 (4): 325–41. doi:10.1023 / A: 1005385302064. PMID 9635276. S2CID 27448238.
- ^ Stránka 174 v: Buňka hladkého svalstva cév: molekulární a biologické reakce na extracelulární matrici. Autoři: Stephen M. Schwartz, Robert P. Mecham. Střih: Stephen M. Schwartz, Robert P. Mecham. Přispěvatelé: Stephen M. Schwartz, Robert P. Mecham. Vydavatel: Academic Press, 1995. ISBN 0-12-632310-0, ISBN 978-0-12-632310-8
- ^ A b C d Webb, R. Clinton (listopad 2003). "Hladká svalová kontrakce a relaxace". Pokroky ve výuce fyziologie. 27 (4): 201–6. doi:10.1152 / advan.00025.2003. PMID 14627615.
- ^ Hurley, Thomas; Yang, Jie; et al. (18. července 2007). "Strukturální základ pro regulaci proteinové fosfatázy 1 inhibitorem-2". J. Biol. Chem. 282 (39): 28874–83. doi:10.1074 / jbc.m703472200. PMID 17636256.
- ^ Cohen, Patricia T. W. (15. ledna 2002). "Protein Phosphatase 1-Targeted in Many Directions". J Cell Sci. 115 (2): 780–4. PMID 11839776.
- ^ Fujioka, M; Takahashi, N (1. dubna 1998). "Nová izoforma lidské myosin fosfatázy cílené / regulační podjednotky (MYPT2): cDNA klonování, tkáňová exprese a chromozomální mapování". Genomika. 49 (1): 325–41. doi:10.1006 / geno.1998.5222. PMID 9570949.
- ^ Gomperts, Bastein D. (19. srpna 2009). Transdukce signálu: 2. vydání. London: Academic Press. ISBN 978-0123694416.
- ^ Shi, Yigong (30. října 2009). "Serin / threoninové fosfatázy: mechanismus strukturou". Buňka. 139 (3): 468–84. doi:10.1016 / j.cell.2009.10.006. PMID 19879837. S2CID 13903804. Citováno 9. března 2015.
- ^ Lee, Ernest Y.C .; Zhang, Lifang; et al. (15. března 1999). „Fosforyláza fosfatáza: Nové horizonty pro starý enzym“. Frontiers in Bioscience. 4 (1–3): 270–85. doi:10,2741 / závětří. PMID 10077543. Citováno 9. března 2015.
- ^ Wang, Yuepeng; Riddick, Nadeen; et al. (27. února 2009). "Regulace ROCK izoformy myosin fosfatázy a kontraktility v buňkách hladkého svalstva cév". Circ. Res. 104 (4): 531–40. doi:10.1161 / circresaha.108.188524. PMC 2649695. PMID 19131646.
- ^ A b Khromov, ES; Momotani, K .; et al. (27. dubna 2012). "Molekulární mechanismus tukinkinem zprostředkované disinhibice myosinové fosfatázy lehkého řetězce a cAMP / cGMP-indukované relaxace hladkého svalu gastrointestinálního traktu". J Biol Chem. 287 (25): 20975–85. doi:10.1074 / jbc.m112.341479. PMC 3375521. PMID 22544752.
- ^ Somlyo, Andrew P .; Somlyo, Avril V. (10. listopadu 1999). „Transdukce signálu G-proteiny, rho-kinázou a proteinovou fosfatázou k hladkému svalu a mimosvalovému myosinu II“. Fyziologický časopis. 522 (2): 177–85. doi:10.1111 / j.1469-7793.2000.t01-2-00177.x. PMC 2269761. PMID 10639096.
- ^ Aguilar, Hector; Mitchell, B.F. (7. května 2010). „Fyziologické cesty a molekulární mechanismy regulující kontraktilitu dělohy“. Aktualizace lidské reprodukce. 16 (6): 725–44. doi:10.1093 / humupd / dmq016. PMID 20551073. Citováno 9. března 2015.
Další čtení
- Pato MD, Adelstein RS (1983). „Čištění a charakterizace více podjednotkové fosfatázy z hladkého svalstva krůtího žaludku. Účinek navázání kalmodulinu na kinázu lehkého řetězce myosinu na defosforylaci“. J. Biol. Chem. 258 (11): 7047–54. PMID 6304072.
- Kimura K; et al. (1996). "Regulace myosin fosfatázy pomocí Rho a Rho-asociované kinázy (Rho-kináza)". Věda. 273 (5272): 245–248. Bibcode:1996Sci ... 273..245K. doi:10.1126 / science.273.5272.245. PMID 8662509. S2CID 37249779.