DNáza aktivovaná kaspázou - Caspase-activated DNase





| Fragmentační faktor DNA 40 kDa | |||||||||
|---|---|---|---|---|---|---|---|---|---|
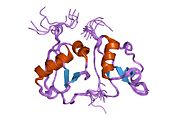
 Krystalová struktura kaspázou aktivované DNAsy (CAD) | |||||||||
| Identifikátory | |||||||||
| Symbol | DFF40 | ||||||||
| Pfam | PF09230 | ||||||||
| InterPro | IPR015311 | ||||||||
| SCOP2 | 1v0d / Rozsah / SUPFAM | ||||||||
| |||||||||
DNáza aktivovaná kaspázou (CAD) nebo Fragmentační faktor DNA podjednotky beta je protein že u lidí je kódován DFFB gen.[5][6][7] Rozkládá DNA během apoptózy a podporuje buněčnou diferenciaci. Obvykle se jedná o neaktivní monomer inhibovaný ICAD. To je štěpeno před dimerizací.
Funkce
Apoptóza je proces autodestrukce buněk, který odstraňuje toxické a / nebo zbytečné buňky během vývoje savců a dalších životních procesů. Apoptotický proces je doprovázen smršťováním a fragmentací buněk a jader a degradací chromozomální DNA na nukleozomální jednotky. Fragmentační faktor DNA (DFF) je heterodimerní protein o obsahu 40 kD (DFFB) a 45 kD (DFFA ) podjednotky. DFFA je substrátem pro kaspázu-3 a během apoptózy spouští fragmentaci DNA. DFF se aktivuje, když je DFFA štěpena kaspázou-3. Štěpené fragmenty DFFA disociují od DFFB, aktivní složky DFF. Bylo zjištěno, že DFFB během apoptózy spouští fragmentaci DNA i kondenzaci chromatinu. Pro tento gen bylo nalezeno několik alternativně sestřižených variant transkriptu kódujících odlišné izoformy, ale biologická platnost některých variant nebyla stanovena.[7]

Přestože je tento gen přítomen v každé buňce, je tento protein exprimován pouze v různých tkáních a buněčných rozmanitostech, jako je pankreas, srdce, tlusté střevo, leukocyty, prostata, vaječníky, placenta, ledviny, slezina a brzlík.[8]
Je také známý jako kaspázou aktivovaná nukleáza (CPAN), faktor fragmentace DNA 40 (DFF-40), DFF2 a DFFB. Kromě toho existují další nomenklatury, které jsou výsledkem kombinace předchozích.[8][9][10][11]
Struktura
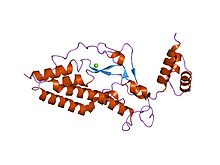
Tento heterodimer je endonukleáza[9][12][13] s vysokým obsahem cystein zbytky.[11] Zůstává neaktivní v rostoucích buňkách, zatímco je spojen s jeho inhibitorem (ICAD, fragmentační faktor DNA 45 kDa, DFFA nebo DFF45), jejichž výsledkem je komplexní ICAD-CAD.[8][9][11][12][14][15][16][17] Jejich disociace umožňuje DFF40 oligomerizovat za vzniku velkého funkčního komplexu, který je sám o sobě aktivní DNázou.[11][12][15][16][17]
DFF40 podjednotka nebo CAD
Váží 40 kDa. Kromě toho obsahuje tři domény tvořící monomer CAD: C1 nebo N-terminální CAD; C2, které odpovídají třem samostatným α řetězcům, a nakonec C3, který je největší a funkčně nejdůležitější. Navíc kombinace aminokyselin C3 vede k 5 a helixům, 4 β lamině a smyčce na katalytickém C-konci, které spolu interagují. Proto se vytváří dutina (aktivní místo), do které se DNA vejde, i když je zde další vazebná oblast odpovědná za stabilní DNA komplex během jeho fragmentace.[8][14][18]
DFF45 podjednotka nebo ICAD
DFFA je kódován alternativně šifrovanými mRNA pocházejícími ze dvou odlišných forem: krátké (ICAD-S) a dlouhé (ICAD-L), které fungují jako specifický chaperon zajišťující správné skládání CAD[10][11][17] Kromě toho obsahuje dva zbytky kyseliny asparagové (Asp117 a Asp224), kde je identifikován CAD, a proto zůstává vázán, dokud Caspase-3 rozděluje tuto unii.[10][14]
Proces aktivace
Obvykle se v neaapoptoticky rostoucích buňkách udržuje kaspáza aktivovaná dnasa pod kontrolou inaktivovaná v cytoplazmě díky spojení s jejím inhibitorem, inhibitorem DNázy aktivované kaspázou (ICAD), také známý jako fragmentační faktor DNA 45 kDa (DFF45).
ICAD je kódován alternativně sestřiženými mRNA, které generují dlouhé (ICAD-L) a krátké (ICAD-S) formy ICAD. ICAD má tedy dvojí funkci; působí jako inhibitor CAD a také jako garde pro syntézu CAD napomáhající správnému sestavení proteinu.[19]
ICAD má dvě místa pro rozpoznávání kaspáz na Asp117 a Asp224. Uvolnění CAD z inhibice ICAD se dosáhne štěpením ICAD na těchto zbytcích Asp pomocí kaspáza-3.[20]
Kaspáza-3 je aktivována v apoptotické buňce.[9] Aktivace kaspázy-3 je požadavkem buňky během raných stadií diferenciace kosterního myoblastu. Své katalytické místo zahrnuje sulfohydrylovou skupinu Cys-285 a imidazolový kruh jeho His-237. Kaspáza-3 His-237 stabilizuje cílový aspartát, což způsobí narušení asociace ICAD a CAD a ponechá endonukleázu CAD aktivní, což jí umožní degradovat chromozomální DNA.
Jakmile je inhibitor uvolněn a aby správně fungoval, musí se dva CAD monomery spojit a vytvořit funkční dimer, který má vertikální symetrii.
Interakce
DFFB bylo prokázáno komunikovat s DFFA.[21][22]
Buněčná diferenciace
Caspase 3 je zodpovědný za buněčná diferenciace, i když není jasné, jak může tento druh proteinu podporovat buňku apoptóza. Signály kaspázy vyplývající z aktivace nukleázy CAD naznačují, že diferenciace buněk je způsobena modifikací CAD ve struktuře chromatinu.
CAD vede k iniciaci rozbití řetězce DNA, ke kterému dochází během terminální diferenciace některých buněk, jako jsou buňky kosterního svalstva. Cílení promotoru p21 je zodpovědné za indukci buněčné diferenciace, která je podporována modifikací nukleárního mikroprostředí DNA.[23]
Buněčná rozmanitost pochází z buněčné diferenciace, která se připisuje aktivaci specifických transkripčních faktorů. Závisí to také na aktivitě proteinu nebo společném signálu. Faktor, který, jak se zdá, indukuje větší diferenciaci buněk, je proteáza kaspázy-3.[24] To bylo identifikováno jako předposlední stádium buněk apoptózových drah.
Některé studie ukázaly, že tato diferenciace je způsobena mnoha substráty CAD kinázy. S odkazem na příklad kosterních buněk je jejich diferenciace spojena se štěpením kinázy MST1.[25]
Kromě toho bylo vidět, že CAD se podílí na tvorbě genomu, jehož DNA se rozpadá v raných fázích buněčné diferenciace. Kromě toho kaspáza 3 indukuje zlomy DNA v promotoru faktoru p21 a tento rozpad řetězce souvisí s expresí genu p21.
Buněčná apoptotická smrt
Proteinová kaspáza DNáza je endonukleáza zapojená do buněčného apoptotického procesu, který usnadňuje rozpad DNA.[26] Buněčná apoptotická smrt je proces prováděný uživatelem cystein proteázy[27] který umožňuje zvířatům držet své homeostáza, také regulováno jinými mechanismy, jako je růst a diferenciace buněk. Tato biologická odpověď je charakterizována chromozomální DNA Degradace v drobných fragmentech v jádru buňky.[28] Po mnoha výzkumech a výzkumech bylo možné zajistit, že za tuto destrukci je díky dlouhému seznamu stimulů hlavní odpovědná DNáza aktivovaná kaspázou.
Jeden z experimentů prováděných vyšetřovateli za účelem prokázání, že teorie byla založena na zavedení mutované formy tohoto proteinu do lidských TF-1 lidských buněk a Jurkatovy buňky, kteří již reagovali na obvyklou (nemutovanou) formu endonukleázy a byli mrtví z apoptózy. Ve výsledku tyto buňky zemřely s přihlédnutím k této genetické modifikaci, ale nevykazovaly rozpad DNA. To byl klíčový důkaz, který dokázal, že forma CAD je součástí této části procesu, protože bez jejího přispění fragmentace neproběhla.[29]
Později bylo zjištěno, že způsob, jakým tento protein indukuje rozpad DNA, je vysvětlen jeho formami CAD a ICAD, které usnadňují vstup i výstup v jádru buňky.[28]
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000169598 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000029027 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Liu X, Zou H, Slaughter C, Wang X (duben 1997). „DFF, heterodimerní protein, který funguje downstream od kaspázy-3 a spouští fragmentaci DNA během apoptózy“. Buňka. 89 (2): 175–84. doi:10.1016 / S0092-8674 (00) 80197-X. PMID 9108473. S2CID 14800864.
- ^ Halenbeck R, MacDonald H, Roulston A, Chen TT, Conroy L, Williams LT (duben 1998). „CPAN, lidská nukleáza regulovaná inhibitorem citlivým na kaspázu DFF45“. Aktuální biologie. 8 (9): 537–40. doi:10.1016 / S0960-9822 (98) 79298-X. PMID 9560346. S2CID 9837862.
- ^ A b "Entrez Gene: DFFB DNA fragmentation factor, 40kDa, beta polypeptide (caspase-activated DNase)".
- ^ A b C d Davidson College. „Deoxyribonukleáza aktivovaná kaspázou (CAD)“. Citováno 21. ledna 2016.
- ^ A b C d Yuste VJ, Sánchez-López I, Solé C, Moubarak RS, Bayascas JR, Dolcet X a kol. (Říjen 2005). „Příspěvek faktoru indukujícího apoptózu, DNázy aktivované kaspázou a inhibitoru DNázy aktivované kaspázou k nukleárnímu fenotypu a degradaci DNA během apoptózy“. The Journal of Biological Chemistry. 280 (42): 35670–83. doi:10,1074 / jbc.M504015200. PMID 16049016.
- ^ A b C Sakahira H, Iwamatsu A, Nagata S (březen 2000). „Specifická chaperonová aktivita inhibitoru DNázy aktivované kaspázou pro DNázu aktivovanou kaspázou“. The Journal of Biological Chemistry. 275 (11): 8091–6. doi:10.1074 / jbc.275.11.8091. PMID 10713130.
- ^ A b C d E Sakahira H, Enari M, Nagata S (květen 1999). "Funkční rozdíly dvou forem inhibitoru DNázy aktivované kaspázou, ICAD-L a ICAD-S". The Journal of Biological Chemistry. 274 (22): 15740–4. doi:10.1074 / jbc.274.22.15740. PMID 10336474.
- ^ A b C Jog NR, Frisoni L, Shi Q, Monestier M, Hernandez S, Craft J, Prak ET, Caricchio R (duben 2012). „DNáza aktivovaná kaspázou je vyžadována pro udržení tolerance k jaderným autoantigenům lupusu“. Artritida a revmatismus. 64 (4): 1247–56. doi:10,1002 / článek 33448. PMC 3292632. PMID 22127758.
- ^ Widlak P, Lanuszewska J, Cary RB, Garrard WT (červenec 2003). "Podjednotkové struktury a stechiometrie proteinů lidského fragmentačního faktoru DNA před a po indukci apoptózy". The Journal of Biological Chemistry. 278 (29): 26915–22. doi:10,1074 / jbc.M303807200. PMID 12748178.
- ^ A b C Reh S, Korn C, Gimadutdinow O, Meiss G (prosinec 2005). „Strukturální základ pro stabilní tvorbu komplexu DNA pomocí DNázy aktivované kaspázou“. The Journal of Biological Chemistry. 280 (50): 41707–15. doi:10,1074 / jbc.m509133200. PMID 16236713.
- ^ A b Widlak P, Li P, Wang X, Garrard WT (březen 2000). "Štěpné preference apoptotické endonukleázy DFF40 (DNáza nebo nukleáza aktivovaná kaspázou) na nahé DNA a chromatinových substrátech". The Journal of Biological Chemistry. 275 (11): 8226–32. doi:10.1074 / jbc.275.11.8226. PMID 10713148.
- ^ A b Sharif-Askari E, Alam A, Rhéaume E, Beresford PJ, Scotto C, Sharma K, Lee D, DeWolf WE, Nuttall ME, Lieberman J, Sékaly RP (červen 2001). „Přímé štěpení lidského fragmentačního faktoru DNA-45 granzymem B indukuje uvolňování DNázy aktivované kaspázou a fragmentaci DNA“. Časopis EMBO. 20 (12): 3101–13. doi:10.1093 / emboj / 20.12.3101. PMC 150191. PMID 11406587.
- ^ A b C Liu X, Zou H, Widlak P, Garrard W, Wang X (květen 1999). „Aktivace apoptotické endonukleázy DFF40 (DNáza nebo nukleáza aktivovaná kaspázou). Oligomerizace a přímá interakce s histonem H1“. The Journal of Biological Chemistry. 274 (20): 13836–40. doi:10.1074 / jbc.274.20.13836. PMID 10318789.
- ^ Uegaki K, Otomo T, Sakahira H, Shimizu M, Yumoto N, Kyogoku Y, Nagata S, Yamazaki T (duben 2000). "Struktura domény CAD kasasy aktivované DNázy a interakce s doménou CAD jejího inhibitoru". Journal of Molecular Biology. 297 (5): 1121–8. doi:10.1006 / jmbi.2000.3643. PMID 10764577.
- ^ PDB: 1V0D; Woo EJ, Kim YG, Kim MS, Han WD, Shin S, Robinson H a kol. (Květen 2004). "Strukturální mechanismus pro inaktivaci a aktivaci CAD / DFF40 v apoptotické dráze". Molekulární buňka. 14 (4): 531–9. doi:10.1016 / S1097-2765 (04) 00258-8. PMID 15149602.
- ^ „CASP3 kaspáza 3 [Homo sapiens (člověk)] - gen - NCBI“.
- ^ Ewing RM, Chu P, Elisma F, Li H, Taylor P, Climie S a kol. (2007). „Mapování interakcí lidských proteinů a proteinů ve velkém měřítku hmotnostní spektrometrií“. Molekulární systémy biologie. 3 (1): 89. doi:10.1038 / msb4100134. PMC 1847948. PMID 17353931.
- ^ McCarty JS, Toh SY, Li P (říjen 1999). "Studie DFF45 v roli chaperonu a inhibitoru: dvě nezávislé inhibiční domény aktivity nukleázy DFF40". Sdělení o biochemickém a biofyzikálním výzkumu. 264 (1): 176–80. doi:10.1006 / bbrc.1999.1497. PMID 10527860.
- ^ Larsen BD, Rampalli S, Burns LE, Brunette S, Dilworth FJ, Megeney LA (březen 2010). „DNáza aktivovaná kaspázou 3 / kaspázou podporuje diferenciaci buněk indukcí zlomů řetězců DNA“. Sborník Národní akademie věd Spojených států amerických. 107 (9): 4230–5. Bibcode:2010PNAS..107,4230L. doi:10.1073 / pnas.0913089107. PMC 2840077. PMID 20160104.
- ^ Fernando P, Megeney LA (leden 2007). „Je kaspázově závislá apoptóza pouze extrémní diferenciací buněk?“. FASEB Journal. 21 (1): 8–17. doi:10.1096 / fj.06-5912hyp. PMID 17093139.
- ^ Fernando P, Kelly JF, Balazsi K, Slack RS, Megeney LA (srpen 2002). „Pro diferenciaci kosterního svalstva je vyžadována aktivita kaspázy 3“. Sborník Národní akademie věd Spojených států amerických. 99 (17): 11025–30. Bibcode:2002PNAS ... 9911025F. doi:10.1073 / pnas.162172899. PMC 123204. PMID 12177420.
- ^ Lai SK, Wong CH, Lee YP, Li HY (červen 2011). „Odbourávání kondenzátu Cap-H zprostředkované kaspázou-3 reguluje smrt mitotických buněk“. Buněčná smrt a diferenciace. 18 (6): 996–1004. doi:10.1038 / cdd.2010.165. PMC 3131938. PMID 21151026.
- ^ Marsden VS, O'Connor L, O'Reilly LA, Silke J, Metcalf D, Ekert PG, Huang DC, Cecconi F, Kuida K, Tomaselli KJ, Roy S, Nicholson DW, Vaux DL, Bouillet P, Adams JM, Strasser A (říjen 2002). „Apoptóza zahájená aktivací kaspázy regulovanou Bcl-2 nezávisle na apoptosomu cytochromu c / Apaf-1 / kaspázy-9“. Příroda. 419 (6907): 634–7. Bibcode:2002 Natur.419..634M. doi:10.1038 / nature01101. PMID 12374983. S2CID 4415828.
- ^ A b Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata S (leden 1998). „DNáza aktivovaná kaspázou, která degraduje DNA během apoptózy, a její inhibitor ICAD“. Příroda. 391 (6662): 43–50. Bibcode:1998 Natur.391 ... 43E. doi:10.1038/34112. PMID 9422506. S2CID 4407426.
- ^ McIlroy D, Sakahira H, Talanian RV, Nagata S (srpen 1999). „Zapojení DNázy aktivované kaspázou 3 do štěpení internukleozomální DNA vyvolané různými apoptotickými podněty“. Onkogen. 18 (31): 4401–8. doi:10.1038 / sj.onc.1202868. PMID 10442630.
Další čtení
- Indukce apoptózy (Video). Věnec věnec / Youtube. 2009. Z Murphy K, Travers P, Waldport M, Ehrenstein M (2008). Lanewayova imunobiologie (7. vydání). New York: Garland Science. ISBN 978-0-8153-4123-9.
- Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata S (leden 1998). „DNáza aktivovaná kaspázou, která degraduje DNA během apoptózy, a její inhibitor ICAD“. Příroda. 391 (6662): 43–50. Bibcode:1998 Natur.391 ... 43E. doi:10.1038/34112. PMID 9422506. S2CID 4407426.
- Liu X, Li P, Widlak P, Zou H, Luo X, Garrard WT, Wang X (červenec 1998). „40-kDa podjednotka fragmentačního faktoru DNA indukuje fragmentaci DNA a kondenzaci chromatinu během apoptózy“. Sborník Národní akademie věd Spojených států amerických. 95 (15): 8461–6. Bibcode:1998PNAS ... 95.8461L. doi:10.1073 / pnas.95.15.8461. PMC 21098. PMID 9671700.
- Mukae N, Enari M, Sakahira H, Fukuda Y, Inazawa J, Toh H, Nagata S (srpen 1998). "Molekulární klonování a charakterizace DNázy aktivované lidskou kaspázou". Sborník Národní akademie věd Spojených států amerických. 95 (16): 9123–8. Bibcode:1998PNAS ... 95,9123M. doi:10.1073 / pnas.95.16.9123. PMC 21302. PMID 9689044.
- Gu J, Dong RP, Zhang C, McLaughlin DF, Wu MX, Schlossman SF (červenec 1999). „Funkční interakce DFF35 a DFF45 s nukleázou DFF40 aktivovanou fragmentací DNA kaspázou“. The Journal of Biological Chemistry. 274 (30): 20759–62. doi:10.1074 / jbc.274.30.20759. PMID 10409614.
- McCarty JS, Toh SY, Li P (říjen 1999). „Studie DFF45 v roli chaperonu a inhibitoru: dvě nezávislé inhibiční domény aktivity nukleázy DFF40“. Sdělení o biochemickém a biofyzikálním výzkumu. 264 (1): 176–80. doi:10.1006 / bbrc.1999.1497. PMID 10527860.
- McCarty JS, Toh SY, Li P (říjen 1999). "Více domén DFF45 se synergicky váže na DFF40: role štěpení kaspázy a sekvestrace aktivátorové domény DFF40". Sdělení o biochemickém a biofyzikálním výzkumu. 264 (1): 181–5. doi:10.1006 / bbrc.1999.1498. PMID 10527861.
- Lugovskoy AA, Zhou P, Chou JJ, McCarty JS, Li P, Wagner G (prosinec 1999). „Struktura řešení CIDE-N domény CIDE-B a model interakcí CIDE-N / CIDE-N v dráze fragmentace DNA apoptózy“. Buňka. 99 (7): 747–55. doi:10.1016 / S0092-8674 (00) 81672-4. PMID 10619428.
- Judson H, van Roy N, Strain L, Vandesompele J, Van Gele M, Speleman F, Bonthron DT (duben 2000). „Strukturní a mutační analýza genu kódujícího fragmentační faktor DNA 40 (nukleáza aktivovaná kaspázou), kandidátský supresorový gen pro neuroblastom“. Genetika člověka. 106 (4): 406–13. doi:10,1007 / s004390000257. PMID 10830907. S2CID 38271068.
- Otomo T, Sakahira H, Uegaki K, Nagata S, Yamazaki T (srpen 2000). "Struktura heterodimerního komplexu mezi doménami CAD CAD a ICAD". Přírodní strukturní biologie. 7 (8): 658–62. doi:10.1038/77957. PMID 10932250. S2CID 12925074.
- Durrieu F, Samejima K, Fortune JM, Kandels-Lewis S, Osheroff N, Earnshaw WC (2001). „DNA topoizomeráza IIalfa interaguje s CAD nukleázou a podílí se na kondenzaci chromatinu během apoptotického provedení“. Aktuální biologie. 10 (15): 923–6. doi:10.1016 / S0960-9822 (00) 00620-5. PMID 10959840. S2CID 17443069.
- Zhou P, Lugovskoy AA, McCarty JS, Li P, Wagner G (květen 2001). "Struktura řešení komplexu N-terminální domény DFF40 a DFF45 a vzájemná chaperonová aktivita DFF40 a DFF45". Sborník Národní akademie věd Spojených států amerických. 98 (11): 6051–5. Bibcode:2001PNAS ... 98.6051Z. doi:10.1073 / pnas.111145098. PMC 33420. PMID 11371636.
- Nie Z, Phenix BN, Lum JJ, Alam A, Lynch DH, Beckett B, Krammer PH, Sekaly RP, Badley AD (listopad 2002). „HIV-1 proteáza zpracovává prokaspázu 8 za účelem mitochondriálního uvolňování cytochromu c, štěpení kaspázy a jaderné fragmentace“. Buněčná smrt a diferenciace. 9 (11): 1172–84. doi:10.1038 / sj.cdd.4401094. PMID 12404116. S2CID 38809690.
- Hsieh SY, Liaw SF, Lee SN, Hsieh PS, Lin KH, Chu CM, Liaw YF (Leden 2003). „Aberantní transkripty DNasy (CAD) aktivované kaspázou v buňkách lidského hepatomu“. British Journal of Cancer. 88 (2): 210–6. doi:10.1038 / sj.bjc.6600695. PMC 2377037. PMID 12610505.
- Liu QL, Kishi H, Ohtsuka K, Muraguchi A (září 2003). „Protein 70 tepelného šoku váže kaspázou aktivovanou DNázu a zvyšuje její aktivitu v T-buňkách stimulovaných TCR“. Krev. 102 (5): 1788–96. doi:10.1182 / krev-2002-11-3499. PMID 12738667.
- Widlak P, Lanuszewska J, Cary RB, Garrard WT (červenec 2003). "Podjednotkové struktury a stechiometrie proteinů lidského fragmentačního faktoru DNA před a po indukci apoptózy". The Journal of Biological Chemistry. 278 (29): 26915–22. doi:10,1074 / jbc.M303807200. PMID 12748178.
- Hillman RT, Green RE, Brenner SE (2005). „Nedoceněná role pro dohled nad RNA“. Genome Biology. 5 (2): R8. doi:10.1186 / gb-2004-5-2-r8. PMC 395752. PMID 14759258.
- Bayascas JR, Yuste VJ, Solé C, Sánchez-López I, Segura MF, Perera R, Comella JX (květen 2004). "Charakterizace sestřihových variant lidské DNázy aktivované lidskou kaspázou se strukturou a funkcí CIDE-N". FEBS Dopisy. 566 (1–3): 234–40. doi:10.1016 / j.febslet.2004.04.050. PMID 15147901. S2CID 22464440.
Galerie PDB | |
|---|---|
|