Parakrinní signalizace - Paracrine signaling
Parakrinní signalizace je forma buněčná signalizace nebo komunikace mezi buňkami ve kterém buňka produkuje signál k vyvolání změn v okolních buňkách, čímž mění chování těchto buněk. Signální molekuly známé jako parakrinní faktory difundují na relativně krátkou vzdálenost (místní působení), na rozdíl od buněčné signalizace endokrinní faktory, hormony, které cestují podstatně delší vzdálenosti přes oběhový systém; interakce juxtacrinu; a autokrinní signalizace. Buňky, které produkují parakrinní faktory, je vylučují okamžitě extracelulární životní prostředí. Faktory pak putují do blízkých buněk, ve kterých gradient přijatého faktoru určuje výsledek. Přesná vzdálenost, kterou parakrinní faktory mohou cestovat, však není jistá.
Ačkoli parakrinní signalizace vyvolává v indukovaných buňkách různorodé spektrum odpovědí, většina parakrinních faktorů využívá relativně efektivní soubor receptory a cesty. Ve skutečnosti jiný orgány v těle - dokonce i mezi různými druhy - je známo, že při diferenciálním vývoji využívá podobné sady parakrinních faktorů.[1] Vysoce konzervované receptory a dráhy mohou být organizovány do čtyř hlavních rodin na základě podobných struktur: fibroblastový růstový faktor (FGF) rodina, Ježek rodina, Wnt rodina a Nadrodina TGF-β. Zahájí se vazba parakrinního faktoru na jeho příslušný receptor signální transdukce kaskády, vyvolávající různé reakce.
Parakrinní faktory vyvolávají kompetentní respondenty
Aby parakrinní faktory úspěšně vyvolaly reakci v přijímající buňce, musí mít tato buňka k dispozici příslušné receptory na buněčné membráně pro příjem signálů, také známé jako kompetentní. Reagující buňka musí mít navíc schopnost být mechanicky indukována.
Rodina růstových faktorů fibroblastů (FGF)
Ačkoli rodina parakrinních faktorů FGF má širokou škálu funkcí, hlavní zjištění podporují myšlenku, že primárně stimulují proliferaci a diferenciaci.[2][3] Pro splnění mnoha různých funkcí mohou být FGF alternativně spojeny nebo dokonce mít různé iniciační kodony k vytvoření stovek různých FGF izoformy.[4]
Jednou z nejdůležitějších funkcí receptorů FGF (FGFR) je vývoj končetin. Tato signalizace zahrnuje devět různých alternativně sestříhané izoformy receptoru.[5] Fgf8 a Fgf10 jsou dva z kritických hráčů ve vývoji končetin. Při zahájení přední končetiny a růstu končetin u myší, axiální (podélné) narážky z meziproduktu mezoderm vyrábí Tbx5, který následně signalizuje stejné mezoderm k výrobě Fgf10. Fgf10 pak signalizuje do ektoderm zahájit výrobu Fgf8, který také stimuluje produkci Fgf10. Vymazání Fgf10 výsledky u myší bez končetin.[6]
Parakrinní signalizace Fgf je navíc nezbytná pro vyvíjející se oko kuřat. The fgf8 mRNA lokalizuje se v tom, co se odlišuje v neurální sítnice z optický pohár. Tyto buňky jsou v kontaktu s vnějšími buňkami ektodermy, které se nakonec stanou čočkou.[4]
Fenotyp a přežití myší po vyřazení některých genů FGFR:[5]
| Gén vyřazení FGFR | Přežití | Fenotyp |
|---|---|---|
| Fgf1 | Životaschopný | Nejasný |
| Fgf3 | Životaschopný | Vnitřní ucho, kosterní (ocasní) diferenciace |
| Fgf4 | Smrtící | Proliferace vnitřní buňky |
| Fgf8 | Smrtící | Gastrulace defekt, vývoj CNS, vývoj končetin |
| Fgf10 | Smrtící | Vývoj více orgánů (včetně končetin, brzlíku, hypofýzy) |
| Fgf17 | Životaschopný | Vývoj mozečku |
Dráha receptorové tyrosinkinázy (RTK)
Parakrinní signalizace skrz růstové faktory fibroblastů a jeho příslušné receptory tento receptor využívá tyrosin cesta. Tato signální dráha byla velmi studována pomocí Drosophila oči a lidské rakoviny.[7]
Vazba FGF na FGFR fosforyláty volnoběh kináza a aktivuje dráhu RTK. Tato dráha začíná na povrchu buněčné membrány, kde a ligand váže se na svůj specifický receptor. Ligandy, které se vážou na RTK, zahrnují růstové faktory fibroblastů, epidermální růstové faktory, růstové faktory odvozené z krevních destiček a faktor kmenových buněk.[7] To dimerizuje transmembránový receptor na jiný RTK receptor, což způsobí autofosforylaci a následnou konformační změna z homodimerizováno receptor. Tato konformační změna aktivuje spící kinázu každé RTK na zbytku tyrosinu. Vzhledem k tomu, že se receptor rozprostírá přes membránu z extracelulárního prostředí, přes lipidová dvojvrstva, a do cytoplazma vazba receptoru na ligand také způsobuje trans fosforylaci cytoplazmatické domény receptoru.[8]
An adaptační protein (jako je SOS) rozpoznává fosforylovaný tyrosin na receptoru. Tento protein funguje jako můstek, který spojuje RTK s intermediárním proteinem (jako je GNRP) a začíná intracelulární signální kaskádu. Intermediární protein zase stimuluje Ras vázaný na GDP k aktivovanému Ras vázanému na GTP. GAP nakonec vrátí Ras do neaktivního stavu. Aktivace Ras má potenciál iniciovat tři signální dráhy pod Ras: Ras → Raf → MAP kinázová dráha, PI3 kinázová dráha a Ral dráha. Každá cesta vede k aktivaci transkripčních faktorů, které vstupují do jádra a mění genovou expresi.[9]

RTK receptor a rakovina
Ukázalo se, že parakrinní signalizace růstových faktorů mezi blízkými buňkami se zhoršuje karcinogeneze. Ve skutečnosti mohou mutantní formy jediné RTK hrát příčinnou roli u velmi odlišných typů rakoviny. Kit protoonkogen kóduje tyrosinkinázový receptor, jehož ligandem je parakrinní protein zvaný faktor kmenových buněk (SCF), který je důležitý krvetvorba (tvorba buněk v krvi).[10] Receptor Kit a související receptory tyrosinkinázy jsou ve skutečnosti inhibiční a účinně potlačují vypalování receptorů. Mutantní formy receptoru Kit, které se konstitutivně aktivují způsobem nezávislým na ligandu, se nacházejí v rozmanité škále rakovinných malignit.[11]
Cesta RTK a rakovina
Výzkum v oblasti rakovina štítné žlázy objasnil teorii, že parakrinní signalizace může pomoci při vytváření mikroprostředí nádorů. Chemokin transkripce je nadregulovaná, když je Ras ve stavu vázaném na GTP. Chemokiny se poté uvolňují z buňky a mohou se volně vázat na další blízkou buňku. Parakrinní signalizace mezi sousedními buňkami vytváří tuto smyčku pozitivní zpětné vazby. Konstitutivní transkripce upregulovaných proteinů tedy vytváří ideální prostředí pro vznik nádorů.[12] Účinně mnohonásobné vazby ligandů na receptory RTK nadměrně stimulují cestu Ras-Raf-MAPK, která nadměrně vyjadřuje the mitogenní a invazivní kapacita buněk.[13]
Jak-STAT cesta
Kromě RTK dráhy, růstové faktory fibroblastů může také aktivovat signalizační kaskádu Jak-STAT. Místo přenosu kovalentně spojených domén tyrosinkinázy tvoří receptory Jak-STAT nekovalentní komplexy s tyrosinkinázami Jak (Janus kináza ) třída. Tyto receptory se vážou pro erytropoetin (důležité pro erytropoéza ), trombopoetin (důležité pro destička formace) a interferon (důležité pro zprostředkování funkce imunitních buněk).[14]
Po dimerizaci cytokinových receptorů po navázání ligandu se Jaks navzájem transfosforylují. Výsledné fosfotyrosiny přitahují proteiny STAT. Proteiny STAT dimerizují a vstupují do jádra, aby působily jako transkripční faktory změnit genovou expresi.[14] STATS zejména transkribuje geny, které napomáhají buněčné proliferaci a přežití - například myc.[15]
Fenotyp a přežití myší po vyřazení některých genů Jak nebo STAT:[16]
| Vyřazovací gen | Přežití | Fenotyp |
|---|---|---|
| Jak1 | Smrtící | Neurologické deficity |
| Jak2 | Smrtící | Selhání v erytropoéze |
| Stat1 | Životaschopný | Lidský nanismus a kraniosynostóza syndromy |
| Stat3 | Smrtící | Tkáňově specifické fenotypy |
| Stat4 | Životaschopný | defektní diferenciace Th1 řízená IL-12, zvýšená náchylnost k intracelulárním patogenům |
Aberantní Jak-STAT dráha a kostní mutace
Dráha Jak-STAT má zásadní význam pro vývoj končetin, konkrétně pro schopnost regulovat růst kostí pomocí parakrinní signalizace cytokinů. Mutace v této cestě však byly zapleteny do těžkých forem zakrslosti: thanatoforická dysplázie (smrtící) a achondroplazmatický nanismus (životaschopný).[17] Je to způsobeno mutací v a Fgf gen způsobující předčasnou a konstitutivní aktivaci Stat1 transkripční faktor. Chondrocyt buněčné dělení je předčasně ukončeno, což má za následek smrtelný nanismus. Buňky růstových destiček kostí žeber a končetin nejsou přepsány. Neschopnost hrudního koše se tak roztáhnout brání dýchání novorozence.[18]
Jak-STAT dráha a rakovina
Výzkum parakrinní signalizace cestou Jak-STAT odhalil jeho potenciál při aktivaci invazivního chování vaječníků epitelové buňky. Tento epitel do mezenchymální přechod je velmi patrný v metastáza.[19] Parakrinní signalizace cestou Jak-STAT je nezbytná při přechodu ze stacionárních epiteliálních buněk na mobilní mezenchymální buňky, které jsou schopné napadnout okolní tkáň. Bylo zjištěno, že pouze dráha Jak-STAT indukuje migrační buňky.[20]
Ježková rodina
The Rodina proteinů ježek podílí se na indukci buněčných typů a vytváření hranic a vzorování tkání a nachází se ve všech dvoustranných organismech. Ježkové proteiny byly poprvé objeveny a studovány v roce Drosophila. Ježkové proteiny produkují klíčové signály pro vytvoření končetiny a tělesný plán ovocných mušek i homeostáza dospělých tkání, zapojených do pozdních embryogeneze a metamorfóza. Nejméně tři ježci „Drosophila“ homology byly nalezeny u obratlovců: zvukový ježek, pouštní ježek a indický ježek. Sonic ježek (SHH ) má různé role ve vývoji obratlů, zprostředkování signalizace a regulaci organizace centrálního nervového systému, končetiny a somit polarita. Pouštní ježek (DHH ) je vyjádřen v Sertoliho buňky zahrnutý do něčeho, zůčastnit se čeho spermatogeneze. Indický ježek (IHH ) je vyjádřen ve střevech a chrupavkách, což je důležité pro postnatální růst kostí.[21][22][23]
Ježková signální dráha


Členové rodiny proteinů Ježek jednají vazbou na a transmembránový "Oprava "receptor, který je vázán na"Uhlazen "protein, kterým může být signál Ježek." transdukované. V nepřítomnosti Ježka inhibuje Patched receptor vyhlazený účinek. Inhibice vyhlazených způsobuje Cubitus interruptus (Ci), Fused a Cos proteinový komplex připojený k mikrotubulům, aby zůstaly nedotčené. V této konformaci je protein Ci rozštěpen tak, že části proteinu je umožněno vstoupit do jádra a působit jako transkripční represor. V přítomnosti Ježka Patched již neinhibuje Smoothened. Pak je aktivní vyhlazený protein schopen inhibovat PKA a Slimb, aby nedošlo k odštěpení proteinu Ci. Tento intaktní protein Ci může vstoupit do jádra, asociovat s proteinem CPB a působit jako transkripce aktivátor, indukující expresi genů Hedgehog-response.[23][24][25]
Ježková signální dráha a rakovina
Hedgehog Signaling pathway je rozhodující pro správné vzorkování a orientaci tkáně během normálního vývoje většiny zvířat. Ježkové proteiny indukují proliferace buněk v určitých buňkách a diferenciace v jiných. Aberantní aktivace dráhy Ježek byla zapletena do několika typů rakoviny, Bazocelulární karcinom zejména. Tato nekontrolovaná aktivace proteinů Hedgehog může být způsobena mutacemi signální dráhy, které by byly ligand nezávislá nebo mutace, která způsobuje nadměrná exprese proteinu Ježek, který by byl závislý na ligandu. Kromě toho se ukázalo, že aktivace dráhy Ježek vyvolané terapií je nezbytná pro progresi nádorů prostaty po androgenní deprivační terapie.[26] Toto spojení mezi signální cestou Hedgehog a lidskými rakovinami může poskytnout možnou terapeutickou intervenci jako léčbu těchto rakovin. Signální dráha Ježek je také zapojena do normální regulace kmenová buňka populace a je nutný pro normální růst a regeneraci poškozených orgánů. To může poskytnout další možnou cestu pro tumorigeneze cestou Ježek.[27][28][29]
Wnt rodina
The Wnt protein rodina zahrnuje velké množství cystein -bohatý glykoproteiny. Wnt proteiny se aktivují signální transdukce kaskády třemi různými cestami, kanonickými Wnt cesta, nekanonický dráha polarity polárních buněk (PCP) a nekanonický Wnt / Ca2+ cesta. Zdá se, že proteiny Wnt řídí širokou škálu vývojových procesů a byly považovány za nezbytné pro kontrolu vřeteno orientace, polarita buněk, adheze zprostředkovaná kadherinem a časný vývoj embryí v mnoha různých organismech. Současný výzkum ukázal, že deregulace signalizace Wnt hraje roli při tvorbě nádoru, protože na buněčné úrovni proteiny Wnt často regulovány proliferace buněk, morfologie buněk, buňka pohyblivost a osud buňky.[30]
Kanonická signální cesta Wnt

V kanonická cesta, Wnt proteiny se váží na svůj transmembránový receptor Frizzled rodina proteinů. Vazba Wnt na kadeřavý protein aktivuje Rozcuchaný protein. Ve svém aktivním stavu disheveled protein inhibuje aktivitu glykogen syntázy kinázy 3 (GSK3 ) enzym. Normálně aktivní GSK3 zabraňuje disociaci β-kateninu na APC protein, jehož výsledkem je β-katenin degradace. Takto inhibovaný GSK3 umožňuje p-katenin disociovat se z APC, akumulovat se a cestovat do jádra. V jádru se β-katenin sdružuje s Lef / Tcf transkripční faktor, který již pracuje na DNA jako represoru a inhibuje transkripci genů, na které se váže. Vazba β-kateninu na Lef / Tcf funguje jako transkripční aktivátor, který aktivuje transkripci genů reagujících na Wnt.[31][32][33]
Nekanonické signální dráhy Wnt
Nekanonické dráhy Wnt poskytují cestu přenosu signálu pro Wnt, která nezahrnuje β-katenin. V nekanonických drahách Wnt ovlivňuje aktin a mikrotubulární cytoskelet stejně jako genová transkripce.
Cesta nekanonické polarity polárních buněk (PCP)

Nekanonická dráha PCP reguluje buňku morfologie, divize, a hnutí. Opět se Wnt proteiny váží na Frizzled a aktivují jej, takže Frizzled aktivuje rozcuchaný protein, který je připoután k plazmatické membráně prostřednictvím Ostnatý protein a transmembránový protein Stbm. Aktivní Disheveled aktivuje RhoA GTPáza prostřednictvím rozcuchaného přidruženého aktivátoru služby morfogeneze 1 (Daam1) a Rac protein. Aktivní RhoA je schopen vyvolat změny cytoskeletu aktivací kinázy spojené s Roh (ROCK) a přímo ovlivnit genovou transkripci. Aktivní Rac může přímo vyvolat změny cytoskeletu a ovlivnit transkripci genů aktivací JNK.[31][32][33]
Nekanonický Wnt / Ca2+ cesta
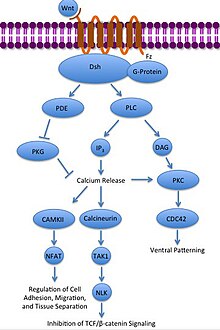
Nekanonický Wnt / Ca2+ dráha reguluje intracelulárně vápník úrovně. Opět se Wnt váže a aktivuje se na Frizzled. V tomto případě však aktivovaný Frizzled způsobí, že spojený G-protein aktivuje a fosfolipáza (PLC), který interaguje s PIP a rozděluje jej2 do DAG a IP3. IP3 se pak může vázat na receptor na endoplazmatické retikulum uvolňovat intracelulární zásoby vápníku, indukovat expresi genu závislého na vápníku.[31][32][33]
Wnt signální dráhy a rakovina
Signální dráhy Wnt jsou kritické v signalizaci buňky-buňky během normálního vývoje a embryogeneze a jsou vyžadovány pro údržbu dospělé tkáně, proto není těžké pochopit, proč narušení signálních drah Wnt může podporovat člověka degenerativní onemocnění a rakovina.
Signální dráhy Wnt jsou složité a zahrnují mnoho různých prvků, a proto mají mnoho cílů pro nesprávnou regulaci. Mutace, které způsobují konstitutivní aktivaci signální dráhy Wnt, vedou k tvorbě nádoru a rakovině. Aberantní aktivace dráhy Wnt může vést ke zvýšení buněčné proliferace. Současný výzkum je zaměřen na působení signální dráhy Wnt, regulaci volby kmenových buněk k proliferaci a obnově. Tato akce Wnt signalizace v možné kontrole a údržbě kmenových buněk může poskytnout možnou léčbu u rakovin vykazujících aberantní Wnt signalizaci.[34][35][36]
Nadrodina TGF-β
"TGF „(Transforming Growth Factor) je rodina proteinů, která zahrnuje 33 členů, které kódují dimerní, vylučované polypeptidy, které regulují vývoj.[37] Řada vývojových procesů je pod jeho kontrolou, včetně gastrulace, osové symetrie těla, morfogeneze orgánů a homeostázy tkání u dospělých.[38] Všechno TGF-p ligandy se vážou na receptory typu I nebo typu II a vytvářejí heterotetramické komplexy.[39]
Dráha TGF-β
The Dráha TGF-β reguluje mnoho buněčných procesů ve vývoji embrya a dospělých organismů, včetně růst buněk, diferenciace, apoptóza, a homeostáza. U lidí a dalších savců existuje pět druhů receptorů typu II a sedm typů receptorů typu I. Tyto receptory jsou známé jako „kinázy duální specificity“, protože jejich cytoplazmatická kinázová doména má slabou aktivitu tyrosinkinázy, ale silnou serin /threonin kinázová aktivita.[40] Když se ligand TGF-p superrodiny váže na receptor typu II, rekrutuje receptor typu I a aktivuje ho fosforylací serinových nebo threoninových zbytků ve svém boxu „GS“.[41] To vytváří aktivační komplex, který pak může fosforylovat proteiny SMAD.

Cesta SMAD
Existují tři třídy SMAD:
Příklady SMAD v každé třídě:[42][43][44]
| Třída | SMAD |
|---|---|
| R-SMAD | SMAD1, SMAD2, SMAD3, SMAD5 a SMAD8 / 9 |
| Co-SMAD | SMAD4 |
| JE ŠÍLENÝ | SMAD6 a SMAD7 |
Nadrodina TGF-β aktivuje členy SMAD rodina, která funguje jako transkripční faktory. Specificky receptor typu I, aktivovaný receptorem typu II, fosforyluje R-SMAD které se pak vážou k co-SMAD, SMAD4. R-SMAD / Co-SMAD tvoří komplex s importovat a vstupuje do jádra, kde působí transkripční faktory a buď up-regulovat nebo down-regulovat v expresi cílového genu.
Specifické ligandy TGF-β povedou k aktivaci buď SMAD2 / 3 nebo SMAD1 / 5 R-SMAD. Například kdy aktivin, Nodal nebo Ligand TGF-p váže se na receptory, fosforylovaný receptorový komplex se může aktivovat SMAD2 a SMAD3 fosforylací. Když se však ligand BMP váže na receptory, aktivuje se komplex fosforylovaného receptoru SMAD1 a SMAD5. Poté komplexy Smad2 / 3 nebo Smad1 / 5 tvoří dimerní komplex SMAD4 a stát se transkripční faktory. I když jich je mnoho R-SMAD účastnící se cesty je pouze jeden společný SMAD, SMAD4.[45]
Cesta, která není SMAD
Signální proteiny jiné než Smad přispívají k odpovědím dráhy TGF-p třemi způsoby. Nejprve signální dráhy jiné než Smad fosforylují Smady. Za druhé, Smads přímo signalizuje do jiných drah přímou komunikací s jinými signálními proteiny, jako jsou kinázy. Nakonec receptory TGF-β přímo fosforylují proteiny jiné než Smad.[46]
Členové nadrodiny TGF-β
1. Rodina TGF-p
Tato rodina zahrnuje TGF-β1, TGF-β2, TGF-p3 a TGF-p5. Podílejí se na pozitivní a negativní regulaci buněčné dělení, vznik extracelulární matrix mezi buňkami, apoptóza, a embryogeneze. Oni se vážou Receptor TGF-β typu II (TGFBRII).
TGF-β1 stimuluje syntézu kolagen a fibronektin a inhibuje degradaci extracelulární matrix. Nakonec zvyšuje produkci extracelulární matrice o epitelové buňky.[39]Proteiny TGF-β regulují epitel tím, že kontrolují, kde a kdy se rozvětvují, aby vytvořily kanály ledvin, plic a slinných žláz.[39]
2. Rodina kostních morfogenetických proteinů (BMP)
Původně bylo zjištěno, že členové rodiny BMP indukují tvorba kostí, jak napovídá jejich název. BMP jsou však velmi multifunkční a mohou také regulovat apoptóza, migrace buněk, buněčné dělení, a diferenciace. Specifikují také přední / zadní osu, indukují růst a regulují homeostáza.[37]
BMP se váží na kostní morfogenetický proteinový receptor typu II (BMPR2). Některé z proteinů BMP rodina jsou BMP4 a BMP7. BMP4 podporuje tvorbu kostí, způsobuje buněčnou smrt nebo signalizuje tvorbu pokožka, v závislosti na tkáni, na kterou působí. BMP7 je zásadní pro vývoj ledvin, syntézu spermií a polarizaci neurální trubice. Oba BMP4 a BMP7 regulují stabilitu a zpracování zralého ligandu, včetně degradace ligandů v lysozomech.[37] BMP působí rozptylováním z buněk, které je vytvářejí.[47]
Další členové nadrodiny TFG-β
- Rodina Vg1
- Rodina Activin
- Zahrnutý do něčeho, zůčastnit se čeho embryogeneze a osteogeneze
- Regulovat inzulín a hypofýza, gonadal a hypotalamus hormony
- Faktory přežití nervových buněk
- 3 Activins: Aktivin A, Aktivin B. a Activin AB.
- Gliálně derivovaný neurotrofický faktor (GDNF)
- Potřebné pro ledviny a enterický neuron diferenciace
- Müllerianův inhibiční faktor
- Podílí se na stanovení pohlaví u savců
- Nodal
- Váže se Receptor aktivinu A typu 2B
- Tvoří komplex receptorů s Receptor aktivinu A typu 1B nebo s Receptor aktivinu A typu 1C.[48]
- Faktory růstu a diferenciace (GDF)
Souhrnná tabulka signální dráhy TFG-β
| TGF Beta nadrodina ligand | Receptor typu II | Receptor typu I. | R-SMAD | Co-SMAD | Inhibitory ligandu |
|---|---|---|---|---|---|
| Aktivin A | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | Follistatin |
| GDF1 | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | |
| GDF11 | ACVR2B | ACVR1B (ALK4), TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | |
| Kostní morfogenetické proteiny | BMPR2 | BMPR1A (ALK3), BMPR1B (ALK6) | SMAD1 SMAD5, SMAD8 | SMAD4 | Člun ze dřeva, Chordin, DAN |
| Nodal | ACVR2B | ACVR1B (ALK4), ACVR1C (ALK7) | SMAD2, SMAD3 | SMAD4 | Levičák |
| TGFβs | TGFpRII | TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | LTBP1, THBS1, Decorin |
Příklady
Růstový faktor a koagulační faktory jsou parakrinní signalizační prostředky. Místní působení signalizace růstového faktoru hraje obzvláště důležitou roli ve vývoji tkání. Taky, kyselina retinová, aktivní forma vitamin A., funguje parakrinním způsobem k regulaci genové exprese během embryonálního vývoje u vyšších zvířat.[49] U hmyzu Allatostatin řídí růst parakrinním působením na corpora allata.[Citace je zapotřebí ]
U dospělých organismů je parakrinní signalizace zapojena do odpovědí na alergeny, oprava tkáně, tvorba jizva a krev srážení.[Citace je zapotřebí ]
Viz také
- cAMP závislá cesta
- Buněčná komunikace (biologie)
- Přeslech (biologie)
- Signální dráha JAK-STAT
- Lipidová signalizace
- Místní hormon - buď parakrinní hormon, nebo hormon působící parakrinně i endokrinně
- Signální dráha MAPK
- Netpath - Vybraný zdroj drah přenosu signálu u lidí
- Parakrinní regulátor
Reference
- ^ "Parakrinní faktory". Citováno 27. července 2018.
- ^ Gospodarowicz, D .; Ferrara, N .; Schweigerer, L .; Neufeld, G. (1987). "Strukturální charakterizace a biologické funkce růstového faktoru fibroblastů". Endokrinní hodnocení. 8 (2): 95–114. doi:10.1210 / edrv-8-2-95. PMID 2440668.
- ^ Rifkin, Daniel B .; Moscatelli, David (1989). „Poslední vývoj v buněčné biologii základního růstového faktoru fibroblastů“. The Journal of Cell Biology. 109 (1): 1–6. doi:10.1083 / jcb.109.1.1. JSTOR 1613457. PMC 2115467. PMID 2545723.
- ^ A b Lappi, Douglas A. (1995). "Cílení na nádor prostřednictvím receptorů růstových faktorů fibroblastů". Semináře z biologie rakoviny. 6 (5): 279–88. doi:10.1006 / scbi.1995.0036. PMID 8562905.
- ^ A b Xu, J .; Xu, J; Colvin, JS; McEwen, DG; MacArthur, CA; Coulier, F; Gao, G; Goldfarb, M (1996). „Specifičnost receptoru rodiny fibroblastových růstových faktorů“. Journal of Biological Chemistry. 271 (25): 15292–7. doi:10.1074 / jbc.271.25.15292. PMID 8663044.
- ^ Logan, M. (2003). „Prst nebo prst: Molekulární základ identity končetiny“. Rozvoj. 130 (26): 6401–10. doi:10,1242 / dev.00956. PMID 14660539.
- ^ A b Fantl, Wendy J; Johnson, Daniel E; Williams, Lewis T (1993). "Signalizace receptorovými tyrosinkinázami". Roční přehled biochemie. 62: 453–81. doi:10.1146 / annurev.bi.62.070193.002321. PMID 7688944.
- ^ Yarden, Yosef; Ullrich, Axel (1988). "Tyrosin kinázy receptoru růstového faktoru". Roční přehled biochemie. 57: 443–78. doi:10.1146 / annurev.bi.57.070188.002303. PMID 3052279.
- ^ Katz, Michael E; McCormick, Frank (1997). "Transdukce signálu z více Ras efektorů". Aktuální názor na genetiku a vývoj. 7 (1): 75–9. doi:10.1016 / S0959-437X (97) 80112-8. PMID 9024640.
- ^ Zsebo, Krisztina M .; Williams, David A .; Geissler, Edwin N .; Broudy, Virginie C .; Martin, Francis H .; Atkins, Harry L .; Hsu, Rou-Yin; Birkett, Neal C .; Okino, Kenneth H .; Murdock, Douglas C .; Jacobsen, Frederick W .; Langley, Keith E .; Smith, Kent A .; Takeish, Takashi; Cattanach, Bruce M .; Galli, Stephen J .; Suggs, Sidney V. (1990). „Faktor kmenových buněk je kódován na SI lokusu myši a je ligandem pro tyrosinkinázový receptor c-kit.“ Buňka. 63 (1): 213–24. doi:10.1016 / 0092-8674 (90) 90302-U. PMID 1698556.
- ^ Rönnstrand, L. (2004). "Transdukce signálu prostřednictvím receptoru faktoru kmenových buněk / c-Kit". Buněčné a molekulární biologické vědy. 61 (19–20): 2535–48. doi:10.1007 / s00018-004-4189-6. PMID 15526160.
- ^ Melillo, Rosa Marina; Castellone, Maria Domenica; Guarino, Valentina; De Falco, Valentina; Cirafici, Anna Maria; Salvatore, Giuliana; Caiazzo, Fiorina; Basolo, Fulvio; Giannini, Riccardo; Kruhoffer, Mogens; Orntoft, Torben; Fusco, Alfredo; Santoro, Massimo (2005). „Lineární signální kaskáda RET / PTC-RAS-BRAF zprostředkovává pohyblivý a mitogenní fenotyp buněk rakoviny štítné žlázy“. Journal of Clinical Investigation. 115 (4): 1068–81. doi:10,1172 / JCI22758. PMC 1062891. PMID 15761501.
- ^ Kolch, Walter (2000). „Smysluplné vztahy: Regulace dráhy Ras / Raf / MEK / ERK proteinovými interakcemi“. The Biochemical Journal. 351 (2): 289–305. doi:10.1042/0264-6021:3510289. PMC 1221363. PMID 11023813.
- ^ A b Aaronson, David S .; Horvath, Curt M. (2002). „Cestovní mapa pro ty, kteří neznají JAK-STAT“. Věda. 296 (5573): 1653–5. Bibcode:2002Sci ... 296.1653A. doi:10.1126 / science.1071545. PMID 12040185.
- ^ Rawlings, Jason S .; Rosler, Kristin M .; Harrison, Douglas A. (2004). „Signální cesta JAK / STAT“. Journal of Cell Science. 117 (8): 1281–3. doi:10.1242 / jcs.00963. PMID 15020666.
- ^ O'Shea, John J; Gadina, Massimo; Schreiber, Robert D (2002). „Cytokinová signalizace v roce 2002: nová překvapení v dráze Jak / Stat“. Buňka. 109 (2): S121–31. doi:10.1016 / S0092-8674 (02) 00701-8. PMID 11983158.
- ^ Shiang, Rita; Thompson, Leslie M .; Zhu, Ya-Zhen; Church, Deanna M .; Fielder, Thomas J .; Bocian, Maureen; Winokur, Sara T .; Wasmuth, John J. (1994). „Mutace v transmembránové doméně FGFR3 způsobují nejběžnější genetickou formu nanismu, achondroplázii.“ Buňka. 78 (2): 335–42. doi:10.1016/0092-8674(94)90302-6. PMID 7913883.
- ^ Kalluri, Raghu; Weinberg, Robert A. (2009). "Základy epiteliálně-mezenchymálního přechodu". Journal of Clinical Investigation. 119 (6): 1420–8. doi:10,1172 / JCI39104. PMC 2689101. PMID 19487818.
- ^ Silver, Debra L .; Montell, Denise J. (2001). „Parakrinní signalizace cestou JAK / STAT aktivuje invazivní chování ovariálních epiteliálních buněk v Drosophile“. Buňka. 107 (7): 831–41. doi:10.1016 / S0092-8674 (01) 00607-9. PMID 11779460.
- ^ Ingham, P. W .; McMahon, AP (2001). „Ježková signalizace ve vývoji zvířat: paradigmata a principy“. Geny a vývoj. 15 (23): 3059–87. doi:10,1101 / gad. 938601. PMID 11731473.
- ^ Bitgood, Mark J .; McMahon, Andrew P. (1995). "Ježek a Bmp Geny jsou koexprimovány na mnoha různých místech interakce buňka-buňka v myším embryu “. Vývojová biologie. 172 (1): 126–38. doi:10.1006 / dbio.1995.0010. PMID 7589793.
- ^ A b Jacob, L .; Lum, L. (2007). "Signalizační cesta ježka". Věda STKE. 2007 (407): cm6. doi:10.1126 / št. 4072007cm6. PMID 17925577.
- ^ Johnson, Ronald L; Scott, Matthew P (1998). "Noví hráči a hlavolamy v signální dráze Ježek". Aktuální názor na genetiku a vývoj. 8 (4): 450–6. doi:10.1016 / S0959-437X (98) 80117-2. PMID 9729722.
- ^ Nybakken, K; Perrimon, N (2002). "Transdukce signálu ježka: nedávné nálezy". Aktuální názor na genetiku a vývoj. 12 (5): 503–11. doi:10.1016 / S0959-437X (02) 00333-7. PMID 12200154.
- ^ Lubik AA, Nouri M, Truong S, Ghaffari M, Adomat HH, Corey E, Cox ME, Li N, Guns ES, Yenki P, Pham S, Buttyan R (2016). „Signalizace parakrinního sonického ježka významně přispívá k získané steroidogenezi v mikroprostředí nádoru prostaty“. International Journal of Cancer. 140 (2): 358–369. doi:10.1002 / ijc.30450. PMID 27672740.
- ^ Collins, R. T .; Cohen, SM (2005). „Genetická obrazovka v Drosophile pro identifikaci nových složek signální dráhy ježka“. Genetika. 170 (1): 173–84. doi:10.1534 / genetika.104.039420. PMC 1449730. PMID 15744048.
- ^ Evangelista, M .; Tian, H .; De Sauvage, F. J. (2006). „Ježková signální dráha rakoviny“. Klinický výzkum rakoviny. 12 (20): 5924–8. doi:10.1158 / 1078-0432.CCR-06-1736. PMID 17062662.
- ^ Taipale, Jussi; Beachy, Philip A. (2001). "Ježkové a Wnt signální dráhy v rakovině". Příroda. 411 (6835): 349–54. Bibcode:2001 Natur.411..349T. doi:10.1038/35077219. PMID 11357142.
- ^ Cadigan, K. M .; Nusse, R. (1997). „Signalizace Wnt: Společné téma ve vývoji zvířat“. Geny a vývoj. 11 (24): 3286–305. doi:10.1101 / gad.11.24.3286. PMID 9407023.
- ^ A b C Dale, Trevor C. (1998). "Přenos signálu pomocí rodiny ligandů Wnt". The Biochemical Journal. 329 (Pt 2): 209–23. doi:10.1042 / bj3290209. PMC 1219034. PMID 9425102.
- ^ A b C Chen, Xi; Yang, červen; Evans, Paul M; Liu, Chunming (2008). „Wnt signaling: the good and bad“. Acta Biochimica et Biophysica Sinica. 40 (7): 577–94. doi:10.1111 / j.1745-7270.2008.00440.x. PMC 2532600. PMID 18604449.
- ^ A b C Komiya, Yuko; Habas, Raymond (2008). "Wnt signální transdukční cesty". Organogeneze. 4 (2): 68–75. doi:10,4161 / org. 4.2.5851. PMC 2634250. PMID 19279717.
- ^ Logan, Catriona Y .; Nusse, Roel (2004). "Cesta signalizace Wnt ve vývoji a nemoci". Roční přehled buněčné a vývojové biologie. 20: 781–810. CiteSeerX 10.1.1.322.311. doi:10.1146 / annurev.cellbio.20.010403.113126. PMID 15473860.
- ^ Lustig, B; Behrens, J (2003). "Signální dráha Wnt a její role ve vývoji nádoru". Journal of Cancer Research and Clinical Oncology. 129 (4): 199–221. doi:10.1007 / s00432-003-0431-0. PMID 12707770.
- ^ Neth, Peter; Ries, Christian; Karow, Marisa; Egea, Virginie; Ilmer, Matthias; Jochum, Marianne (2007). „Cesta přenosu signálu Wnt v kmenových buňkách a rakovinných buňkách: vliv na buněčnou invazi“. Recenze kmenových buněk. 3 (1): 18–29. doi:10.1007 / s12015-007-0001-r. PMID 17873378.
- ^ A b C Bandyopadhyay, Amitabha; Tsuji, Kunikazu; Cox, Karen; Harfe, Brian D .; Rosen, Vicki; Tabin, Clifford J. (2006). „Genetická analýza rolí BMP2, BMP4 a BMP7 ve vzorcích končetin a skeletogenezi“. Genetika PLOS. 2 (12): e216. doi:10.1371 / journal.pgen.0020216. PMC 1713256. PMID 17194222.
- ^ Attisano, Liliana; Wrana, Jeffrey L. (2002). "Transdukce signálu nadrodinou TGF-β". Věda. 296 (5573): 1646–7. Bibcode:2002Sci ... 296.1646A. doi:10.1126 / science.1071809. PMID 12040180.
- ^ A b C Wrana, Jeffrey L .; Ozdamar, Barish; Le Roy, Christine; Benchabane, Hassina (2008). "Signální receptory rodiny TGF-β". V Derynck, Rik; Miyazono, Kohei (eds.). Rodina TGF-β. str. 151–77. ISBN 978-0-87969-752-5.
- ^ ten Dijke, Peter; Heldin, Carl-Henrik (2006). „Rodina Smadů“. V deseti Dijke, Peter; Heldin, Carl-Henrik (eds.). Smad Signal Transduction: Smads v šíření, diferenciaci a nemoci. Regulace proteinů a buněk. 5. Dordrecht: Springer. s. 1–13. ISBN 978-1-4020-4709-1.
- ^ Moustakas, Aristidis (01.09.2002). "Signální síť Smad". Journal of Cell Science. 115 (17): 3355–6. PMID 12154066.
- ^ Wu, Jia-Wei; Hu, Min; Chai, Jijie; Seoane, Joan; Huse, Morgan; Li, Carey; Rigotti, Daniel J .; Kyin, Saw; Muir, Tom W .; Fairman, Robert; Massagué, Joan; Shi, Yigong (2001). "Krystalová struktura fosforylovaného Smad2". Molekulární buňka. 8 (6): 1277–89. doi:10.1016 / S1097-2765 (01) 00421-X. PMID 11779503.
- ^ Pavletich, Nikola P .; Hata, Yigong; Lo, Akiko; Massagué, Roger S .; Pavletich, Joan (1997). "Strukturální základ pro mutační inaktivaci nádorového supresoru Smad4". Příroda. 388 (6637): 87–93. Bibcode:1997 Natur.388R..87S. doi:10.1038/40431. PMID 9214508.
- ^ Itoh, Fumiko; Asao, Hironobu; Sugamura, Kazuo; Heldin, Carl-Henrik; Ten Dijke, Peter; Itoh, Susumu (2001). „Podpora signalizace kostního morfogenetického proteinu prostřednictvím negativní regulace inhibičních Smadů“. Časopis EMBO. 20 (15): 4132–42. doi:10.1093 / emboj / 20.15.4132. PMC 149146. PMID 11483516.
- ^ Schmierer, Bernhard; Hill, Caroline S. (2007). „Transdukce signálu TGFβ – SMAD: Molekulární specificita a funkční flexibilita“. Nature Reviews Molecular Cell Biology. 8 (12): 970–82. doi:10.1038 / nrm2297. PMID 18000526.
- ^ Moustakas, Aristidis; Heldin, Carl-Henrik (2005). „Nesmadové signály TGF-β“. Journal of Cell Science. 118 (16): 3573–84. doi:10.1242 / jcs.02554. PMID 16105881.
- ^ Ohkawara, Bisei; Iemura, Shun-Ichiro; Ten Dijke, Peter; Ueno, Naoto (2002). „Akční rozsah BMP je definován základním aminokyselinovým jádrem N-terminálu“. Aktuální biologie. 12 (3): 205–9. doi:10.1016 / S0960-9822 (01) 00684-4. PMID 11839272.
- ^ Munir, Sadia; Xu, Guoxiong; Wu, Yaojiong; Yang, Burton; Lala, Peeyush K .; Peng, Chun (2004). „Nodal a ALK7 inhibují šíření a indukují apoptózu v lidských trofoblastových buňkách“. Journal of Biological Chemistry. 279 (30): 31277–86. doi:10,1074 / jbc.M400641200. PMID 15150278.
- ^ Duester, Gregg (září 2008). „Syntéza a signalizace kyseliny retinové během časné organogeneze“. Buňka. 134 (6): 921–31. doi:10.1016 / j.cell.2008.09.002. PMC 2632951. PMID 18805086.
externí odkazy
- Parakrin + signalizace v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- "parakrin " na Dorlandův lékařský slovník