Gastrulace - Gastrulation
| Gastrulace | |
|---|---|
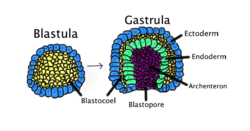 Gastrulace nastane, když se blastula, složená z jedné vrstvy, sklopí dovnitř a zvětší se, aby vytvořila gastrulu. Tento diagram je barevně odlišen: ektoderm, modrý; endoderm, zelená; blastocoel (žloutkový pytel), žlutý; a archenteron (střeva), fialová. | |
| Identifikátory | |
| Pletivo | D054262 |
| Anatomická terminologie | |
v vývojová biologie, gastrulace je fáze brzy v embryonální vývoj většiny zvířata, během kterého blastula (jednovrstvá dutá koule z buňky ) je reorganizována do vícevrstvé struktury známé jako gastrula. Před gastrulací embryo je spojitý epiteliální list buněk; na konci gastrulace embryo začalo diferenciace vytvořit odlišné buněčné linie, nastavit základní osy těla (např. hřbetní-ventrální, předozadní ) a internalizoval jeden nebo více typů buněk včetně potenciálních střevo.
v triploblastický organismy je gastrula trilaminární („třívrstvá“). Tyto tři zárodečné vrstvy jsou známé jako ektoderm (vnější vrstva), mezoderm (střední vrstva) a endoderm (vnitřní vrstva).[1][2] v diploblastický organismy, jako jsou Cnidaria a Ctenophora, gastrula má pouze ektoderm a endoderm. Tyto dvě vrstvy jsou také někdy označovány jako hypoblast a epiblast.[3]
Gastrulace probíhá po výstřih a vznik blastula. Po gastrulaci následuje organogeneze, když je to individuální orgány rozvíjet v nově vytvořených zárodečných vrstvách.[4] Každá vrstva dává vzniknout specifickým papírové kapesníky a orgány ve vyvíjejícím se embryu.
- Ektoderm dává vzniknout pokožka, nervový systém, a do neurální lišta u obratlovců.
- Endoderm vede k epitel z zažívací ústrojí a dýchací systém a orgány spojené s zažívací ústrojí, tak jako játra a slinivka břišní.
- Mesoderm vede k mnoha buněčným typům, jako jsou sval, kost, a pojivová tkáň. U obratlovců zahrnují mesodermové deriváty notochord, srdce, krev a cévy, chrupavka z žebra a obratel a dermis.[5]
Po gastrulaci jsou buňky v těle buď uspořádány do listů spojených buněk (jako v epitel ), nebo jako síť izolovaných buněk, jako je mezenchymu.[2][6]
Ačkoli gastrulační vzorce vykazují v živočišné říši obrovské rozdíly, jsou sjednoceny pět základních typů buněčných pohybů[7] které se vyskytují během gastrulace:
Pojmy „gastrula“ a „gastrulace“ byly vytvořeny Ernst Haeckel, ve své práci z roku 1872 "Biologie vápenatých hub".[8] Lewis Wolpert, průkopnický vývojový biolog v oboru, byl oceněn za to, že poznamenal, že „není to narození, manželství nebo smrt, ale gastrulace je skutečně nejdůležitější dobou ve vašem životě.“[9]

Klasické modelové systémy
Gastrulace je v živočišné říši velmi variabilní, ale má základní podobnosti. Gastrulace byla studována u mnoha zvířat, ale některé modely byly používány déle než jiné. Dále je snazší studovat vývoj u zvířat, která se vyvíjejí mimo matku. Zvířata, jejichž gastrulace je chápána nejpodrobněji, zahrnují:
Protostomy versus deuterostomy
The rozdíl mezi protostomy a deuterostomy je založen na směru, kterým se vyvíjí ústa (stoma) ve vztahu k blastopore. Protostome pochází z řeckého slova protostoma, což znamená „první ústa“ (πρώτος + στόμα), zatímco etymologie Deuterostome je „druhá ústa“ ze slov druhý a ústa (δεύτερος + στόμα).
Hlavní rozdíly mezi deuterostomy a protostomy se nacházejí v embryonálním vývoji:
- Ústa / konečník
- Výstřih
- Protostomy mají to, co je známé jako spirální štěpení který je určit, což znamená, že osud buněk je určen při jejich formování.
- Deuterostomy mají to, co je známé jako radiální štěpení to je neurčitý.
Mořští ježci
Mořští ježci Euechinoidea jsou důležitým modelovým systémem ve vývojové biologii od 19. století.[10] Jejich gastrulace je často považována za archetyp pro bezobratlé deuterostomy.[11] K získání znalostí o gastrulaci u mořského ježka byly použity experimenty spolu s počítačovými simulacemi. Nedávné simulace zjistily, že polarita rovinných buněk je dostatečná pro řízení gastrulace mořského ježka.[12]
Stanovení zárodečné vrstvy
Mořští ježci vykazují vysoce stereotypní vzory štěpení a buněčné osudy. Mateřsky uloženo mRNA zřídit organizační centrum embrya mořského ježka. Kanonický Wnt a Delta zářez signalizace progresivně odděluje progresivní endoderm a mezoderm.[13]
Internalizace buněk
v mořští ježci první buňky k internalizaci jsou primární mezenchymu buňky (PMC), které mají a kostogenní osud, který pronikne během fáze blastuly. Gastrulace - internalizace potenciálního zákazníka endoderm a neskeletogenní mezoderm - začíná krátce nato invaginací a dalšími buněčnými přestavbami rostlinného pólu, které přispívají přibližně 30% ke konečnému archenteron délka. The konečná délka střeva závisí na přeskupení buněk v archenteronu.[14]
Obojživelníci
The žába, Xenopus byl použit jako modelový organismus pro studium gastrulace.
Symetrie lámání
Spermie přispívá jedním ze dvou mitotické astry potřebné k dokončení prvního štěpení. Spermie může vstoupit kdekoli ve zvířeti do poloviny vajíčka, ale její přesný bod vstupu rozbije radiální symetrii vajíčka uspořádáním cytoskelet. Před prvním štěpením se kůra vajíčka otáčí vzhledem k vnitřnímu cytoplazma koordinovanou činností mikrotubuly, v procesu známém jako kortikální rotace. Toto posunutí přináší do styku mateřsky zatížené determinanty buněčného osudu z rovníkové cytoplazmy a rostlinné kůry a společně tyto determinanty nastavují organizátor. Organizátorem se tedy stane oblast na rostlinné straně naproti vstupu do spermatu.[15] Hilde Mangold, pracující v laboratoři Hans Spemann, prokázali, že tento speciální „organizátor“ embrya je nezbytný a dostatečný k vyvolání gastrulace.[16][17][18]
Stanovení zárodečné vrstvy
Specifikace endodermu závisí na přeskupení determinantů uložených v matce, což vede k nuklearizaci Beta-katenin. Mesoderm je indukovaný signalizací z předpokládaného endodermu do buněk, které by se jinak staly ektodermem.[15]
Internalizace buněk
The hřbetní ret blastoporu je mechanickým pohonem gastrulace. První známka invaginace viděná v toto video gastrulace žab je hřbetní ret.
Buněčná signalizace
V žábě Xenopus, jedním ze signálů je kyselina retinová (RA).[19] Signalizace RA v tomto organismu může ovlivnit tvorbu endodermu a v závislosti na načasování signalizace může určit osud, zda jeho pankreatický, intestinální nebo respirační. Další signály, jako jsou Wnt a BMP, hrají také roli v respiračním osudu Xenopus aktivací sledovačů buněčné linie.[19]
Amniotes
Přehled
v amniotes (plazi, ptáci a savci), gastrulace zahrnuje vytvoření blastoporu, ústí do archenteron. Všimněte si, že blastopore není otvor do blastocoel, prostor uvnitř blastula, ale představuje nové zasunutí, které tlačí existující povrchy blastuly k sobě. v amniotes, gastrulace probíhá v následující sekvenci: (1) the embryo se stává asymetrický; (2) primitivní pruh formuláře; (3) buňky z epiblast na primitivní pruh podstoupit přechod z epitelu na mezenchym a vniknutí na primitivní pruh tvořit zárodečné vrstvy.[5]
Symetrie lámání
Při přípravě na gastrulaci musí být embryo asymetrické podél obou proximální-distální osa a přední a zadní osa. Proximální-distální osa se vytvoří, když buňky embrya vytvoří „vaječný válec“, který se skládá z extraembryonálních tkání, které vytvářejí struktury jako placenta, na proximálním konci a epiblast na distálním konci. Mnoho signálních drah přispívá k této reorganizaci, včetně BMP, FGF, uzlový, a Wnt. Viscerální endoderm obklopuje epiblast. The distální viscerální endoderm (DVE) migruje na přední část embrya, tvořící „přední viscerální endoderm“ (AVE). To narušuje předozadní symetrii a je regulováno uzlový signalizace.[5]
Stanovení zárodečné vrstvy
The primitivní pruh se tvoří na začátku gastrulace a nachází se na křižovatce mezi extraembryonální tkání a epiblast na zadní straně embrya a místě ingrese.[20] Formace primitivní pruh je odkázán na uzlový signalizace[5] v Kollerův srp v buňkách přispívajících k primitivnímu pruhu a BMP4 signalizace z extraembryonální tkáně.[20][21] Dále Cer1 a Levý1 omezit primitivní pruh na vhodné místo antagonizací uzlový signalizace.[22] Oblast definovaná jako primitivní pruh nadále roste směrem k distální špičce.[5]
Během raných fází vývoje je primitivní pruh strukturou, která se vytvoří bilaterální symetrie, určit místo gastrulace a zahájit tvorbu zárodečné vrstvy. Aby vytvořili pruh, plazi, ptáci a savci uspořádají mezenchymální buňky podél budoucí středové linie a vytvářejí první embryonální osu, stejně jako místo, kde budou buňky vnikat a migrovat během procesu gastrulace a tvorby zárodečné vrstvy.[23] Primitivní pruh prochází touto středovou čarou a vytváří předozadní osu těla,[24] stávat se první událostí narušující symetrii v embryo, a znamená začátek gastrulace.[25] Tento proces zahrnuje ingresi předků mezodermu a endodermu a jejich migraci do jejich konečné polohy,[24][26] kde se budou diferencovat do tří zárodečných vrstev.[23] Lokalizace buněčné adheze a signální molekuly beta-katenin je rozhodující pro správnou tvorbu oblasti organizátoru, která je zodpovědná za zahájení gastrulace.
Internalizace buněk
Aby se buňky mohly přesunout z epitel z epiblast skrz primitivní pruh k vytvoření nové vrstvy musí buňky podstoupit přechod z epitelu na mezenchym (EMT) ztratit své epiteliální vlastnosti, jako např buněčná adheze. FGF pro správnou EMT je nutná signalizace. FGFR1 je zapotřebí pro vyšší regulaci SNAI1, který dolů reguluje E-kadherin, což způsobuje ztrátu buněčné adheze. Po EMT, buňky vniknutí skrz primitivní pruh a rozložit se tak, aby vytvořily novou vrstvu buněk nebo se spojily s existujícími vrstvami. FGF8 se podílí na procesu tohoto rozptýlení od primitivní pruh.[22]
Buněčná signalizace
Existují určité signály, které hrají roli při určování a tvorbě tří zárodečných vrstev, jako jsou FGF, RA a Wnt.[19] U savců, jako jsou myši, může hrát při tvorbě plic roli signalizace RA. Pokud není dostatek RA, dojde k chybě ve výrobě plic. RA také reguluje respirační kompetenci v tomto modelu myši.
Buněčná signalizace vede k gastrulaci
Během gastrulace se buňky diferencují na ektoderm nebo mesendoderm, který se poté dělí na mezoderm a endoderm.[19] Endoderm a mezoderm se tvoří díky nodální signalizaci. Uzlová signalizace používá ligandy, které jsou součástí rodiny TGFp. Tyto ligandy budou signalizovat transmembránové receptory serin / threonin kinázy, a to pak fosforyluje Smad2 a Smad3. Tento protein se poté připojí k Smad4 a přemístí se do jádra, kde se začnou přepisovat geny mesendodermu. Dráha Wnt spolu s β-kateninem hraje klíčovou roli v nodální signalizaci a tvorbě endodermu.[27] Fibroblastové růstové faktory (FGF), kanonická dráha Wnt, kostní morfogenetický protein (BMP) a kyselina retinová (RA) jsou důležité při tvorbě a vývoji endodermy.[19] FGF jsou důležité při produkci genu homeoboxu, který reguluje časný anatomický vývoj. Signalizace BMP hraje roli v játrech a podporuje jaterní osud. Signalizace RA také indukuje geny homeoboxu, jako jsou Hoxb1 a Hoxa5. U myší, pokud chybí RA signalizace, se u myši nevyvinou plíce.[19] Signalizace RA má také mnohostranné použití při tvorbě orgánů hltanových oblouků, předního a zadního střeva.[19]
Gastrulace in vitro
Existuje řada pokusů porozumět procesům gastrulace pomocí in vitro techniky paralelní a doplňkové ke studiím na embryích, obvykle při použití 2D[28][29][30] a 3D buňka (Embryonální organoidy ) kultivační techniky[31][32][33][34] použitím Embryonální kmenové buňky (ESC) nebo indukované pluripotentní kmenové buňky (iPSC). Ty jsou spojeny s řadou jasných výhod při používání protokolů založených na tkáňové kultuře, z nichž některé zahrnují snížení nákladů na související in vivo práce (čímž se omezí, nahradí a zdokonalí používání zvířat v experimentech; 3R ), kteří jsou schopni přesně aplikovat agonisty / antagonisty prostorově a časově specifickým způsobem[32][33] což může být technicky obtížné provést během Gastrulation. Je však důležité spojit pozorování v kultuře s procesy probíhajícími v embryu pro kontext.
Pro ilustraci vedla řízená diferenciace myších ESC ke generování primitivní pruh -jako buňky, které zobrazují mnoho charakteristik buněk epiblastů, které procházejí primitivním pruhem[28] (např. přechodné brachyury nahoru regulace a buněčné změny spojené s přechod z epitelu na mezenchym[28]) a lidské ESC kultivované na mikroskopických vzorcích, ošetřené BMP4, může generovat prostorový diferenciační vzor podobný uspořádání zárodečné vrstvy v lidském embryu.[29][30] Nakonec pomocí 3D embryoidní tělo - a organoid - techniky založené na malých agregátech myších ESC (Embryonální organoidy nebo gastruloidy ) jsou schopni ukázat řadu procesů časného vývoje embryí savců, jako je rozbití symetrie, polarizace genové exprese, pohyby podobné gastrulaci, axiální prodloužení a tvorba všech tří embryonálních os (anteroposteriorní, dorsoventrální a levo-pravá osa) .[31][32][33][35]
Viz také
Reference
Poznámky
- ^ Mundlos 2009: str. 422
- ^ A b McGeady, 2004: str. 34
- ^ Jonathon M.W., Slack (2013). Základní vývojová biologie. West Sussex, Velká Británie: Wiley-Blackwell. str. 122. ISBN 978-0-470-92351-1.
- ^ Hall, 1998: 132-134
- ^ A b C d E Arnold & Robinson, 2009
- ^ Hall, 1998: str. 177
- ^ Gilbert 2010: str. 164.
- ^ Ereskovsky 2010: str. 236
- ^ Wolpert L. (2008) Triumf embrya. Courier Corporation, strana 12. ISBN 9780486469294
- ^ Laubichler, M.D. a Davidson, E. H. (2008). „Boveriho dlouhý experiment: merogony mořského ježka a stanovení úlohy jaderných chromozomů ve vývoji“. "Vývojová biologie". „314 (1): 1-11“. „doi: 10.1016 / j.ydbio.2007.11.024“.
- ^ McClay, David R .; Gross, J.M .; Range, Ryan; Peterson, R.E .; Bradham, Cynthia (2004). „Kapitola 9: Gastrulace mořských ježků“. V Stern, Claudio D. (ed.). Gastrulace: Od buněk po embrya. Cold Spring Harbor Laboratory Press. str. 123–137. ISBN 978-0-87969-707-5.
- ^ Nielsen, Bjarke Frost; Nissen, Silas Boye; Sneppen, Kim; Mathiesen, Joachim; Trusina, Ala (21. února 2020). „Model propojující tvar a polaritu buněk s organogenezí“. iScience. 23 (2): 100830. Bibcode:2020iSci ... 23j0830N. doi:10.1016 / j.isci.2020.100830. PMC 6994644. PMID 31986479. S2CID 210934521.
- ^ McClay, D. R. 2009. Štěpení a gastrulace v mořském ježku. eLS. doi:10.1002 / 9780470015902.a0001073.pub2
- ^ Hardin J. D. (1990). „Kontextové chování buněk během gastrulace“ (PDF). Semin. Dev. Biol. 1: 335–345.
- ^ A b Gilbert, Scott F. (2000). „Tvorba os u obojživelníků: Fenomén organizátora, postupné stanovení os obojživelníků“. Vývojová biologie. Sinauer Associates.
- ^ Gilbert, Scott F. (2000). „Obrázek 10.20, [Organizace sekundární osy ...]“. www.ncbi.nlm.nih.gov. Citováno 1. června 2020.
- ^ Spemann H., Mangold H. (1924). „Über Induktion von Embryonanlagen durch Implantation artfremder Organisatoren“. Roux 'Arch. F. Entw. Mech. 100 (3–4): 599–638. doi:10.1007 / bf02108133. S2CID 12605303.
- ^ De Robertis Edward (2006). „Spemannův organizátor a samoregulace v embryích obojživelníků“. Nature Reviews Molecular Cell Biology. 7 (4): 296–302. doi:10.1038 / nrm1855. PMC 2464568. PMID 16482093.
- ^ A b C d E F G Zorn A, Wells J (2009). „Vývoj endodermu obratlovců a tvorba orgánů“. Annu Rev Cell Dev Biol. 25: 221–251. doi:10.1146 / annurev.cellbio.042308.113344. PMC 2861293. PMID 19575677.
- ^ A b Tam & Behringer, 1997
- ^ Catala, 2005: str. 1535
- ^ A b Tam, P.P .; Loebel, D.A (2007). "Genová funkce v myší embryogenezi: připravte se na gastrulaci". Nat Rev Genet. 8 (5): 368–81. doi:10.1038 / nrg2084. PMID 17387317. S2CID 138874.
- ^ A b Mikawa T, Poh AM, Kelly KA, Ishii Y, Reese DE (2004). "Indukce a vzorování primitivní řady, organizační centrum gastrulace v amniotu". Dev Dyn. 229 (3): 422–32. doi:10.1002 / dvdy.10458. PMID 14991697. S2CID 758473.
- ^ A b Downs KM. (2009). „Záhadný primitivní pruh: převládající představy a výzvy týkající se osy těla savců“. BioEssays. 31 (8): 892–902. doi:10.1002 / bies.200900038. PMC 2949267. PMID 19609969.
- ^ Chuai M, Zeng W, Yang X, Boychenko V, Glazier JA, Weijer CJ (2006). "Pohyb buněk během tvorby primitivního pruhu kuřat". Dev. Biol. 296 (1): 137–49. doi:10.1016 / j.ydbio.2006.04.451. PMC 2556955. PMID 16725136.
- ^ Chuai M, Weijer CJ (2008). Mechanismy, které jsou základem tvorby primitivních pruhů v kuřecím embryu. Curr Top Dev Biol. Aktuální témata ve vývojové biologii. 81. str. 135–56. doi:10.1016 / S0070-2153 (07) 81004-0. ISBN 9780123742537. PMID 18023726.
- ^ Grapin-Botton, A .; Constam, D. (2007). "Vývoj mechanismů a molekulární řízení tvorby endodermu". Mechanismy rozvoje. 124 (4): 253–78. doi:10.1016 / j.mod.2007.01.001. PMID 17307341. S2CID 16552755.
- ^ A b C Turner, David A .; Rué, Pau; Mackenzie, Jonathan P .; Davies, Eleanor; Martinez Arias, Alfonso (01.01.2014). „Brachyury spolupracuje se signalizací Wnt / β-katenin na vyvolání chování podobného primitivnímu pruhu při diferenciaci myších embryonálních kmenových buněk“. Biologie BMC. 12: 63. doi:10.1186 / s12915-014-0063-7. ISSN 1741-7007. PMC 4171571. PMID 25115237.
- ^ A b Warmflash, Aryeh; Sorre, Benoit; Etoc, Fred; Siggia, Eric D; Brivanlou, Ali H (2014). „Metoda rekapitulace časných embryonálních prostorových vzorů v lidských embryonálních kmenových buňkách“. Přírodní metody. 11 (8): 847–854. doi:10.1038 / nmeth.3016. PMC 4341966. PMID 24973948.
- ^ A b Etoc, Fred; Metzger, Jakob; Ruzo, Albert; Kirst, Christoph; Yoney, Anna; Ozair, M. Zeeshan; Brivanlou, Ali H .; Siggia, Eric D. (2016). „Rovnováha mezi vylučovanými inhibitory a kontrolou hranového snímání gastruloidní samoorganizace“. Vývojová buňka. 39 (3): 302–315. doi:10.1016 / j.devcel.2016.09.016. PMC 5113147. PMID 27746044.
- ^ A b Brink, Susanne C. van den; Baillie-Johnson, Peter; Balayo, Tina; Hadjantonakis, Anna-Katerina; Nowotschin, Sonja; Turner, David A .; Arias, Alfonso Martinez (15. 11. 2014). "Lámání symetrie, specifikace zárodečné vrstvy a axiální organizace v agregátech myších embryonálních kmenových buněk". Rozvoj. 141 (22): 4231–4242. doi:10.1242 / dev.113001. ISSN 0950-1991. PMC 4302915. PMID 25371360.
- ^ A b C Turner, David Andrew; Glodowski, Cherise R .; Luz, Alonso-Crisostomo; Baillie-Johnson, Peter; Hayward, Penny C .; Collignon, Jérôme; Gustavsen, Carsten; Serup, Palle; Schröter, Christian (2016-05-13). "Interakce mezi nodální a Wnt signalizací vedou k robustnímu rozbití symetrie a axiální organizaci u gastruloidů (embryonálních organoidů)". bioRxiv 10.1101/051722.
- ^ A b C Turner, David; Alonso-Crisostomo, Luz; Girgin, Mehmet; Baillie-Johnson, Peter; Glodowski, Cherise R .; Hayward, Penelope C .; Collignon, Jérôme; Gustavsen, Carsten; Serup, Palle (2017-01-31). „Gastruloidy rozvíjejí tři tělesné osy v nepřítomnosti extraembryonálních tkání a prostorově lokalizované signalizace“. bioRxiv 10.1101/104539.
- ^ Beccari, Leonardo; Moris, Naomi; Girgin, Mehmet; Turner, David A .; Baillie-Johnson, Peter; Cossy, Anne-Catherine; Lutolf, Matthias P .; Duboule, Denis; Arias, Alfonso Martinez (říjen 2018). „Víceosé samoorganizační vlastnosti myších embryonálních kmenových buněk do gastruloidů“. Příroda. 562 (7726): 272–276. Bibcode:2018Natur.562..272B. doi:10.1038 / s41586-018-0578-0. ISSN 0028-0836. PMID 30283134. S2CID 52915553.
- ^ Turner, David A .; Girgin, Mehmet; Alonso-Crisostomo, Luz; Trivedi, Vikas; Baillie-Johnson, Peter; Glodowski, Cherise R .; Hayward, Penelope C .; Collignon, Jérôme; Gustavsen, Carsten (01.11.2017). „Anteroposteriorní polarita a prodloužení v nepřítomnosti mim embryonálních tkání a prostorově lokalizované signalizace v gastruloidech: embryonální organoidy savců“. Rozvoj. 144 (21): 3894–3906. doi:10.1242 / dev.150391. ISSN 0950-1991. PMC 5702072. PMID 28951435.
Bibliografie
- Arnold, Sebastian J .; Robertson, Elizabeth J. (2009). "Přijetí závazku: alokace buněčné linie a vzorování os v časném embryu myši". Nat. Rev. Mol. Cell Biol. 10 (2): 91–103. doi:10.1038 / nrm2618. PMID 19129791. S2CID 94174.

- Catala, Martin (2005). "Embryologie páteře a míchy". V Tortori-Donati, Paolo; et al. (eds.). Dětská neuroradiologie: mozek. Springer. ISBN 978-3-540-41077-5.
- Ereskovsky, Alexander V. (2010). Srovnávací embryologie hub. Springer. ISBN 978-90-481-8574-0.
- Gilbert, Scott F. (2010). Vývojová biologie (Deváté vydání.). Sinauer Associates. ISBN 978-0-87893-558-1.
- Hall, Brian Keith (1998). „8.3.3 Gastrula a gastrulace“. Evoluční vývojová biologie (2. vyd.). Nizozemsko: Kluwer Academic Publishers. ISBN 978-0-412-78580-1.
- Harrison, Lionel G. (2011). Tvarování života: Generování biologického vzorce. Cambridge University Press. ISBN 978-0-521-55350-6.
- McGeady, Thomas A., ed. (2006). „Gastrulation“. Veterinární embryologie. Wiley-Blackwell. ISBN 978-1-4051-1147-8.
- Mundlos, Stefan (2009). „Genová akce: vývojová genetika“. In Speicher, Michael; et al. (eds.). Lidská genetika Vogela a Motulského: Problémy a přístupy (4. vydání). Springer. doi:10.1007/978-3-540-37654-5. ISBN 978-3-540-37653-8.
- Tam, Patrick P.L .; Behringer, Richard R. (1997). „Myší gastrulace: tvorba tělesného plánu savce“. Mech. Dev. 68 (1–2): 3–25. doi:10.1016 / S0925-4773 (97) 00123-8. PMID 9431800. S2CID 14052942.

Další čtení
- Baron, Margaret H. (2001). „Embryonální indukce hematopoézy savců a vaskulogeneze“. V Zon, Leonard I. (ed.). Hematopoéza: vývojový přístup. Oxford University Press. ISBN 978-0-19-512450-7.
- Cullen, K.E. (2009). „embryologie a časný vývoj zvířat“. Encyclopedia of life science, svazek 2. Infobase. ISBN 978-0-8160-7008-4.
- Forgács, G. & Newman, Stuart A. (2005). „Štěpení a tvorba blastul“. Biologická fyzika vyvíjejícího se embrya. Cambridge University Press. Bibcode:2005bpde.book ..... F. ISBN 978-0-521-78337-8.CS1 maint: používá parametr autoři (odkaz)
- Forgács, G. & Newman, Stuart A. (2005). "Epiteliální morfogeneze: gastrulace a neurulace". Biologická fyzika vyvíjejícího se embrya. Cambridge University Press. Bibcode:2005bpde.book ..... F. ISBN 978-0-521-78337-8.CS1 maint: používá parametr autoři (odkaz)
- Hart, Nathan H. & Fluck, Richard A. (1995). "Epiboly a Gastrulation". V Capco, David (ed.). Cytoskeletální mechanismy během vývoje zvířat. Akademický tisk. ISBN 978-0-12-153131-7.CS1 maint: používá parametr autoři (odkaz)
- Knust, Elizabeth (1999). "Gastrulační pohyby". V Birchmeier, Walter; Birchmeier, Carmen (eds.). Epiteliální morfogeneze ve vývoji a nemoci. CRC Press. str. 152–153. ISBN 978-90-5702-419-1.
- Kunz, Yvette W. (2004). "Gastrulation". Vývojová biologie ryb Teleost. Springer. ISBN 978-1-4020-2996-7.
- Nation, James L., ed. (2009). "Gastrulation". Fyziologie a biochemie hmyzu. CRC Press. ISBN 978-0-8493-1181-9.
- Ross, Lawrence M .; Lamperti, Edward D., eds. (2006). „Lidská ontogeneze: Gastrulace, neurulace a tvorba somitů“. Atlas anatomie: obecná anatomie a pohybový aparát. Thieme. ISBN 978-3-13-142081-7.
- Sanes, Dan H. a kol. (2006). "Brzy embryologie metazoanů". Vývoj nervového systému (2. vyd.). Akademický tisk. s. 1–2. ISBN 978-0-12-618621-5.CS1 maint: používá parametr autoři (odkaz)
- Stanger, Ben Z. & Melton, Douglas A. (2004). „Vývoj endodermálních derivátů v plicích, játrech, slinivce břišní a ve střevech“. In Epstein, Charles J .; et al. (eds.). Vrozené chyby vývoje: molekulární podstata klinických poruch morfogeneze. Oxford University Press. ISBN 978-0-19-514502-1.CS1 maint: používá parametr autoři (odkaz)