Nekódující RNA - Non-coding RNA
Tento článek je hlavní část není adekvátně shrnout klíčové body jeho obsahu. Zvažte prosím rozšíření potenciálního zákazníka na poskytnout přístupný přehled všech důležitých aspektů článku. (Ledna 2018) |

A nekódující RNA (ncRNA) je RNA molekula, která není přeloženo do protein. The DNA sekvence, ze které je transkribována funkční nekódující RNA, se často nazývá RNA gen. Bohaté a funkčně důležité typy nekódujících RNA zahrnout přenos RNA (tRNA) a ribozomální RNA (rRNA), stejně jako malé RNA, jako je mikroRNA, siRNA, piRNA, snoRNA, snRNA, exRNA, scaRNA a dlouhé ncRNA jako Xist a HORKÝ VZDUCH.
Počet nekódujících RNA v lidském genomu není znám; nicméně nedávné transkriptomický a bioinformatické studie naznačují, že jich jsou tisíce.[1][2][3][4][5][6] Mnoho z nově identifikovaných ncRNA nebylo ověřeno pro svou funkci.[7] Je také pravděpodobné, že mnoho ncRNA je nefunkčních (někdy označovaných jako zbytečná RNA) a jsou produktem falešného přepisu.[8][9]
Předpokládá se, že nekódující RNA přispívají k onemocněním včetně rakovina a Alzheimerova choroba.
Historie a objevy
Nukleové kyseliny byly poprvé objeveny v roce 1868 Friedrich Miescher[10] a do roku 1939 byla RNA zapletena do proteosyntéza.[11] O dvě desetiletí později Francis Crick předpověděl funkční složku RNA, která zprostředkovala překlad; usoudil, že RNA je vhodnější pro pár bází s transkriptem mRNA než čistá polypeptid.[12]

První nekódující RNA, která byla charakterizována, byla alanin tRNA nalezena v pekařské droždí, jeho struktura byla zveřejněna v roce 1965.[13] Chcete-li vyrobit vyčištěný vzorek alaninové tRNA, Robert W. Holley et al. použitý 140kg komerčních pekárenských kvasnic dát jen 1G purifikované tRNAAla pro analýzu.[14] 80 nukleotid tRNA byla sekvenována nejprve štěpením Pankreatická ribonukleáza (produkující fragmenty končící na Cytosin nebo Uridin ) a poté ribonukleázou takadiastázy Tl (produkující fragmenty, které skončily Guanosin ). Chromatografie a identifikace 5 'a 3' konců pak pomohla uspořádat fragmenty za účelem stanovení sekvence RNA.[14] Ze tří struktur původně navržených pro tuto tRNA,[13] struktura „čtyřlístku“ byla nezávisle navržena v několika následujících publikacích.[15][16][17][18] Čtyřlístek sekundární struktura byl dokončen následovně Rentgenová krystalografie analýza provedená dvěma nezávislými výzkumnými skupinami v roce 1974.[19][20]
Ribozomální RNA byl dále objeven, následovaný URNA na začátku 80. let. Od té doby pokračuje objev nových nekódujících RNA snoRNA, Xist, CRISPR a mnoho dalších.[21] Mezi nedávné významné doplňky patří riboswitche a miRNA; objev RNAi mechanismus spojené s posledně jmenovaným získal Craig C. Mello a Andrew Fire 2006 Nobelova cena za fyziologii nebo medicínu.[22]
Nedávných objevů ncRNA bylo dosaženo prostřednictvím experimentálních i bioinformatické metody.
Biologické role
Nekódující RNA patří do několika skupin a účastní se mnoha buněčných procesů. Ty sahají od ncRNA centrálního významu, které jsou konzervovány po celý nebo většinu buněčného života, až po přechodnější ncRNA specifické pro jeden nebo několik blízce příbuzných druhů. Za konzervativnější ncRNA se považují molekulární fosilie nebo relikvie z poslední univerzální společný předek a Svět RNA a jejich současné role zůstávají většinou v regulaci toku informací z DNA do proteinu.[23][24][25]
V překladu

Mnoho konzervovaných, esenciálních a hojných ncRNA je zapojeno do překlad. Ribonukleoprotein (RNP) nazývané částice ribozomy jsou „továrny“, kde překlad probíhá v buňce. Ribozom se skládá z více než 60% ribozomální RNA; jsou tvořeny 3 ncRNA v prokaryoty a 4 ncRNA v eukaryoty. Ribozomální RNA katalyzují translaci nukleotidových sekvencí na protein. Další sada ncRNA, Přeneste RNA, tvoří „molekulu adaptéru“ mezi mRNA a bílkoviny. The H / ACA box a C / D box snoRNA jsou ncRNA nalezené v archaeách a eukaryotech. RNáza MRP je omezeno na eukaryoty. Obě skupiny ncRNA se podílejí na zrání rRNA. SnoRNA vedou kovalentní modifikace rRNA, tRNA a snRNA; RNáza MRP štěpí vnitřní transkribovaný spacer 1 mezi 18S a 5,8S rRNA. Všudypřítomná ncRNA, RNáza P, je evoluční příbuzný RNázy MRP.[27] RNáza P maturuje sekvence tRNA generováním zralých 5'-konců tRNA štěpením 5'-vedoucích prvků prekurzorových tRNA. Volal další všudypřítomný RNP SRP rozpoznává a transportuje specifické rodící se proteiny do endoplazmatické retikulum v eukaryoty a plazmatická membrána v prokaryoty. V bakteriích Transfer-messenger RNA (tmRNA) je RNP zapojený do záchrany zastavených ribozomů, označování neúplné polypeptidy a podpora degradace aberantní mRNA.[Citace je zapotřebí ]
Při sestřihu RNA
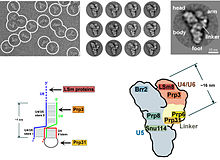
V eukaryotech spliceosome provádí sestřih reakce nezbytné pro odstranění intron sekvencí, je tento proces nutný pro tvorbu zralých mRNA. The spliceosome je další RNP, často také známý jako snRNP nebo tri-snRNP. Existují dvě různé formy spliceosomu, hlavní a vedlejší formy. Složky ncRNA hlavního spliceosomu jsou U1, U2, U4, U5, a U6. Složky ncRNA minoritního spliceosomu jsou U11, U12, U5, U4atac a U6atac.[Citace je zapotřebí ]
Další skupina intronů může katalyzovat své vlastní odstranění z hostitelských přepisů; tito jsou voláni self-sestřih RNA. Existují dvě hlavní skupiny samo-sestřihových RNA: skupina I katalytický intron a katalytický intron skupiny II. Tyto ncRNA katalyzují vlastní excizi z prekurzorů mRNA, tRNA a rRNA v široké škále organismů.[Citace je zapotřebí ]
U savců bylo zjištěno, že snoRNA mohou také regulovat alternativní sestřih mRNA, například snoRNA HBII-52 reguluje sestřih serotoninový receptor 2C.[28]
U hlístic se SmY Zdá se, že ncRNA je zapojena do mRNA trans-sestřih.[Citace je zapotřebí ]
Při replikaci DNA

Y RNA jsou smyčky stonků, nutné pro replikace DNA prostřednictvím interakcí s chromatin a iniciační proteiny (včetně komplex rozpoznávání původu ).[30][31] Jsou také součástí Ribonukleoproteinová částice Ro60[32] který je cílem autoimunitních protilátek u pacientů s systémový lupus erythematodes.[33]
V regulaci genů
The výraz mnoha tisíců geny jsou regulovány ncRNA. K této regulaci může dojít v trans nebo v cis. Existuje stále více důkazů, že se nazývá speciální typ ncRNA enhancerové RNA, transkribované z oblasti zesilovače genu, působí na podporu genové exprese.[Citace je zapotřebí ]
Trans-působící
U vyšších eukaryot mikroRNA regulovat genovou expresi. Jedna miRNA může snížit hladinu exprese stovek genů. Mechanismus, kterým zralé molekuly miRNA působí, je částečnou komplementární k jedné nebo více molekulám messenger RNA (mRNA), obvykle v 3 'UTR. Hlavní funkcí miRNA je snížit regulaci genové exprese.
NcRNA RNáza P Bylo také prokázáno, že ovlivňuje genovou expresi. V lidském jádru RNáza P je vyžadován pro normální a efektivní transkripci různých ncRNA transkribovaných pomocí RNA polymeráza III. Patří mezi ně tRNA, 5S rRNA, SRP RNA a U6 snRNA geny. RNáza P uplatňuje svoji roli v transkripci prostřednictvím asociace s Pol III a chromatin aktivních genů tRNA a 5S rRNA.[34]
Bylo prokázáno, že 7SK RNA, a metazoan ncRNA, působí jako negativní regulátor RNA polymeráza II činitel prodloužení P-TEFb, a že tato aktivita je ovlivněna způsoby reakce na stres.[Citace je zapotřebí ]
Bakteriální ncRNA, 6S RNA, konkrétně se asociuje s holoenzymem RNA polymerázy obsahujícím sigma70 faktor specificity. Tato interakce potlačuje výraz závislý na sigma70 promotér v době stacionární fáze.[Citace je zapotřebí ]
Další bakteriální ncRNA, OxyS RNA potlačuje překlad vazbou na Shine-Dalgarno sekvence, čímž uzavírá vazbu ribozomu. OxyS RNA je indukována v reakci na oxidační stres u Escherichia coli.[Citace je zapotřebí ]
B2 RNA je malý nekódující transkript RNA polymerázy III, který potlačuje transkripci mRNA v reakci na tepelný šok v myších buňkách. B2 RNA inhibuje transkripci vazbou na jádro Pol II. Prostřednictvím této interakce se B2 RNA shromažďuje do preiniciačních komplexů na promotoru a blokuje syntézu RNA.[35]
Nedávná studie ukázala, že právě transkripce sekvence ncRNA může mít vliv na genovou expresi. RNA polymeráza II transkripce ncRNA je nutná pro chromatin přestavba v Schizosaccharomyces pombe. Chromatin se postupně převádí na otevřenou konfiguraci, protože se přepisuje několik druhů ncRNA.[36]
Cis-herectví
V 5 'je vložena řada ncRNA UTR (Nepřeložené regiony) z geny kódující proteiny a ovlivňovat jejich projev různými způsoby. Například a riboswitch může přímo vázat a malá cílová molekula; vazba cíle ovlivňuje aktivitu genu.[Citace je zapotřebí ]
Vůdce RNA sekvence se nacházejí upstream od prvního genu biosyntetických operonů aminokyselin. Tyto RNA prvky tvoří jednu ze dvou možných struktur v oblastech kódujících velmi krátké peptidové sekvence, které jsou bohaté na aminokyselinu konečného produktu operonu. Struktura terminátoru se tvoří, když je nadbytek regulační aminokyseliny a není bráněn pohyb ribozomu přes transkript vedoucího. Když je nedostatek nabité tRNA regulační aminokyseliny, ribosom překládající vedoucí peptid se zastaví a vytvoří se antiterminátorová struktura. To umožňuje RNA polymeráze přepsat operon. Známí vůdci RNA jsou Vedoucí histidinového operonu, Vůdce leucinových operonů, Vedoucí treoninského operonu a Vůdce tryptofanového operonu.[Citace je zapotřebí ]
Železné prvky odezvy (IRE) jsou vázáni proteiny reakce na železo (IRP). IRE se nachází v UTR různých mRNA jejichž výrobky jsou součástí metabolismus železa. Když je koncentrace železa nízká, IRP se vážou na feritin mRNA IRE, což vede k translační represi.[Citace je zapotřebí ]
Interní místa vstupu ribozomu (IRES) jsou Struktury RNA které umožňují překlad iniciace uprostřed sekvence mRNA jako součást procesu proteosyntéza.[Citace je zapotřebí ]
V obraně genomu
RNA reagující na Piwi (piRNA) vyjádřené vsavčí testy a somatické buňky tvoří komplexy RNA-protein s Piwi bílkoviny. Tyto piRNA komplexy (piRC) byly spojeny s umlčováním transkripčních genů retrotranspozony a další genetické prvky v zárodečná linie buňky, zejména ty v spermatogeneze.
Klastrovaná pravidelně rozložená krátká palindromická opakování (CRISPR) jsou opakování nalezená v DNA z mnoha bakterie a archaea. Opakování je odděleno rozpěrkami podobné délky. Bylo prokázáno, že tyto spacery lze odvodit z fága a následně pomáhat chránit buňku před infekcí.
Struktura chromozomu
Telomeráza je RNP enzym který přidává konkrétní DNA sekvence se opakuje („TTAGGG“ u obratlovců) až telomerní oblasti, které se nacházejí na koncích eukaryotických chromozomy. Telomery obsahují kondenzovaný materiál DNA, který poskytuje stabilitu chromozomům. Enzym je a reverzní transkriptáza který nese Telomerázová RNA, který se používá jako šablona, když prodlužuje telomery, které se po každém zkracují replikační cyklus.
Xist (X-inactive-specific transkript) je dlouhý gen ncRNA na X chromozom z placentární savci který působí jako hlavní efektor Inaktivace chromozomu X proces formování Barrova těla. An antisense RNA, Tsix, je negativní regulátor společnosti Xist. X chromozomy postrádající expresi Tsix (a tedy s vysokou úrovní transkripce Xist) jsou inaktivovány častěji než normální chromozomy. v drosophilids, které také používají Systém určování pohlaví XY, roX (RNA na X) RNA se účastní kompenzace dávky.[37] Xist i roX fungují epigenetický regulace transkripce prostřednictvím náboru enzymy modifikující histon.
Bifunkční RNA
Bifunkční RNAnebo dvojfunkční RNA, jsou RNA, které mají dvě odlišné funkce.[38][39] Většina známých bifunkčních RNA jsou mRNA, které kódují jak protein, tak ncRNA. Rostoucí počet ncRNA však spadá do dvou různých kategorií ncRNA; např., H / ACA box snoRNA a miRNA.[40][41]
Dva dobře známé příklady bifunkčních RNA jsou SgrS RNA a RNAIII. Je však známo, že existuje několik dalších bifunkčních RNA (např. Aktivátor steroidních receptorů / SRA,[42] VegT RNA,[43][44] Oskarova RNA,[45] ENOD40,[46] RNA p53[47] a SR1 RNA.[48] Bifunkční RNA byly v poslední době předmětem zvláštního vydání Biochimie.[49]
Jako hormon
Mezi určitými nekódujícími RNA a kontrolou hormonálně regulovaných drah existuje důležité spojení. v Drosophila, hormony jako ekdyson a juvenilní hormon může podporovat expresi určitých miRNA. Kromě toho k této regulaci dochází v různých časových bodech uvnitř C. elegans rozvoj.[50] U savců miR-206 je rozhodujícím regulátorem estrogen -receptor-alfa.[51]
Nekódující RNA jsou klíčové při vývoji několika endokrinních orgánů, stejně jako při endokrinních onemocněních, jako je diabetes mellitus.[52] Konkrétně v buněčné linii MCF-7 bylo přidání 17β-estradiol zvýšená globální transkripce nekódujících RNA nazývaných lncRNA poblíž kódujících genů aktivovaných estrogenem.[53]
V patogenní vyhýbání
C. elegans bylo prokázáno, že se učí a dědí patogenní vyhýbání se po expozici jedné nekódující RNA a bakteriální patogen.[54][55]
Role v nemoci
Stejně jako u bílkoviny, mutace nebo nerovnováha v repertoáru ncRNA v těle mohou způsobit různé nemoci.
Rakovina
Mnoho ncRNA vykazuje abnormální vzorce exprese v rakovinný papírové kapesníky.[5] Tyto zahrnují miRNA, dlouhé mRNA podobné ncRNA,[56][57] GAS5,[58] SNORD50,[59] telomerázová RNA a Y RNA.[60] MiRNA se účastní regulace velkého rozsahu mnoha genů kódujících proteiny,[61][62] Y RNA jsou důležité pro zahájení replikace DNA,[30] telomerázová RNA, která slouží jako primer pro telomerázu, RNP, která se rozšiřuje telomerické oblasti na koncích chromozomů (viz telomery a nemoci Pro více informací). Přímá funkce dlouhých nRNA podobných mRNA je méně jasná.
Zárodečná linie mutace v miR-16-1 a miR-15 Bylo prokázáno, že primární prekurzory jsou mnohem častější u pacientů s chronická lymfocytární leukémie ve srovnání s kontrolní populací.[63][64]
Bylo navrženo, že vzácné SNP (rs11614913 ), který se překrývá has-mir-196a2 bylo zjištěno, že je spojen s nemalobuněčný karcinom plic.[65] Podobně obrazovka 17 miRNA, u nichž se předpokládá, že regulují řadu genů spojených s rakovinou prsu, našla variace v mikroRNA miR-17 a miR-30c-1of pacientů; tito pacienti nebyli nositeli BRCA1 nebo BRCA2 mutace, což dává možnost, že familiární rakovina prsu může být způsobena změnami v těchto miRNA.[66]The p53 supresor nádoru je pravděpodobně nejdůležitějším činidlem v prevenci tvorby a progrese nádoru. Protein p53 funguje jako transkripční faktor s klíčovou rolí při řízení buněčné stresové reakce. Kromě své klíčové role v rakovině se p53 podílí na dalších onemocněních včetně cukrovky, buněčné smrti po ischemii a různých neurodegenerativních onemocnění, jako jsou Huntington, Parkinson a Alzheimer. Studie naznačují, že exprese p53 podléhá regulaci nekódující RNA.[4]
Dalším příkladem nekódující RNA dysregulované v rakovinných buňkách je dlouhá nekódující RNA Linc00707. Linc00707 je upregulovaný a houby miRNA v mezenchymálních kmenových buňkách odvozených z lidské kostní dřeně,[67] u hepatocelulárního karcinomu,[68] rakovina žaludku[69] nebo rakovina prsu,[70][71] a tím podporuje osteogenezi, přispívá k progresi hepatocelulárního karcinomu, podporuje proliferaci a metastázy nebo nepřímo reguluje expresi proteinů podílejících se na agresivitě rakoviny.
Prader-Williho syndrom
Odstranění 48 kopií snoRNA boxu C / D 116 bylo prokázáno, že je primární příčinou Prader-Williho syndrom.[72][73][74][75] Prader-Willi je vývojová porucha spojená s přejídáním a problémy s učením. SNORD116 má potenciální cílová místa v řadě genů kódujících proteiny a mohl by hrát roli v regulaci alternativního sestřihu.[76]
Autismus
Chromozomální lokus obsahující malá nukleolární RNA SNORD115 genový klastr byl duplikován přibližně u 5% jedinců s autistické rysy.[77][78] Model myši navržený tak, aby měl duplikaci klastru SNORD115, zobrazuje chování podobné autismu.[79] Nedávná malá studie posmrtné mozkové tkáně prokázala ve srovnání s kontrolami změněnou expresi dlouhých nekódujících RNA v prefrontální kůře a mozečku autistických mozků.[80]
Hypoplázie chrupavky a vlasů
Mutace uvnitř RNáza MRP bylo prokázáno, že způsobují hypoplázie chrupavky – vlasů, onemocnění spojené s řadou příznaků, jako je nízký vzrůst, řídké vlasy, abnormality skeletu a potlačený imunitní systém, který je častý u Amish a Finština.[81][82][83] Nejlépe charakterizovanou variantou je A-to-G přechod na nukleotidu 70, který je ve smyčkové oblasti dvě báze 5 'a konzervovaný pseudoknot. Mnoho dalších mutací v rámci RNázy MRP však také způsobuje CHH.
Alzheimerova choroba
Antisense RNA, BACE1-AS je přepsán z opačného řetězce do BACE1 a je upregulován u pacientů s Alzheimerova choroba.[84] BACE1-AS reguluje expresi BACE1 zvyšováním stability mACE RNA BACE1 a generováním dalšího BACE1 prostřednictvím post-transkripčního dopředného mechanismu. Stejným mechanismem také zvyšuje koncentrace beta amyloid, hlavní složka senilních plaků. Koncentrace BACE1-AS jsou zvýšené u subjektů s Alzheimerovou chorobou a u transgenních myší s prekurzorem amyloidního proteinu.
miR-96 a ztráta sluchu
Variace v oblasti semen dospělých miR-96 byl spojen s autosomálně dominantní, progresivní ztráta sluchu u lidí a myší. The homozygotní mutantní myši byly hluché hluché a nevykazovaly žádné kochleární odpovědi. Heterozygotní myši a lidé postupně ztrácejí schopnost slyšet.[85][86][87]
Rozdíl mezi funkční RNA (fRNA) a ncRNA
Vědci začali rozlišovat funkční RNA (fRNA) z ncRNA, k popisu oblastí funkčních na úrovni RNA, které mohou nebo nemusí být samostatnými transkripty RNA.[88][89][90] To znamená, že fRNA (jako jsou riboswitche, prvky SECIS a další cis-regulační oblasti) není ncRNA. Přesto by fRNA mohla také zahrnovat mRNA, protože toto je RNA kódující protein, a proto je funkční. dodatečně uměle vyvinuté RNA spadají také pod zastřešující termín fRNA. Některé publikace[21] uveďte to ncRNA a fRNA jsou téměř synonymní, avšak jiní poukázali na to, že velká část anotovaných ncRNA pravděpodobně nemá žádnou funkci.[8][9] Rovněž bylo navrženo jednoduše používat tento výraz RNA, protože rozdíl od proteinu kódujícího RNA (messenger RNA ) je již dán kvalifikátorem mRNA.[91] To eliminuje nejednoznačnost při adresování genu "kódujícího nekódující" RNA. Kromě toho může existovat řada ncRNA, které jsou v publikované literatuře a souborech dat nesprávně označeny.[92][93][94]
Viz také
- Extracelulární RNA
- Seznam RNA
- Struktura nukleové kyseliny
- Rfam
- Riboswitch
- Ribozym
- RNA přítomné ve vzorcích prostředí
- VA (virová asociovaná) RNA
Reference
- ^ Cheng J, Kapranov P, Drenkow J, Dike S, Brubaker S, Patel S a kol. (Květen 2005). "Transkripční mapy 10 lidských chromozomů v rozlišení 5 nukleotidů". Věda. 308 (5725): 1149–54. Bibcode:2005Sci ... 308.1149C. doi:10.1126 / science.1108625. PMID 15790807. S2CID 13047538.
- ^ Konsorcium projektu ENCODE, Birney E, Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, et al. (Červen 2007). „Identifikace a analýza funkčních prvků v 1% lidského genomu pilotním projektem ENCODE“. Příroda. 447 (7146): 799–816. Bibcode:2007Natur.447..799B. doi:10.1038 / nature05874. PMC 2212820. PMID 17571346.
- ^ Washietl S, Pedersen JS, Korbel JO, Stocsits C, Gruber AR, Hackermüller J a kol. (Červen 2007). "Strukturované RNA v ENCODE vybraných oblastech lidského genomu". Výzkum genomu. 17 (6): 852–64. doi:10,1101 / gr. 5650707. PMC 1891344. PMID 17568003.
- ^ A b Morris KV, vyd. (2012). Nekódující RNA a epigenetická regulace genové exprese: ovladače přirozeného výběru. Caister Academic Press. ISBN 978-1-904455-94-3.
- ^ A b Shahrouki P, Larsson E (2012). „Nekódující onkogen: případ chybějících důkazů DNA?“. Frontiers in Genetics. 3: 170. doi:10.3389 / fgene.2012.00170. PMC 3439828. PMID 22988449.
- ^ van Bakel H, Nislow C, Blencowe BJ, Hughes TR (květen 2010). Eddy SR (ed.). „Většina přepisů„ temné hmoty “je spojena se známými geny“. PLOS Biology. 8 (5): e1000371. doi:10.1371 / journal.pbio.1000371. PMC 2872640. PMID 20502517.
- ^ Hüttenhofer A, Schattner P, Polacek N (květen 2005). „Nekódující RNA: naděje nebo humbuk?“. Trendy v genetice. 21 (5): 289–97. doi:10.1016 / j.tig.2005.03.007. PMID 15851066.
- ^ A b Brosius J (květen 2005). "Neplýtvejte, nechci - přepis přebytku u mnohobuněčných eukaryot". Trendy v genetice. 21 (5): 287–8. doi:10.1016 / j.tig.2005.02.014. PMID 15851065.
- ^ A b Palazzo AF, Lee ES (2015). „Nekódující RNA: co je funkční a co je haraburdí?“. Frontiers in Genetics. 6: 2. doi:10.3389 / fgene.2015.00002. PMC 4306305. PMID 25674102.
- ^ Dahm R (únor 2005). „Friedrich Miescher a objev DNA“. Vývojová biologie. 278 (2): 274–88. doi:10.1016 / j.ydbio.2004.11.028. PMID 15680349.
- ^ Caspersson T, Schultz J (1939). "Pentosové nukleotidy v cytoplazmě rostoucích tkání". Příroda. 143 (3623): 602–3. Bibcode:1939Natur.143..602C. doi:10.1038 / 143602c0. S2CID 4140563.
- ^ Crick FH (1958). "Na syntézu bílkovin". Sympózia Společnosti pro experimentální biologii. 12: 138–63. PMID 13580867.
- ^ A b Holley RW a kol. (Březen 1965). "Struktura ribonukleové kyseliny". Věda. 147 (3664): 1462–5. Bibcode:1965Sci ... 147.1462H. doi:10.1126 / science.147.3664.1462. PMID 14263761. S2CID 40989800.
- ^ A b „Nobelova cena za fyziologii nebo medicínu 1968“. Nobelova nadace. Citováno 2007-07-28.
- ^ Madison JT, Everett GA, Kung H (červenec 1966). "Nukleotidová sekvence kvasinkové tyrosinové transferové RNA". Věda. 153 (3735): 531–4. Bibcode:1966Sci ... 153..531M. CiteSeerX 10.1.1.1001.2662. doi:10.1126 / science.153.3735.531. PMID 5938777. S2CID 9265016.
- ^ Zachau HG, Dütting D, Feldmann H, Melchers F, Karau W (1966). "Serinově specifické transferové ribonukleové kyseliny. XIV. Srovnání nukleotidových sekvencí a modelů sekundární struktury". Cold Spring Harbor Symposia o kvantitativní biologii. 31: 417–24. doi:10.1101 / SQB.1966.031.01.054. PMID 5237198.
- ^ Dudock BS, Katz G, Taylor EK, Holley RW (březen 1969). "Primární struktura přenosové RNA fenylalaninu z pšeničných klíčků". Sborník Národní akademie věd Spojených států amerických. 62 (3): 941–5. Bibcode:1969PNAS ... 62..941D. doi:10.1073 / pnas.62.3.941. PMC 223689. PMID 5257014.
- ^ Cramer F, Doepner H, Haar F VD, Schlimme E, Seidel H (prosinec 1968). „O konformaci přenosové RNA“. Sborník Národní akademie věd Spojených států amerických. 61 (4): 1384–91. Bibcode:1968PNAS ... 61.1384C. doi:10.1073 / pnas.61.4.1384. PMC 225267. PMID 4884685.
- ^ Ladner JE a kol. (Listopad 1975). "Struktura kvasinkové fenylalaninové přenosové RNA při rozlišení 2,5 A". Sborník Národní akademie věd Spojených států amerických. 72 (11): 4414–8. Bibcode:1975PNAS ... 72,4414L. doi:10.1073 / pnas.72.11.4414. PMC 388732. PMID 1105583.
- ^ Kim SH a kol. (Leden 1973). "Trojrozměrná struktura kvasinkové fenylalaninové transferové RNA: skládání polynukleotidového řetězce". Věda. 179 (4070): 285–8. Bibcode:1973Sci ... 179..285K. doi:10.1126 / science.179.4070.285. PMID 4566654. S2CID 28916938.
- ^ A b Eddy SR (prosinec 2001). "Nekódující RNA geny a moderní svět RNA". Recenze přírody. Genetika. 2 (12): 919–29. doi:10.1038/35103511. PMID 11733745. S2CID 18347629.
- ^ Daneholt, Bertil. „Advanced Information: RNA interference“. Nobelova cena za fyziologii nebo medicínu 2006. Archivovány od originál dne 2007-01-20. Citováno 2007-01-25.
- ^ Jeffares DC, Poole AM, Penny D (leden 1998). "Pozůstatky ze světa RNA". Journal of Molecular Evolution. 46 (1): 18–36. Bibcode:1998JMolE..46 ... 18J. doi:10.1007 / PL00006280. PMID 9419222. S2CID 2029318.
- ^ Poole AM, Jeffares DC, Penny D (leden 1998). "Cesta ze světa RNA". Journal of Molecular Evolution. 46 (1): 1–17. Bibcode:1998JMolE..46 .... 1P. doi:10.1007 / PL00006275. PMID 9419221. S2CID 17968659.
- ^ Poole A, Jeffares D, Penny D (říjen 1999). „Raná evoluce: prokaryoty, nové děti v bloku“. BioEssays. 21 (10): 880–9. doi:10.1002 / (SICI) 1521-1878 (199910) 21:10 <880 :: AID-BIES11> 3.0.CO; 2-P. PMID 10497339.
- ^ Ban N, Nissen P, Hansen J, Moore PB, Steitz TA (srpen 2000). "Kompletní atomová struktura velké ribozomální podjednotky v rozlišení 2,4 A". Věda. 289 (5481): 905–20. Bibcode:2000Sci ... 289..905B. CiteSeerX 10.1.1.58.2271. doi:10.1126 / science.289.5481.905. PMID 10937989.
- ^ Zhu Y, Stribinskis V, Ramos KS, Li Y (květen 2006). „Sekvenční analýza RNA RNázy MRP odhaluje její původ z eukaryotické RNA RNázy P“. RNA. 12 (5): 699–706. doi:10,1261 / rna.2284906. PMC 1440897. PMID 16540690.
- ^ Kishore S, Stamm S (leden 2006). „SnoRNA HBII-52 reguluje alternativní sestřih serotoninového receptoru 2C“. Věda. 311 (5758): 230–2. Bibcode:2006Sci ... 311..230K. doi:10.1126 / science.1118265. PMID 16357227. S2CID 44527461.
- ^ Stein AJ, Fuchs G, Fu C, Wolin SL, Reinisch KM (květen 2005). „Strukturální pohledy na kontrolu kvality RNA: autoantigen Ro váže špatně poskládané RNA prostřednictvím své centrální dutiny“. Buňka. 121 (4): 529–39. doi:10.1016 / j.cell.2005.03.009. PMC 1769319. PMID 15907467.
- ^ A b Christov CP, Gardiner TJ, Szüts D, Krude T (září 2006). „Funkční požadavek nekódujících Y RNA pro replikaci lidské chromozomální DNA“. Molekulární a buněčná biologie. 26 (18): 6993–7004. doi:10.1128 / MCB.01060-06. PMC 1592862. PMID 16943439.
- ^ Zhang AT, Langley AR, Christov CP, Kheir E, Shafee T, Gardiner TJ, Krude T (červen 2011). „Dynamická interakce Y RNA s chromatinem a iniciačními proteiny během replikace lidské DNA“. Journal of Cell Science. 124 (Pt 12): 2058–69. doi:10.1242 / jcs.086561. PMC 3104036. PMID 21610089.
- ^ Hall AE, Turnbull C, Dalmay T (duben 2013). „Y RNA: nedávný vývoj“. Biomolekulární koncepty. 4 (2): 103–10. doi:10.1515 / bmc-2012-0050. PMID 25436569. S2CID 12575326.
- ^ Lerner MR, Boyle JA, Hardin JA, Steitz JA (leden 1981). "Dvě nové třídy malých ribonukleoproteinů detekovaných protilátkami spojenými s lupus erythematosus". Věda. 211 (4480): 400–2. Bibcode:1981Sci ... 211..400L. doi:10.1126 / science.6164096. PMID 6164096.
- ^ Reiner R, Ben-Asouli Y, Krilovetzky I, Jarrous N (červen 2006). „Role pro katalytický ribonukleoprotein RNázu P v transkripci RNA polymerázy III“. Geny a vývoj. 20 (12): 1621–35. doi:10,1101 / gad.386706. PMC 1482482. PMID 16778078.
- ^ Espinoza CA, Allen TA, Hieb AR, Kugel JF, Goodrich JA (září 2004). „B2 RNA se váže přímo na RNA polymerázu II, aby potlačila syntézu transkriptů“. Přírodní strukturní a molekulární biologie. 11 (9): 822–9. doi:10.1038 / nsmb812. PMID 15300239. S2CID 22199826.
- ^ Hirota K, Miyoshi T, Kugou K, Hoffman CS, Shibata T, Ohta K (listopad 2008). "Postupná remodelace chromatinu kaskádou iniciace transkripce nekódujících RNA". Příroda. 456 (7218): 130–4. Bibcode:2008 Natur.456..130H. doi:10.1038 / nature07348. PMID 18820678. S2CID 4416402.
- ^ Park Y, Kelley RL, Oh H, Kuroda MI, Meller VH (listopad 2002). "Rozsah šíření chromatinu stanoven náborem RNA roX RNA proteinů MSL". Věda. 298 (5598): 1620–3. Bibcode:2002Sci ... 298.1620P. doi:10.1126 / science.1076686. PMID 12446910. S2CID 27167367.
- ^ Wadler CS, Vanderpool CK (prosinec 2007). „Duální funkce pro bakteriální malou RNA: SgrS provádí regulaci závislou na párování bází a kóduje funkční polypeptid“. Sborník Národní akademie věd Spojených států amerických. 104 (51): 20454–9. Bibcode:2007PNAS..10420454W. doi:10.1073 / pnas.0708102104. PMC 2154452. PMID 18042713.
- ^ Dinger ME, Pang KC, Mercer TR, Mattick JS (listopad 2008). McEntyre J (ed.). „Diferenciace proteinové a nekódující RNA: výzvy a nejednoznačnosti“. PLOS výpočetní biologie. 4 (11): e1000176. Bibcode:2008PLSCB ... 4E0176D. doi:10.1371 / journal.pcbi.1000176. PMC 2518207. PMID 19043537.
- ^ Saraiya AA, Wang CC (listopad 2008). Goldberg DE (ed.). „snoRNA, nový předchůdce mikroRNA v Giardia lamblia“. PLOS patogeny. 4 (11): e1000224. doi:10.1371 / journal.ppat.1000224. PMC 2583053. PMID 19043559.
- ^ Ender C, Krek A, Friedländer MR, Beitzinger M, Weinmann L, Chen W, Pfeffer S, Rajewsky N, Meister G (listopad 2008). "Lidská snoRNA s funkcemi podobnými mikroRNA". Molekulární buňka. 32 (4): 519–28. doi:10.1016 / j.molcel.2008.10.017. PMID 19026782.
- ^ Leygue E (srpen 2007). „Aktivátor RNA steroidních receptorů (SRA1): neobvyklé bifacetované genové produkty s podezřením na význam pro rakovinu prsu“. Signalizace jaderného receptoru. 5: e006. doi:10.1621 / nrs.05006. PMC 1948073. PMID 17710122.
- ^ Zhang J, King ML (prosinec 1996). „RNA Xenopus VegT je během oogeneze lokalizována ve vegetativní kůře a kóduje nový transkripční faktor T-boxu, který se podílí na mezodermálním vzorování“. Rozvoj. 122 (12): 4119–29. PMID 9012531.
- ^ Kloc M, Wilk K, Vargas D, Shirato Y, Bilinski S, Etkin LD (srpen 2005). „Potenciální strukturální role nekódujících a kódujících RNA v organizaci cytoskeletu ve vegetativní kůře oocytů Xenopus“. Rozvoj. 132 (15): 3445–57. doi:10,1242 / dev.01919. PMID 16000384.
- ^ Jenny A, Hachet O, Závorszky P, Cyrklaff A, Weston MD, Johnston DS, Erdélyi M, Ephrussi A (srpen 2006). „Role Oskarovy RNA nezávislá na translaci v časné oogenezi Drosophila“. Rozvoj. 133 (15): 2827–33. doi:10.1242 / dev.02456. PMID 16835436.
- ^ Gultyaev AP, Roussis A (2007). „Identifikace konzervovaných sekundárních struktur a expanzních segmentů v enod40 RNA odhaluje nové homology enod40 v rostlinách“. Výzkum nukleových kyselin. 35 (9): 3144–52. doi:10.1093 / nar / gkm173. PMC 1888808. PMID 17452360.
- ^ Candeias MM, Malbert-Colas L, Powell DJ, Daskalogianni C, Maslon MM, Naski N, Bourougaa K, Calvo F, Fåhraeus R (září 2008). Msgstr "P53 mRNA řídí aktivitu p53 řízením funkcí Mdm2". Přírodní buněčná biologie. 10 (9): 1098–105. doi:10.1038 / ncb1770. PMID 19160491. S2CID 5122088.
- ^ Gimpel M, Preis H, Barth E, Gramzow L, Brantl S (prosinec 2012). „SR1 - malá RNA se dvěma pozoruhodně zachovanými funkcemi“. Výzkum nukleových kyselin. 40 (22): 11659–72. doi:10.1093 / nar / gks895. PMC 3526287. PMID 23034808.
- ^ Francastel C, Hubé F (listopad 2011). „Kódování nebo nekódování: Potřebujete, aby byly exkluzivní?“. Biochimie. 93 (11): vi – vii. doi:10.1016 / S0300-9084 (11) 00322-1. PMID 21963143.
- ^ Sempere LF, Sokol NS, Dubrovsky EB, Berger EM, Ambros V (červenec 2003). "Časová regulace exprese mikroRNA v Drosophila melanogaster zprostředkovaná hormonálními signály a široko-komplexní genovou aktivitou". Vývojová biologie. 259 (1): 9–18. doi:10.1016 / S0012-1606 (03) 00208-2. PMID 12812784.
- ^ Adams BD, Furneaux H, White BA (květen 2007). „Mikro-ribonukleová kyselina (miRNA) miR-206 cílí na lidský estrogenový receptor alfa (ERalpha) a potlačuje expresi RNA a proteinu ERalpha v buněčných liniích rakoviny prsu“. Molekulární endokrinologie. 21 (5): 1132–47. doi:10.1210 / me.2007-0022. PMID 17312270.
- ^ Knoll M, Lodish HF, Sun L (březen 2015). „Dlouhé nekódující RNA jako regulátory endokrinního systému“. Recenze přírody. Endokrinologie. 11 (3): 151–60. doi:10.1038 / nrendo.2014.229. hdl:1721.1/116703. PMC 4376378. PMID 25560704.
- ^ Li W, Notani D, Ma Q, Tanasa B, Nunez E, Chen AY, Merkurjev D, Zhang J, Ohgi K, Song X, Oh S, Kim HS, Glass CK, Rosenfeld MG (červen 2013). „Funkční role enhancerových RNA pro aktivaci transkripce závislou na estrogenu“. Příroda. 498 (7455): 516–20. Bibcode:2013Natur.498..516L. doi:10.1038 / příroda12210. PMC 3718886. PMID 23728302.
- ^ „Vědci objevují, jak červi předávají znalosti o patogenu potomkům“. phys.org. Citováno 11. října 2020.
- ^ Kaletsky, Rachel; Moore, Rebecca S .; Vrla, Geoffrey D .; Parsons, Lance R .; Gitai, Zemer; Murphy, Coleen T. (9. září 2020). „C. elegans interpretuje bakteriální nekódující RNA, aby se naučil patogenní vyhýbání“. Příroda: 1–7. doi:10.1038 / s41586-020-2699-5. ISSN 1476-4687. Citováno 11. října 2020.
- ^ Pibouin L, Villaudy J, Ferbus D, Muleris M, Prospéri MT, Remvikos Y, Goubin G (únor 2002). „Klonování mRNA nadměrné exprese v karcinomu tlustého střeva-1: sekvence nadměrně exprimovaná v podskupině karcinomů tlustého střeva“. Genetika a cytogenetika rakoviny. 133 (1): 55–60. doi:10.1016 / S0165-4608 (01) 00634-3. PMID 11890990.
- ^ Fu X, Ravindranath L, Tran N, Petrovics G, Srivastava S (březen 2006). „Regulace apoptózy nekódujícím genem specifickým pro prostatu a souvisejícím s rakovinou prostaty, PCGEM1“. DNA a buněčná biologie. 25 (3): 135–41. doi:10.1089 / dna.2006.25.135. PMID 16569192.
- ^ Mourtada-Maarabouni M, Pickard MR, Hedge VL, Farzaneh F, Williams GT (leden 2009). „GAS5, RNA nekódující proteiny, řídí apoptózu a je downregulován u rakoviny prsu“. Onkogen. 28 (2): 195–208. doi:10.1038 / dne 2008.373. PMID 18836484.
- ^ Dong XY, Guo P, Boyd J, Sun X, Li Q, Zhou W, Dong JT (srpen 2009). "Dopad snoRNA U50 na lidskou rakovinu prsu". Journal of Genetics and Genomics = Yi Chuan Xue Bao. 36 (8): 447–54. doi:10.1016 / S1673-8527 (08) 60134-4. PMC 2854654. PMID 19683667.
- ^ Christov CP, Trivier E, Krude T (březen 2008). „Nekódující lidské Y RNA jsou nadměrně exprimovány v nádorech a jsou nutné pro buněčnou proliferaci“. British Journal of Cancer. 98 (5): 981–8. doi:10.1038 / sj.bjc.6604254. PMC 2266855. PMID 18283318.
- ^ Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, Burge CB, Bartel DP (prosinec 2005). „Široký dopad savčích MicroRNA na represi a evoluci mRNA“. Věda. 310 (5755): 1817–21. Bibcode:2005Sci ... 310.1817F. doi:10.1126 / science.1121158. PMID 16308420. S2CID 1849875.
- ^ Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, Bartel DP, Linsley PS, Johnson JM (únor 2005). „Analýza microarray ukazuje, že některé mikroRNA downregulují velké množství cílových mRNA“. Příroda. 433 (7027): 769–73. Bibcode:2005 Natur.433..769L. doi:10.1038 / nature03315. PMID 15685193. S2CID 4430576.
- ^ Calin GA, Ferracin M, Cimmino A, Di Leva G, Shimizu M, Wojcik SE, Iorio MV, Visone R, Sever NI, Fabbri M, Iuliano R, Palumbo T, Pichiorri F, Roldo C, Garzon R, Sevignani C, Rassenti L, Alder H, Volinia S, Liu CG, Kipps TJ, Negrini M, Croce CM (říjen 2005). "MikroRNA podpis spojený s prognózou a progresí u chronické lymfocytární leukémie". The New England Journal of Medicine. 353 (17): 1793–801. doi:10.1056 / NEJMoa050995. PMID 16251535.
- ^ Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, Rassenti L, Kipps T, Negrini M, Bullrich F, Croce CM (listopad 2002). „Časté delece a down-regulace mikroRNA genů miR15 a miR16 ve 13q14 u chronické lymfocytární leukémie“. Sborník Národní akademie věd Spojených států amerických. 99 (24): 15524–9. Bibcode:2002PNAS ... 9915524C. doi:10.1073 / pnas.242606799. PMC 137750. PMID 12434020.
- ^ Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L, Zeng Y, Miao R, Jin G, Ma H, Chen Y, Shen H (červenec 2008). „Genetické varianty sekvencí miRNA a přežití nemalobuněčného karcinomu plic“. The Journal of Clinical Investigation. 118 (7): 2600–8. doi:10,1172 / JCI34934. PMC 2402113. PMID 18521189.
- ^ Shen J, Ambrosone CB, Zhao H (březen 2009). "Nové genetické varianty v genech microRNA a familiární rakovině prsu". International Journal of Cancer. 124 (5): 1178–82. doi:10.1002 / ijc.24008. PMID 19048628.
- ^ Jia, Bo; Wang, Zhiping; Sun, Xiang; Chen, červen; Zhao, Jianjiang; Qiu, Xiaoling (prosinec 2019). „Dlouhé nekódující RNA LINC00707 houby miR-370-3p na podporu osteogeneze mezenchymálních kmenových buněk odvozených z lidské kostní dřeně prostřednictvím upregulace WNT2B“. Výzkum a terapie kmenových buněk. 10 (1): 67. doi:10.1186 / s13287-019-1161-9. ISSN 1757-6512. PMC 6387535. PMID 30795799.
- ^ Tu, Jianfei; Zhao, Zhongwei; Xu, min; Chen, Minjiang; Weng, Qiaoyou; Wang, Jiangmei; Ji, Jiansong (červenec 2019). „LINC00707 přispívá k progresi hepatocelulárního karcinomu pomocí hubování miR ‐ 206 ke zvýšení CDK14“. Journal of Cellular Physiology. 234 (7): 10615–10624. doi:10,1002 / jcp.27737. ISSN 0021-9541.
- ^ Xie, Min; Ma, Tianshi; Xue, Jiangyang; Ma, Hongwei; Slunce, Ming; Zhang, Zhihong; Liu, Minjuan; Liu, Yinghua; Ju, Songwen; Wang, Zhaoxia; De, Wei (únor 2019). „Dlouhá intergenní neproteinová kódující RNA 707 podporuje proliferaci a metastázování rakoviny žaludku interakcí s proteinem HuR stabilizujícím mRNA“. Dopisy o rakovině. 443: 67–79. doi:10.1016 / j.canlet.2018.11.032.
- ^ Li, Tong; Li, Yunpeng; Sun, Hongyan (06.06.2019). „MicroRNA-876 je hubená dlouhou nekódující RNA LINC00707 a přímo cílí na metadherin, aby inhibovala malignitu rakoviny prsu“. Management a výzkum rakoviny. doi:10,2147 / cmar.s210845. PMC 6559252. PMID 31239777. Citováno 2020-09-27.
- ^ Yuan, R.-X .; Bao, D .; Zhang, Y. (květen 2020). „Linc00707 podporuje proliferaci, invazi a migraci buněk prostřednictvím regulační smyčky miR-30c / CTHRC1 u rakoviny prsu“. Evropská recenze pro lékařské a farmakologické vědy. 24 (9): 4863–4872. doi:10,26355 / eurrev_202005_21175. ISSN 1128-3602.
- ^ Sahoo T, del Gaudio D, německý JR, Shinawi M, Peters SU, osoba RE, Garnica A, Cheung SW, Beaudet AL (červen 2008). „Prader-Williho fenotyp způsobený otcovským nedostatkem pro HBII-85 C / D box, malá nukleolární shluk RNA“. Genetika přírody. 40 (6): 719–21. doi:10,1038 / ng.158. PMC 2705197. PMID 18500341.
- ^ Skryabin BV, Gubar LV, Seeger B, Pfeiffer J, Handel S, Robeck T, Karpova E, Rozhdestvensky TS, Brosius J (prosinec 2007). „Odstranění klastru genu MBII-85 snoRNA u myší má za následek retardaci růstu po narození“. Genetika PLOS. 3 (12): e235. doi:10.1371 / journal.pgen.0030235. PMC 2323313. PMID 18166085.
- ^ Ding F, Li HH, Zhang S, Solomon NM, Camper SA, Cohen P, Francke U (březen 2008). Akbarian S (ed.). „SleRNA Snord116 (Pwcr1 / MBII-85) delece způsobuje růstový deficit a hyperfágii u myší“. PLOS ONE. 3 (3): e1709. Bibcode:2008PLoSO ... 3.1709D. doi:10.1371 / journal.pone.0001709. PMC 2248623. PMID 18320030.
- ^ Ding F, Prints Y, Dhar MS, Johnson DK, Garnacho-Montero C, Nicholls RD, Francke U (červen 2005). „Nedostatek snoRNA Pwcr1 / MBII-85 je kritický pro neonatální letalitu u myších modelů Prader-Williho syndromu“. Savčí genom. 16 (6): 424–31. doi:10.1007 / s00335-005-2460-2. PMID 16075369. S2CID 12256515.
- ^ Bazeley PS, Shepelev V, Talebizadeh Z, Butler MG, Fedorova L, Filatov V, Fedorov A (leden 2008). „snoTARGET ukazuje, že lidské osiřelé cíle snoRNA se nacházejí poblíž alternativních spojovacích spojů“. Gen. 408 (1–2): 172–9. doi:10.1016 / j.gene.2007.10.037. PMC 6800007. PMID 18160232.
- ^ Bolton PF, Veltman MW, Weisblatt E, Holmes JR, Thomas NS, Youings SA, Thompson RJ, Roberts SE, Dennis NR, Browne CE, Goodson S, Moore V, Brown J (září 2004). „Abnormality chromozomu 15q11-13 a další zdravotní stavy u jedinců s poruchami autistického spektra“. Psychiatrická genetika. 14 (3): 131–7. doi:10.1097/00041444-200409000-00002. PMID 15318025. S2CID 37344935.
- ^ Cook EH, Scherer SW (říjen 2008). "Varianty počtu kopií spojené s neuropsychiatrickými stavy". Příroda. 455 (7215): 919–23. Bibcode:2008Natur.455..919C. doi:10.1038 / nature07458. PMID 18923514. S2CID 4377899.
- ^ Nakatani J, Tamada K, Hatanaka F, Ise S, Ohta H, Inoue K, Tomonaga S, Watanabe Y, Chung YJ, Banerjee R, Iwamoto K, Kato T, Okazawa M, Yamauchi K, Tanda K, Takao K, Miyakawa T , Bradley A, Takumi T (červen 2009). "Abnormální chování v modelu myši s chromozomovým inženýrstvím pro lidskou duplikaci 15q11-13 pozorováno u autismu". Buňka. 137 (7): 1235–46. doi:10.1016 / j.cell.2009.04.024. PMC 3710970. PMID 19563756.
- ^ Ziats MN, Rennert OM (březen 2013). „Aberantní exprese dlouhých nekódujících RNA v autistickém mozku“. Journal of Molecular Neuroscience. 49 (3): 589–93. doi:10.1007 / s12031-012-9880-8. PMC 3566384. PMID 22949041.
- ^ Ridanpää M, van Eenennaam H, Pelin K, Chadwick R, Johnson C, Yuan B, van Venrooij W, Pruijn G, Salmela R, Rockas S, Mäkitie O, Kaitila I, de la Chapelle A (leden 2001). „Mutace v RNA složce RNázy MRP způsobují pleiotropní lidské onemocnění, hypoplázii chrupavky a vlasů“. Buňka. 104 (2): 195–203. doi:10.1016 / S0092-8674 (01) 00205-7. PMID 11207361. S2CID 13977736.
- ^ Martin AN, Li Y (březen 2007). „RNáza MRP RNA a lidské genetické nemoci“. Cell Research. 17 (3): 219–26. doi:10.1038 / sj.cr.7310120. PMID 17189938.
- ^ Kavadas FD, Giliani S, Gu Y, Mazzolari E, Bates A, Pegoiani E, Roifman CM, Notarangelo LD (prosinec 2008). "Variabilita klinických a laboratorních znaků u pacientů s ribonukleázovou mitochondriální RNA zpracovávající genové mutace endoribonukleázy". The Journal of Allergy and Clinical Immunology. 122 (6): 1178–84. doi:10.1016 / j.jaci.2008.07.036. PMID 18804272.
- ^ Faghihi MA, Modarresi F, Khalil AM, Wood DE, Sahagan BG, Morgan TE, Finch CE, St Laurent G, Kenny PJ, Wahlestedt C (červenec 2008). „Exprese nekódující RNA je u Alzheimerovy choroby zvýšena a řídí rychlou dopřednou regulaci beta-sekretázy“. Přírodní medicína. 14 (7): 723–30. doi:10,1038 / nm1784. PMC 2826895. PMID 18587408.
- ^ Mencía A, Modamio-Høybjør S, Redshaw N, Morín M, Mayo-Merino F, Olavarrieta L, Aguirre LA, del Castillo I, Steel KP, Dalmay T, Moreno F, Moreno-Pelayo MA (květen 2009). „Mutace v oblasti semen lidského miR-96 jsou zodpovědné za nesyndromickou progresivní ztrátu sluchu“. Genetika přírody. 41 (5): 609–13. doi:10,1038 / ng.355. PMID 19363479. S2CID 11113852.
- ^ Lewis MA, Quint E, Glazier AM, Fuchs H, De Angelis MH, Langford C, van Dongen S, Abreu-Goodger C, Piipari M, Redshaw N, Dalmay T, Moreno-Pelayo MA, Enright AJ, Steel KP (květen 2009 ). „Mutace miR-96 vyvolaná ENU spojená s progresivní ztrátou sluchu u myší“. Genetika přírody. 41 (5): 614–8. doi:10,1038 / ng.369. PMC 2705913. PMID 19363478.
- ^ Soukup GA (červen 2009). „Malý, ale hlasitý: malé RNA mají výrazný vliv na vývoj uší“. Výzkum mozku. 1277: 104–14. doi:10.1016 / j.brainres.2009.02.027. PMC 2700218. PMID 19245798.
- ^ Carter RJ, Dubchak I, Holbrook SR (říjen 2001). „Výpočtový přístup k identifikaci genů pro funkční RNA v genomových sekvencích“. Výzkum nukleových kyselin. 29 (19): 3928–38. doi:10.1093/nar/29.19.3928. PMC 60242. PMID 11574674.
- ^ Pedersen JS, Bejerano G, Siepel A, Rosenbloom K, Lindblad-Toh K, Lander ES, Kent J, Miller W, Haussler D (April 2006). "Identification and classification of conserved RNA secondary structures in the human genome". PLOS výpočetní biologie. 2 (4): e33. Bibcode:2006PLSCB...2...33P. doi:10.1371/journal.pcbi.0020033. PMC 1440920. PMID 16628248.
- ^ Thomas JM, Horspool D, Brown G, Tcherepanov V, Upton C (January 2007). "GraphDNA: a Java program for graphical display of DNA composition analyses". BMC bioinformatika. 8: 21. doi:10.1186/1471-2105-8-21. PMC 1783863. PMID 17244370.
- ^ Brosius J, Raabe CA (February 2015). "What is an RNA? A top layer for RNA classification". RNA Biology. 13 (2): 140–4. doi:10.1080/15476286.2015.1128064. PMC 4829331. PMID 26818079.
- ^ Ji, Zhe; Song, Ruisheng; Regev, Aviv; Struhl, Kevin (2015-12-19). "Many lncRNAs, 5'UTRs, and pseudogenes are translated and some are likely to express functional proteins". eLife. 4: e08890. doi:10.7554/eLife.08890. ISSN 2050-084X. PMC 4739776. PMID 26687005.
- ^ Tosar, Juan Pablo; Rovira, Carlos; Cayota, Alfonso (2018-01-22). "Non-coding RNA fragments account for the majority of annotated piRNAs expressed in somatic non-gonadal tissues". Komunikační biologie. 1 (1): 2. doi:10.1038/s42003-017-0001-7. ISSN 2399-3642. PMC 6052916. PMID 30271890.
- ^ Housman, Gali; Ulitsky, Igor (January 2016). "Methods for distinguishing between protein-coding and long noncoding RNAs and the elusive biological purpose of translation of long noncoding RNAs". Biochimica et Biophysica Acta (BBA) - regulační mechanismy genů. 1859 (1): 31–40. doi:10.1016/j.bbagrm.2015.07.017. ISSN 0006-3002. PMID 26265145.
externí odkazy
(Wayback Machine kopírovat)
- The Rfam Database — a curated list of hundreds of families of related ncRNAs
- NONCODE.org — a free database of all kinds of noncoding RNAs (except tRNAs and rRNAs)
- RNAcon Prediction and classification of ncRNA BMC Genomics 2014, 15:127
- ENCODE threads explorer Non-coding RNA characterization. Příroda (deník)
- The Non-coding RNA Databases Resource (NRDR) — a curated source of data related to over non-coding RNA databases available over the internet
- DASHR - a database of small non-coding RNAs Bioinformatics 2018
Typy nukleové kyseliny | |||||||
|---|---|---|---|---|---|---|---|
| Složky | |||||||
| Ribonukleové kyseliny (kódování, nekódování ) |
| ||||||
| Deoxyribonukleové kyseliny | |||||||
| Analogy | |||||||
| Klonovací vektory | |||||||
| |||||||