Serinová proteáza - Serine protease - Wikipedia
| Serinové endopeptidázy | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Krystalová struktura hovězího chymotrypsinu. Katalytické zbytky jsou zobrazeny jako žluté tyčinky. Vykresleno z PDB 1CBW. | |||||||||
| Identifikátory | |||||||||
| EC číslo | 3.4.21.- | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Serinové proteázy (nebo serinové endopeptidázy) jsou enzymy to štěpení peptidové vazby v bílkoviny, ve kterém serin slouží jako nukleofilní aminokyselina na (enzym) Aktivní stránky.[1] Vyskytují se všudypřítomně v obou eukaryoty a prokaryoty. Serinové proteázy spadají do dvou širokých kategorií na základě jejich struktury: chymotrypsin -jako (podobné trypsinu) nebo subtilisin -jako.[2]
Klasifikace
The MEROPS systém klasifikace proteáz se počítá 16 superrodiny (od roku 2013), z nichž každý obsahuje mnoho rodiny. Každá nadčeleď používá katalytická triáda nebo dyad v jiném proteinový záhyb a tak představují konvergentní evoluce z katalytický mechanismus. Většina patří do rodiny S1 Klan PA (nadčeleď) proteáz.
Pro superrodiny, P = nadčeleď, obsahující směs nukleofil třídní rodiny, S = čistě serinové proteázy. nadčeleď. V každé nadrodině rodiny jsou označeny jejich katalytickým nukleofilem (S = serinové proteázy).
Rodiny serinových proteáz
| Nadčeleď | Rodiny | Příklady |
|---|---|---|
| SB | S8, S53 | Subtilisin (Bacillus licheniformis ) |
| SC | S9, S10, S15, S28, S33, S37 | Prolyl oligopeptidáza (Sus scrofa ) |
| SE | S11, S12, S13 | D-Ala-D-Ala peptidáza C (Escherichia coli ) |
| SF | S24, S26 | Signální peptidáza Já (Escherichia coli ) |
| SH | S21, S73, S77, S78, S80 | Cytomegalovirus assemblin (člověk herpesvirus 5) |
| SJ | S16, S50, S69 | Lon-A peptidáza (Escherichia coli ) |
| SK | S14, S41, S49 | Clp proteáza (Escherichia coli ) |
| TAK | S74 | Fágový K1F endosialidázový CIMCD samoštěpící protein (Enterobacteria fág K1F ) |
| SP | S59 | Nucleoporin 145 (Homo sapiens ) |
| SR | S60 | Laktoferin (Homo sapiens ) |
| SS | S66 | Murein tetrapeptidáza LD-karboxypeptidáza (Pseudomonas aeruginosa ) |
| SVATÝ | S54 | Kosodélník -1 (Drosophila melanogaster ) |
| PA | S1, S3, S6, S7, S29, S30, S31, S32, S39, S46, S55, S64, S65, S75 | Chymotrypsin A (Bos taurus ) |
| PB | S45, S63 | Penicilin G acyláza předchůdce (Escherichia coli ) |
| PC | S51 | Dipeptidáza E (Escherichia coli ) |
| PE | P1 | DmpA aminopeptidáza (Ochrobactrum anthropi ) |
| Nepřiřazeno | S48, S62, S68, S71, S72, S79, S81 |
Specifičnost podkladu
Serinové proteázy se vyznačují výraznou strukturou, sestávající ze dvou domén beta-barelu, které se sbíhají v katalytickém aktivním místě. Tyto enzymy mohou být dále kategorizovány na základě jejich substrátové specificity jako trypsin-like, chymotrypsin-like nebo elastase-like.[3]
Trypsinový
Trypsinové proteázy štěpí peptidové vazby po pozitivně nabité aminokyselině (lysin nebo arginin ).[4]Tato specificita je způsobena zbytkem, který leží na základně kapsy enzymu S1 (obecně záporně nabitý) kyselina asparagová nebo kyselina glutamová ).
Chymotrypsin
Kapsa S1 enzymů podobných chymotrypsinu je hydrofobnější než v proteázách podobných trypsinu. To má za následek specificitu pro středně velké až velké hydrofobní zbytky, jako je např tyrosin, fenylalanin a tryptofan.
Trombinový
Tyto zahrnují trombin, tkáň aktivující plazminogen a plasmin. Bylo zjištěno, že mají roli při koagulaci a trávení a také v patofyziologii neurodegenerativních poruch, jako je Alzheimerova a Parkinsonova demence.
Jako elastáza
Proteázy podobné elastáze mají mnohem menší rozštěp S1 než proteázy podobné trypsinu nebo chymotrypsinu. V důsledku toho zbytky jako alanin, glycin a valin bývají upřednostňovány.
Subtilisin podobný
Subtilisin je serinová proteáza v prokaryoty. Subtilisin evolučně nesouvisí s chymotrypsinovým klanem, ale sdílí stejný katalytický mechanismus využívající katalytická triáda, k vytvoření nukleofilní látky serin. Toto je klasický příklad používaný k ilustraci konvergentní evoluce, protože stejný mechanismus se během roku vyvinul dvakrát nezávisle vývoj.
Katalytický mechanismus
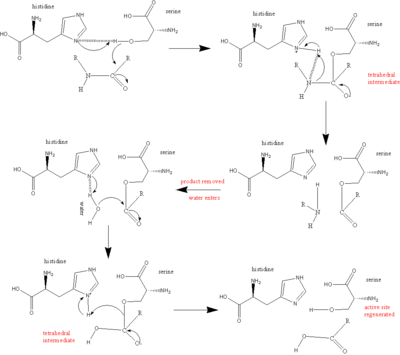
Hlavním hráčem v katalytickém mechanismu serinových proteáz je katalytická triáda. Triáda se nachází v aktivním místě enzymu, kde dochází ke katalýze, a je ve všech zachována superrodiny enzymů serinové proteázy. Triáda je koordinovaná struktura skládající se ze tří aminokyseliny: Jeho 57, Ser 195 (odtud název "serinová proteáza") a Asp 102. Každá z těchto tří klíčových aminokyselin hraje zásadní roli ve schopnosti štěpení proteáz. I když jsou aminokyselinové členy triády umístěny daleko od sebe na sekvenci proteinu, v důsledku skládání budou velmi blízko sebe v srdci enzymu. Zvláštní geometrie členů triády je vysoce charakteristická pro jejich specifickou funkci: bylo prokázáno, že poloha pouhých čtyř bodů triády charakterizuje funkci obsahujícího enzymu.[5]
V případě katalýzy nastane uspořádaný mechanismus, ve kterém je generováno několik meziproduktů. Katalýzu peptidového štěpení lze považovat za a ping-pong katalýza, při které a Podklad váže se (v tomto případě štěpený polypeptid), uvolňuje se produkt („polovina“ peptidu N-konce), váže se další substrát (v tomto případě voda) a uvolňuje se další produkt (C-konec "polovina" peptidu).
Každá aminokyselina v triádě plní v tomto procesu konkrétní úkol:
- The serin má -OH skupinu, která je schopna působit jako nukleofil, útočící na karbonyl uhlík z scissile peptidová vazba substrátu.
- Pár elektronů na histidin dusík má schopnost přijímat vodík z serin -OH skupina, a tak koordinovat útok peptidová vazba.
- The karboxyl skupina na kyselina asparagová na oplátku Vodíkové vazby s histidin, čímž se výše uvedený atom dusíku stává mnohem více elektronegativní.
Celá reakce může být shrnuta následovně:
- The polypeptid substrát se váže na povrch enzymu serinproteázy tak, že scissilní vazba je vložena do aktivního místa enzymu, přičemž karbonylový uhlík této vazby je umístěn v blízkosti nukleofilní serin.
- The serin -OH útočí na karbonyl uhlík a dusík z histidin přijímá vodík z -OH [serinu] a pár elektronů z dvojné vazby karbonyl kyslík se pohybuje na kyslík. Ve výsledku je generován čtyřboký meziprodukt.
- Vazba spojující dusík a uhlík v peptidové vazbě je nyní přerušena. Kovalentní elektrony vytvářející tuto vazbu se pohybují a útočí na vodík histidin, přerušení spojení. Elektrony, které se dříve pohybovaly od karbonyl kyslíková dvojná vazba se pohybuje zpět z negativního kyslíku, aby znovu vytvořila vazbu a vytvořila meziprodukt acyl-enzym.
- Nyní přichází do reakce voda. Voda nahrazuje N-konec štěpeného peptidu a zaútočí na karbonyl uhlík. Opět se elektrony z dvojné vazby pohybují ke kyslíku, což je negativní, protože se vytváří vazba mezi kyslíkem ve vodě a uhlíkem. To je koordinováno dusíkem histidin, který přijímá proton z vody. Celkově to generuje další čtyřboký meziprodukt.
- V závěrečné reakci se vazba vytvořila v prvním kroku mezi serin a karbonyl uhlík se pohybuje k útoku na vodík, který histidin právě získal. Nyní nedostatek elektronů karbonyl uhlík znovu vytváří dvojnou vazbu s kyslíkem. V důsledku toho C-konec peptidu je nyní vysunuto.
Další stabilizační účinky
Bylo zjištěno, že další aminokyseliny proteázy, Gly 193 a Ser 195, se podílejí na tvorbě tzv oxyanion otvor. Oba Gly 193 a Ser 195 může darovat páteřní vodíky pro vodíkové vazby čtyřboký meziprodukt z kroku 1 a kroku 3 jsou generovány záporné ionty kyslíku, které přijaly elektrony z karbonyl dvojná vazba, dokonale zapadá do díry oxyaniontu. Ve skutečnosti se serinové proteázy přednostně vážou na přechodový stav a celková struktura je upřednostňována, což snižuje aktivační energii reakce. Tato „preferenční vazba“ je zodpovědná za velkou část katalytické účinnosti enzymu.
Regulace aktivity serinproteázy
Hostitelské organismy musí zajistit, aby byla aktivita serinových proteáz adekvátně regulována. Toho je dosaženo požadavkem na počáteční aktivaci proteázy a sekreci inhibitorů.
Aktivace zymogenu
Zymogeny jsou obvykle neaktivní prekurzory enzymu. Pokud by trávicí enzymy byly při syntéze aktivní, okamžitě by začaly žvýkat syntetizující orgány a tkáně. Akutní pankreatitida je takový stav, při kterém dochází k předčasné aktivaci trávicích enzymů ve slinivce břišní, což má za následek vlastní trávení (autolýzu). Také to komplikuje posmrtná vyšetřování, protože slinivka často tráví sama, než ji lze vyhodnotit vizuálně.
Zymogeny jsou velké neaktivní struktury, které mají schopnost rozpadat se nebo přecházet na menší aktivované enzymy. Rozdíl mezi zymogeny a aktivovanými enzymy spočívá ve skutečnosti, že aktivní místo pro katalýzu zymogenů je narušeno. Ve výsledku se polypeptid substrátu nemůže účinně vázat a proteolýza nedochází. Teprve po aktivaci, během níž se změní konformace a struktura zymogenu a aktivní místo se otevře proteolýza nastat.
| Zymogen | Enzym | Poznámky |
| Trypsinogen | trypsin | Když trypsinogen vstoupí do tenké střevo ze slinivky břišní, enteropeptidáza sekrece z duodenální sliznice štěpí lysin 15 - isoleucin 16 peptidovou vazbu zymogenu. Výsledkem je, že se zymogen trypsinogen štěpí na trypsin. Připomeňme, že trypsin je také zodpovědný za štěpení lysin peptidové vazby, a tak, jakmile je generováno malé množství trypsinu, podílí se na štěpení vlastního zymogenu a vytváří ještě více trypsinu. Lze tedy nazvat proces aktivace trypsinu autokatalytický. |
| Chymotrypsinogen | chymotrypsin | Poté, co je vazba Arg 15 - Ile 16 v chymotrypsinogenovém zymogenu štěpena trypsinem, nově generovaná struktura zvaná pi-chymotrypsin podstoupí autolýza (vlastní trávení), čímž se získá aktivní chymotrypsin. |
| Proelastáza | elastáza | Aktivuje se štěpením trypsinem. |
Jak je vidět, aktivace trypsinogenu na trypsin je zásadní, protože aktivuje svou vlastní reakci, stejně jako reakci obou chymotrypsin a elastáza. Proto je nezbytné, aby k této aktivaci nedocházelo předčasně. Organismus přijme několik ochranných opatření, která zabrání trávení:
- Aktivace trypsinogenu trypsinem je relativně pomalá
- Zymogeny jsou uloženy v granulích zymogenu, kapslích, které mají stěny, o nichž se předpokládá, že jsou rezistentní vůči proteolýze.
Inhibice
Jsou jisté inhibitory které připomínají čtyřboký meziprodukt, a tak zaplňují aktivní místo a zabraňují správnému fungování enzymu. Trypsin, silný trávicí enzym, se vytváří ve slinivce břišní. Inhibitory zabraňují vlastnímu trávení samotné slinivky břišní.
Serinové proteázy jsou spárovány se serinovou proteázou inhibitory, které vypnou jejich aktivitu, když už nejsou potřeba.[6]
Serinové proteázy jsou inhibovány různou skupinou inhibitory, včetně syntetických chemických inhibitorů pro výzkumné nebo terapeutické účely, a také přírodních proteinových inhibitorů. Jedna rodina přírodních inhibitorů zvaných "serpiny" (zkráceně z inhibitory serinové proteázy ) může tvořit a kovalentní vazba na serinproteázu, inhibice její funkce. Nejlépe studovaný serpiny jsou antitrombin a alfa 1-antitrypsin, studovali svou roli v koagulace /trombóza a emfyzém /A1AT Mezi umělé ireverzibilní malé molekulové inhibitory patří AEBSF a PMSF.
Rodina členovec inhibitory serinové peptidázy, tzv pacifastin, byl identifikován v kobylky a rak a může fungovat v členovci imunitní systém.[7]
Role v nemoci
Mutace mohou vést ke snížené nebo zvýšené aktivitě enzymů. To může mít různé důsledky v závislosti na normální funkci serinové proteázy. Například mutace v protein C. může vést k nedostatek proteinu C. a předisponující k trombóza. Některé proteázy také hrají zásadní roli při aktivaci fúze virus hostitelská buňka-virus aktivací proteinu Spike viru, aby ukázal protein s názvem „fúzní protein“ (TMPRSS2 aktivovat SARS-CoV-2 fúze).
Diagnostické použití
Stanovení hladin serinové proteázy může být užitečné v souvislosti s konkrétními chorobami.
- Koagulační faktor hladiny mohou být vyžadovány při diagnostice hemoragických nebo trombotických stavů.
- Fekální elastáza se používá ke stanovení exokrinní aktivity slinivky břišní, např cystická fibróza nebo chronická pankreatitida.
- Sérum prostatický specifický antigen se používá v screening rakoviny prostaty, stratifikace rizika a monitorování po léčbě.
- Serinová proteáza, jak ji uvolňuje žírné buňky, je důležitým diagnostickým markerem pro přecitlivělost typu 1 reakce (např. anafylaxe ). Užitečnější než např. histamin kvůli delšímu poločas rozpadu, což znamená, že v systému zůstane po klinicky užitečnou dobu.
Viz také
- Serinová hydroláza
- Proteáza
- Klan PA
- Konvergentní evoluce
- Proteolýza
- Katalytická triáda
- Mapa proteolýzy
- Proteázy v angiogenezi
- Intramembránové proteázy
- Inhibitor proteázy (farmakologie)
- Inhibitor proteázy (biologie)
- TopFIND - databáze specificity proteázy, substrátů, produktů a inhibitorů
- MEROPS - Databáze evolučních skupin proteáz
Reference
- ^ Hedstrom, L. (prosinec 2002). "Mechanismus a specificita serinové proteázy". Chem Rev. 102 (12): 4501–24. doi:10.1021 / cr000033x. PMID 12475199.
- ^ Madala PK, Tyndall JD, Nall T, Fairlie DP (červen 2010). "Aktualizace 1 z: Proteasy obecně rozpoznávají řetězce beta na svých aktivních stránkách". Chem Rev. 110 (6): PR1–31. doi:10.1021 / cr900368a. PMID 20377171.
- ^ Ovaere P, Lippens S, Vandenabeele P, Declercq W (srpen 2009). "Vznikající role serinproteázy kaskádovitě v epidermis". Trends Biochem Sci. 34 (9): 453–63. doi:10.1016 / j.tibs.2009.08.001. PMID 19726197.
- ^ Evnin, Luke B .; Vásquez, John R .; Craik, Charles S. (1990). „Substrátová specificita trypsinu vyšetřována pomocí genetické selekce“. Sborník Národní akademie věd Spojených států amerických. 87 (17): 6659–63. doi:10.1073 / pnas.87.17.6659. JSTOR 2355359. PMC 54596. PMID 2204062.
- ^ Iván, Gábor .; Szabadka, Zoltán; Ordög, Rafael; Grolmusz, Vince; Náray-Szabó, Gábor (2009). „Čtyři prostorové body, které definují rodiny enzymů“. Sdělení o biochemickém a biofyzikálním výzkumu. 383 (4): 417–420. CiteSeerX 10.1.1.150.1086. doi:10.1016 / j.bbrc.2009.04.022. PMID 19364497.
- ^ Kimball's Biology Pages, Serine Protease[self-publikoval zdroj? ]
- ^ Breugelmans B, Simonet G, van Hoef V, van Soest S, Vanden BJ (2009). „Peptidy související s pacifastinem: strukturální a funkční charakteristiky rodiny inhibitorů serinové peptidázy“. Peptidy. 30 (3): 622–32. doi:10.1016 / j.peptides.2008.07.026. PMID 18775459.
externí odkazy
- The MEROPS online databáze peptidáz a jejich inhibitorů: Serinová peptidáza
- Serinové proteázy místo na Univerzita v Saint Louis (SLU)
- Serin + proteázy v americké národní lékařské knihovně Lékařské předměty (Pletivo)