METAP2 - METAP2






Methioninaminopeptidáza 2 je enzym že u lidí je kódován METAP2 gen.[5][6]
Methioninaminopeptidáza 2, člen rodiny dimetallohydrolázy, je a cytosolický metaloenzym, který katalyzuje hydrolytické odstranění N-konce methionin zbytky z rodícího se bílkoviny.[7][8][9]
- peptid-methionin peptid + methionin

MetAP2 se vyskytuje ve všech organismech a je obzvláště důležitý kvůli své kritické roli při opravě tkáně a degradaci proteinů.[7] Kromě toho je obzvláště zajímavý MetAP2, protože enzym hraje klíčovou roli angiogeneze, růst nových krevních cév, který je nezbytný pro progresi onemocnění včetně solidního nádoru rakoviny a revmatoidní artritida.[10] MetAP2 je také cílem dvou skupin antiangiogenních přírodních produktů, ovalicin a fumagillin a jejich analogy.[11][12][13][14]
Struktura
V živých organismech začátek kodon který iniciuje bílkoviny syntéza kódy buď methioninu (eukaryoty ) nebo formylmethionin (prokaryoty ). v E-coli (prokaryot), an enzym volala formylmethionin deformyláza může štěpit formylovou skupinu a ponechat pouze N-koncový methioninový zbytek. U proteinů s malými, nenabitými předposledními N-koncovými zbytky může methionin aminopeptidáza štěpit methioninový zbytek.[7]Počet geny kódování methionin aminopeptidázy se mezi organismy liší. V E. coli existuje pouze jeden známý MetAP, 29 333 Da monomerní enzym kódovaný genem skládajícím se z 264 kodony.[7] Vyřazení tohoto genu v E. coli vede k buněčné neviditelnosti.[15] U lidí existují dva geny kódující MetAP, MetAP1 a MetAP2. MetAP1 kóduje enzym 42 kDa, zatímco MetAP2 kóduje 67 kDa enzym. Droždí MetAP1 je ze 40 procent homologní s E. coli MetAP; v rámci S. cerevisiae, MetAP2 je 22 procent homologní se sekvencí MetAP1; MetAP2 je mezi nimi vysoce konzervovaný S. cerevisiae a lidé.[16] Na rozdíl od prokaryot jsou eukaryotické kmeny S. cerevisiae, kterým chybí gen buď pro MetAP1 nebo MetAP2, životaschopné, ale vykazují pomalejší růst než kontrolní kmen exprimující oba geny.
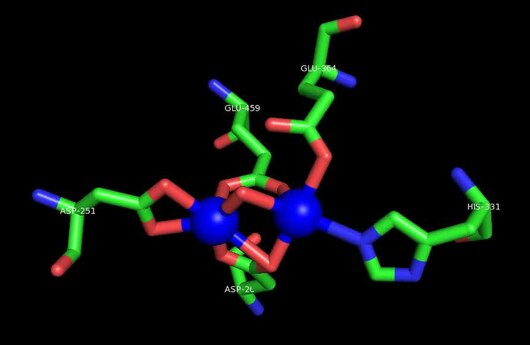
Aktivní stránky
Aktivní místo MetAP2 má strukturální motiv charakteristický pro mnoho metaloenzymů - včetně proteinu nosiče dioxygenu, hemerythrin; dvoujaderný nehemový železný protein, ribonukleotid reduktáza; leucinaminopeptidáza; ureáza; argináza; několik fosfatázy a fosfoesterázy —To zahrnuje dvě přemostění karboxylát ligandy a přemostění voda nebo hydroxid ligand.[7][8][17][18][19][20][21] Konkrétně v lidském MetAP2 (PDB: 1BOA) je jeden z iontů katalytického kovu vázán na His331, Glu364, Glu459, Asp263 a můstkovou vodu nebo hydroxid, zatímco druhý kovový iont je vázán na Asp251 (bidentate ), App262 (bidentate), Glu459 a stejná přemosťující voda nebo hydroxid. Zde jsou dvěma přemosťujícími karboxyláty Asp262 a Glu459.
Dimenzionální centrum
Totožnost Aktivní stránky kov ionty za fyziologických podmínek nebyla úspěšně stanovena a zůstává kontroverzním problémem. MetAP2 ukazuje aktivitu v přítomnosti iontů Zn (II), Co (II), Mn (II) a Fe (II) a různí autoři tvrdí, že jakýkoli daný kovový ion je fyziologický: některé v přítomnosti žehlička,[22] ostatní v kobalt,[23][24] ostatní v mangan,[25] a ještě další v přítomnosti zinek.[26] Většina krystalografů to však má vykrystalizoval MetAP2 buď v přítomnosti Zn (II) nebo Co (II) (viz databáze PDB).
Mechanismus

Přemosťovací voda nebo hydroxidový ligand působí během hydrolýzní reakce jako nukleofil, ale přesný mechanismus katalýzy dosud není znám.[10][19][28] Katalytické mechanismy hydrolázových enzymů velmi závisí na identitě můstkového ligandu,[29] což může být obtížné určit kvůli obtížnosti studia atomů vodíku pomocí rentgenová krystalografie.
Histidinové zbytky ukázané v mechanismu vpravo, H178 a H79, jsou konzervovány ve všech MetAPs (MetAP1s a MetAP2s) sekvenovaných doposud, což naznačuje, že jejich přítomnost je důležitá pro katalytickou aktivitu.[30] Na základě rentgenových krystalografických dat byl navržen histidin 79 (H79), který pomáhá umístit methioninový zbytek v aktivním místě a přenášet proton na nově exponovaný N-koncový amin.[12] Lowther a kolegové navrhli dva možné mechanismy pro MetAP2 v E. coli, zobrazené vpravo.[14]
Funkce
Zatímco předchozí studie ukázaly, že MetAP2 katalyzuje odstranění N-koncových zbytků methioninu in vitro, funkce tohoto enzymu in vivo může být složitější. Například existuje významná korelace mezi inhibicí enzymatické aktivity MetAP2 a inhibicí buněčného růstu, což implikuje enzym v endoteliální buňka proliferace.[13] Z tohoto důvodu vědci označili MetAP2 za potenciální cíl inhibice angiogeneze. Studie navíc prokázaly, že MetAP2 kopurifikuje a interaguje s podjednotkou α eukaryotického iniciačního faktoru 2 (eIF2 ), protein, který je nezbytný pro syntézu proteinů in vivo.[31] Konkrétně MetAP2 chrání eIF-2α před inhibicí fosforylace z enzymu eIF-2α kináza, inhibuje fosforylaci eIF-2 R-podjednotky katalyzovanou na RNA závislou protein kinázou (PKR) a také reverzuje PKR zprostředkovanou inhibici syntézy proteinů v intaktních buňkách.
Klinický význam

Četné studie implikují MetAP2 v angiogenezi.[13][20][32][33][34] Konkrétně kovalentní vazba ovalicinu nebo fumagilinu epoxid Ukázalo se, že zbytek na aktivním histidickém zbytku MetAP2 inaktivuje enzym, čímž inhibuje angiogenezi. Způsob, jakým MetAP2 reguluje angiogenezi, je však ještě třeba stanovit, takže je zapotřebí další studie k ověření, že antiangiogenní aktivita vyplývá přímo z inhibice MetAP2. Nicméně, jak s růstem, tak s metastáza pevných nádorů silně závislých na angiogenezi, fumagilinu a jeho analogech - včetně TNP-470, kaplostatin, a beloranib - stejně jako ovalicin představují potenciální protirakovinné látky.[33][34]Navíc schopnost MetAP2 snižovat životaschopnost buněk v prokaryotických a malých eukaryotických organismech z něj učinila cíl antibakteriálních látek.[13] Dosud bylo prokázáno, že fumagillin i TNP-470 mají antimalarickou aktivitu jak in vitro, tak in vivo, a fumarranol, další analog fumagilinu, představuje slibný náskok.[34]
Inhibitor METAP2 beloranib (ZGN-433) prokázal účinnost při snižování hmotnosti u těžce obézních jedinců.[35] Inhibitory MetAP2 fungují tak, že obnovují rovnováhu mezi způsoby, jak tělo metabolizuje tuky, což vede k podstatnému úbytku tělesné hmotnosti. Vývoj beloranibu byl zastaven v roce 2016 po úmrtí během klinických studií.[36]
Interakce
Metap2 bylo prokázáno komunikovat s Protein kináza R..[37]
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000111142 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000036112 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Arfin SM, Kendall RL, Hall L, Weaver LH, Stewart AE, Matthews BW, Bradshaw RA (září 1995). „Eukaryotické methionylaminopeptidázy: dvě třídy enzymů závislých na kobaltu“. Proc Natl Acad Sci U S A. 92 (17): 7714–8. Bibcode:1995PNAS ... 92.7714A. doi:10.1073 / pnas.92.17.7714. PMC 41216. PMID 7644482.
- ^ Li X, Chang YH (listopad 1996). „Důkaz, že lidským homologem proteinu asociovaného s krysím iniciačním faktorem 2 (p67) je methionin aminopeptidáza.“ Biochem Biophys Res Commun. 227 (1): 152–9. doi:10.1006 / bbrc.1996.1482. PMID 8858118.
- ^ A b C d E Bennett B, Holz RC (1997). „Studie EPR na mono- a dikobaltitem (II) substituovaných formách aminopeptidázy z Aeromonas proteolytica. Pohled do katalytického mechanismu dinukleárních hydroláz“. J. Am. Chem. Soc. 119 (8): 1923–1933. doi:10.1021 / ja963021v.
- ^ A b Johansson FB, Bond AD, Nielsen UG, Moubaraki B, Murray KS, Berry KJ, Larrabee JA, McKenzie CJ (červen 2008). „Dicobalt II-II, II-III a III-III komplexy jako spektroskopické modely pro aktivní místa dikobaltového enzymu“. Inorg Chem. 47 (12): 5079–92. doi:10.1021 / ic7020534. PMID 18494467.
- ^ Larrabee JA, Leung CH, Moore RL, Thamrong-nawasawat T, Wessler BS (říjen 2004). „Magnetický cirkulární dichroismus a studie rovnováhy vazby kobaltu (II) methionylaminopeptidázy Escherichia coli“. J. Am. Chem. Soc. 126 (39): 12316–24. doi:10.1021 / ja0485006. PMID 15453765.
- ^ A b Folkman J (leden 1995). "Angiogeneze u rakoviny, cévních, revmatických a jiných nemocí". Nat. Med. 1 (1): 27–31. doi:10,1038 / nm0195-27. PMID 7584949. S2CID 5924813.
- ^ Taunton J (červenec 1997). "Jak vyhladovět nádor". Chem. Biol. 4 (7): 493–6. doi:10.1016 / S1074-5521 (97) 90320-3. PMID 9263636.
- ^ A b Sin N, Meng L, Wang MQ, Wen JJ, Bornmann WG, Crews CM (červen 1997). „Antiangiogenní látka fumagilin kovalentně váže a inhibuje methioninaminopeptidázu, MetAP-2“. Proc. Natl. Acad. Sci. USA. 94 (12): 6099–103. Bibcode:1997PNAS ... 94.6099S. doi:10.1073 / pnas.94.12.6099. PMC 21008. PMID 9177176.
- ^ A b C d Griffith EC, Su Z, Turk BE, Chen S, Chang YH, Wu Z, Biemann K, Liu JO (červen 1997). „Methioninaminopeptidáza (typ 2) je běžným cílem inhibitorů angiogeneze AGM-1470 a ovalicinu.“. Chem. Biol. 4 (6): 461–71. doi:10.1016 / S1074-5521 (97) 90198-8. PMID 9224570.
- ^ A b Lowther WT, McMillen DA, Orville AM, Matthews BW (říjen 1998). „Antiangiogenní látka fumagillin kovalentně modifikuje konzervovaný aktivní bod histidinu v methioninaminopeptidáze z Escherichia coli“. Proc. Natl. Acad. Sci. USA. 95 (21): 12153–7. Bibcode:1998PNAS ... 9512153L. doi:10.1073 / pnas.95.21.12153. PMC 22800. PMID 9770455.
- ^ Chang SY, McGary EC, Chang S (červenec 1989). „Gen methioninaminopeptidázy z Escherichia coli je nezbytný pro růst buněk“. J. Bacteriol. 171 (7): 4071–2. doi:10.1128 / jb.171.7.4071-4072.1989. PMC 210164. PMID 2544569.
- ^ Li X, Chang YH (prosinec 1995). „Zpracování aminoterminálního proteinu v Saccharomyces cerevisiae je základní funkcí, která vyžaduje dvě odlišné methioninaminopeptidázy“. Proc. Natl. Acad. Sci. USA. 92 (26): 12357–61. Bibcode:1995PNAS ... 9212357L. doi:10.1073 / pnas.92.26.12357. PMC 40356. PMID 8618900.
- ^ Mizoguchi TJ, Kuzelka J, Spingler B, DuBois JL, Davydov RM, Hedman B, Hodgson KO, Lippard SJ (srpen 2001). „Syntéza a spektroskopické studie nehemových diironových (III) druhů s terminálním hydroperoxidovým ligandem: modely pro hemerythrin“. Inorg Chem. 40 (18): 4662–73. doi:10.1021 / ic010076b. PMID 11511213.
- ^ Hagen KS, Lachicotte R, Kitaygorodskiy A (1993). "Supramolekulární kontrola postupné a selektivní substituce karboxylátového ligandu v komplexech dimetyl (II) s akvakarboxylátem". J. Am. Chem. Soc. 115 (26): 12617–12618. doi:10.1021 / ja00079a064.
- ^ A b Brown DA, Errington W, Glass WK, Haase W, Kemp TJ, Nimir H, Ostrovsky SM, Werner R (listopad 2001). "Magnetické, spektroskopické a strukturní studie dikobaltových hydroxamátů a modelových hydroláz". Inorg Chem. 40 (23): 5962–71. doi:10.1021 / ic0103345. PMID 11681912.
- ^ A b Larrabee JA, Chyun SA, Volwiler AS (listopad 2008). „Studie magnetického cirkulárního dichroismu komplexu dikobalt (II) methionin aminopeptidáza / fumagillin a modelové komplexy dikobalt II-II a II-III“. Inorg Chem. 47 (22): 10499–508. doi:10.1021 / ic8011553. PMID 18921993.
- ^ Wilcox DE (listopad 1996). "Binuclear Metallohydrolases". Chem. Rev. 96 (7): 2435–2458. doi:10.1021 / cr950043b. PMID 11848832.
- ^ D'souza VM, Holz RC (srpen 1999). „Methionylaminopeptidáza z Escherichia coli může fungovat jako enzym železa (II)“. Biochemie. 38 (34): 11079–85. doi:10.1021 / bi990872h. PMID 10460163.
- ^ Chang YH, Teichert U, Smith JA (duben 1992). "Molekulární klonování, sekvenování, delece a nadměrná exprese genu methioninaminopeptidázy z Saccharomyces cerevisiae". J. Biol. Chem. 267 (12): 8007–11. PMID 1569059.
- ^ Ghosh M, Grunden AM, Dunn DM, Weiss R, Adams MW (září 1998). „Charakterizace nativních a rekombinantních forem neobvyklé prolin dipeptidázy (prolidázy) závislé na kobaltu z hypertermofilního archaeonu Pyrococcus furiosus“. J. Bacteriol. 180 (18): 4781–9. doi:10.1128 / JB.180.18.4781-4789.1998. PMC 107500. PMID 9733678.
- ^ Wang J, Sheppard GS, Lou P, Kawai M, Park C, Egan DA, Schneider A, Bouska J, Lesniewski R, Henkin J (květen 2003). „Fyziologicky relevantním kovovým kofaktorem pro methioninaminopeptidázu-2 je mangan“. Biochemie. 42 (17): 5035–42. doi:10.1021 / bi020670c. PMID 12718546.
- ^ Leopoldini M, Russo N, Toscano M (červen 2007). „Který z Zn (II), Co (II), Mn (II) a Fe (II) je nejúčinnějším iontem pro reakci katalyzovanou methionin aminopeptidázou?“. J. Am. Chem. Soc. 129 (25): 7776–84. doi:10.1021 / ja068168t. PMID 17523636.
- ^ Lowther WT, Zhang Y, Sampson PB, Honek JF, Matthews BW (listopad 1999). "Pohledy na mechanismus Escherichia coli methionin aminopeptidázy ze strukturní analýzy reakčních produktů a analogů přechodného stavu na bázi fosforu". Biochemie. 38 (45): 14810–9. doi:10.1021 / bi991711g. PMID 10555963.
- ^ Schultz BE, Ye B, Li X, Chan SI (1997). "Electronic Paramagnetic Resonance and Magnetic Properties of Model Complexes for Binuclear Active Sites in Hydrolase Enzymes". Inorg. Chem. 36 (12): 2617–2622. doi:10.1021 / ic960988r.
- ^ Korendovych IV, Kryatov SV, Reiff WM, Rybak-Akimova EV (listopad 2005). „Diiron (II) mu-aqua-mu-hydroxo model pro nehemová místa železa v proteinech“. Inorg Chem. 44 (24): 8656–8. doi:10.1021 / ic051739i. PMID 16296818.
- ^ Li JY, Cui YM, Chen LL, Gu M, Li J, Nan FJ, Ye QZ (květen 2004). „Mutace v místech S1 methioninaminopeptidáz z Escherichia coli a Homo sapiens odhalují zbytky kritické pro substrátovou specificitu“. J. Biol. Chem. 279 (20): 21128–34. doi:10,1074 / jbc.M401679200. PMID 14976199.
- ^ Wu S, Rehemtulla A, Gupta NK, Kaufman RJ (červen 1996). „S glykoproteinem spojeným s eukaryotickým iniciačním faktorem 2 67 kDa glykoprotein částečně obrací inhibici syntézy proteinů aktivovanou dvouvláknovou RNA-dependentní proteinovou kinázou v intaktních buňkách“. Biochemie. 35 (25): 8275–80. doi:10.1021 / bi953028 +. PMID 8679583.
- ^ Benny O, Fainaru O, Adini A, Cassiola F, Bazinet L, Adini I, Pravda E, Nahmias Y, Koirala S, Corfas G, D'Amato RJ, Folkman J (červenec 2008). „Perorálně podávaná formulace s malou molekulou s antiangiogenní a protirakovinnou aktivitou“. Nat. Biotechnol. 26 (7): 799–807. doi:10.1038 / nbt1415. PMC 2803109. PMID 18587385.
- ^ A b Sato Y (2004). „Aminopeptidázy ve zdraví a nemocech: Role aminopeptidázy v angiogenezi“. Biol. Pharm. Býk. 27 (6): 772–776. doi:10,1248 / bpb.27,772. PMID 15187415.
- ^ A b C Chen X, Xie S, Bhat S, Kumar N, Shapiro TA, Liu JO (únor 2009). „Fumagillin a fumarranol interagují s methioninaminopeptidázou 2 P. falciparum a inhibují růst parazitů malárie in vitro a in vivo“. Chem. Biol. 16 (2): 193–202. doi:10.1016 / j.chembiol.2009.01.006. PMID 19246010.
- ^ „Zafgen oznamuje pozitivní data topline fáze 1b pro ZGN-433 při obezitě“. MedNews. Drugs.com. 01.01.2011. Citováno 2011-04-13.
- ^ „Zafgen zastavuje vývoj Beloranibu, aby snížil počet pracovních míst o ~ 34%“. nasdaq.com. 20. července 2016.
- ^ Gil J, Esteban M, Roth D (prosinec 2000). „Regulace in vivo dsRNA-dependentní protein kinázy PKR buněčným glykoproteinem p67“. Biochemie. 39 (51): 16016–25. doi:10.1021 / bi001754t. PMID 11123929.
Další čtení
- Prigmore E, Ahmed S, Best A, Kozma R, Manser E, Segal AW, Lim L (květen 1995). „A 68-kDa kináza a NADPH oxidázová složka p67phox jsou cíle pro Cdc42Hs a Rac1 v neutrofilech“. J. Biol. Chem. 270 (18): 10717–22. doi:10.1074 / jbc.270.18.10717. PMID 7738010.
- Li X, Chang YH (únor 1995). "Molekulární klonování lidské komplementární DNA kódující protein spojený s iniciačním faktorem 2 (p67)". Biochim. Biophys. Acta. 1260 (3): 333–6. doi:10.1016 / 0167-4781 (94) 00227-t. PMID 7873610.
- Ray MK, Chakraborty A, Datta B, Chattopadhyay A, Saha D, Bose A, Kinzy TG, Wu S, Hileman RE, Merrick WC (květen 1993). "Charakteristiky eukaryotického iniciačního faktoru 2 asociovaného 67-kDa polypeptidu". Biochemie. 32 (19): 5151–9. doi:10.1021 / bi00070a026. PMID 8098621.
- Liu S, Widom J, Kemp CW, Crews CM, Clardy J (listopad 1998). "Struktura lidské methionin aminopeptidázy-2 v komplexu s fumagillinem". Věda. 282 (5392): 1324–7. Bibcode:1998Sci ... 282.1324L. doi:10.1126 / science.282.5392.1324. PMID 9812898.
- Griffith EC, Su Z, Niwayama S, Ramsay CA, Chang YH, Liu JO (prosinec 1998). „Molekulární rozpoznávání inhibitorů angiogeneze fumagilinu a ovalicinu methioninaminopeptidázou 2“. Proc. Natl. Acad. Sci. USA. 95 (26): 15183–8. Bibcode:1998PNAS ... 9515183G. doi:10.1073 / pnas.95.26.15183. PMC 28017. PMID 9860943.
- Datta B, Datta R, Mukherjee S, Zhang Z (1999). „Zvýšená fosforylace eukaryotického iniciačního faktoru 2alfa na hranici G2 / M v lidských buňkách osteosarkomu koreluje s deglykosylací p67 a sníženou rychlostí syntézy proteinů“. Exp. Cell Res. 250 (1): 223–30. doi:10.1006 / excr.1999.4508. PMID 10388536.
- Gil J, Esteban M, Roth D (2001). „In vivo regulace dsRNA-dependentní protein kinázy PKR buněčným glykoproteinem p67“. Biochemie. 39 (51): 16016–25. doi:10.1021 / bi001754t. PMID 11123929.
- Catalano A, Romano M, Robuffo I, Strizzi L, Procopio A (srpen 2001). „Methioninaminopeptidáza-2 reguluje přežití lidských mezoteliomových buněk: role exprese Bcl-2 a aktivity telomerázy“. Dopoledne. J. Pathol. 159 (2): 721–31. doi:10.1016 / S0002-9440 (10) 61743-9. PMC 1850571. PMID 11485930.
- Endo H, Takenaga K, Kanno T a kol. (2002). „Methioninaminopeptidáza 2 je novým cílem proteinu spojeného s metastázami, S100A4“. J. Biol. Chem. 277 (29): 26396–402. doi:10,1074 / jbc.M202244200. PMID 11994292.
- Kanno T, Endo H, Takeuchi K a kol. (2002). „Vysoká exprese methioninaminopeptidázy typu 2 v B buňkách zárodečných center a jejich neoplastických protějšcích“. Laboratoř. Investovat. 82 (7): 893–901. doi:10.1097 / 01.LAB.0000020419.25365.C4. PMID 12118091.
- Datta R, Tammali R, Datta B (2003). „Negativní regulace ochrany aktivity fosforylace eIF2alfa jedinečnou kyselou doménou přítomnou na N-konci p67.“ Exp. Cell Res. 283 (2): 237–46. doi:10.1016 / S0014-4827 (02) 00042-3. PMID 12581743.
- Serero A, Giglione C, Sardini A, Martinez-Sanz J, Meinnel T (prosinec 2003). „V lidské mitochondriální N-koncové dráze methioninové excize se objevují neobvyklé peptidové deformylázy“. J. Biol. Chem. 278 (52): 52953–63. doi:10,1074 / jbc.M309770200. PMID 14532271.
- Selvakumar P, Lakshmikuttyamma A, Kanthan R, Kanthan SC, Dimmock JR, Sharma RK (duben 2004). „Vysoká exprese methioninaminopeptidázy 2 v lidských kolorektálních adenokarcinomech“. Clin. Cancer Res. 10 (8): 2771–5. doi:10.1158 / 1078-0432.CCR-03-0218. PMID 15102683.
- Kim S, LaMontagne K, Sabio M, Sharma S, Versace RW, Yusuff N, Phillips PE (květen 2004). „Vyčerpání methioninaminopeptidázy 2 nemění buněčnou odpověď na fumagillin nebo bengamidy.“. Cancer Res. 64 (9): 2984–7. doi:10.1158 / 0008-5472.CAN-04-0019. PMID 15126329.
Galerie PDB | |
|---|---|
|