Orientia tsutsugamushi - Orientia tsutsugamushi - Wikipedia
| Orientia tsutsugamushi | |
|---|---|
 | |
| Orientia tsutsugamushi | |
| Vědecká klasifikace | |
| Doména: | |
| Kmen: | |
| Třída: | |
| Objednat: | |
| Rodina: | |
| Rod: | |
| Druh: | O. tsutsugamushi |
| Binomické jméno | |
| Orientia tsutsugamushi (Hayashi, 1920) (Ogata, 1929) Tamura et al., 1995 | |
Orientia tsutsugamushi (z japonský tsutsuga což znamená "nemoc" a mushi což znamená „hmyz“) je roztoč bakterie patřící do rodiny Rickettsiaceae a je zodpovědný za onemocnění zvané drhnout tyfus u lidí.[1] Je to přirozené a povinná intracelulární parazit roztočů patřících do rodiny Trombiculidae.[2][3] S genomem pouze 2,0–2,7 Mb jej má nejvíce opakované sekvence DNA mezi dosud sekvenovanými bakteriálními genomy. Nemoc, drhnout tyfus, nastává, když infikované larvy roztočů náhodně kousnou lidi. Primárně označeno nediferencovaným febrilní nemoci, infekce může být komplikovaná a často smrtelná.
Orientia tsutsugamushi infekce byla poprvé hlášena v Japonsku Hakuju Hashimotem v roce 1810 a v západním světě Theobaldem Adrianem Palmem v roce 1878. Naosuke Hayashi ji poprvé popsal v roce 1920 a pojmenoval ji Theileria tsutsugamushi. Díky svým jedinečným vlastnostem byl přejmenován Orientia tsutsugamushi v roce 1995. Na rozdíl od jiných Gramnegativní bakterie, není snadno obarvený Gramovo barvení, protože její buněčná stěna postrádá lipofosfoglykan a peptidoglykan. S vysoce variabilním membránovým proteinem, 56-kDa proteinem, lze bakterii antigenně klasifikovat do mnoha kmenů (podtypů). Klasickými kmeny jsou Karp (což představuje asi 50% všech infekcí), Gilliam (25%), Kato (méně než 10%), Shimokoshi, Kuroki a Kawasaki.[4] U každého kmene dále existuje obrovská variabilita.
Orientia tsutsugamushi se přirozeně udržuje v populaci roztočů přenosem ze samice do vajíček (transovariální přenos) a z vajíček na larvy a poté na dospělé (tranštadiální přenos). Volaly larvy roztočů chiggers, jsou přirozené ektoparazity hlodavců. Lidé se nakazí při náhodném kontaktu s infikovanými chiggery. Volala se jizva podobná chrastě eschar je dobrým indikátorem infekce, ale není všudypřítomný. Bakterie je endemická pro takzvaný Tsutsugamushi Triangle, region pokrývající ruský Dálný východ na severu, Japonsko na východě, severní Austrálii na jihu a Afghánistán na západě. Odhaduje se, že se každoročně vyskytne jeden milion infekcí. Antibiotika jako např azithromycin a doxycyklin jsou hlavními léky na předpis; chloramfenikol a tetracyklin jsou také účinné. Diagnóza infekce je obtížná a vyžaduje pracné techniky, jako je Weil – Felixův test, rychlý imunochromatografické test, imunofluorescenční testy, a polymerázová řetězová reakce. Proti infekci neexistuje žádná vakcína.
Dějiny
Nejstarší záznam o O. tsutsugamushi infekce byla ve 3. století (313 n. l.) v Číně.[5] Japonci také po staletí znali souvislost mezi infekcí a roztoči. Dali několik jmen jako shima-mushi, akamushi (červený roztoč) nebo kedani (chlupatý roztoč) onemocnění severního Japonska a nejpopulárněji jako tsutsugamushi (z tsutsuga což znamená horečku, újmu nebo nemoc a mushi což znamená brouk nebo hmyz). Japonský lékař Hakuju Hashimoto poskytl první lékařskou zprávu od Prefektura Niigata v roce 1810. Zaznamenal prevalenci infekce podél břehů horních přítoků Řeka Shinano.[6] První zprávu pro západní svět podal lékař Theobald Adrian Palm Edinburgh Medical Missionary Society v Niigata v roce 1878. Popisující své zkušenosti z první ruky Palm napsal:
Loni v létě [tj. 1877], měl jsem příležitost pozorovat nemoc, která, pokud vím, je pro Japonsko typická a dosud nebyla popsána. Vyskytuje se navíc v určitých dobře označených okresech a v určitém ročním období, takže příležitosti k jeho vyšetřování se často nevyskytují. Je zde známý jako shima-mushinebo ostrovní nemoc hmyzu a je pojmenována podle víry, že je způsobena kousnutím nebo bodnutím nějakého hmyzu zvláštního pro určité ostrovy v řece známé jako Shinagawa, která se vlévá do moře v Niigata.[7]
Etiologie onemocnění byla neznámá až do počátku 20. století. V roce 1908 předpokládali Taichi Kitashima a Mikinosuke Miyajima teorii roztočů o přenosu nemoci tsutsugamushi.[8] V roce 1915 navrhl britský zoolog Stanley Hirst, že larvy roztočů Microtrombidium akamushi (později přejmenováno Leptotrombidium akamushi), který našel na uších polních myší, mohl přenášet a přenášet infekci.[9] V roce 1917 Mataro Nagayo a jeho kolegové podali první úplný popis vývojových stádií, jako je vajíčko, nymfa, larva a dospělý roztoč. Rovněž tvrdili, že pouze larvy kousají savce, a jsou tedy jedinými nositeli parazitů.[10] Ale potom nebyl znám skutečný infekční agens a byl obecně přičítán buď viru, nebo prvokům.[11]
Příčinný patogen poprvé identifikoval Naosuke Hayashi v roce 1920. S přesvědčením, že organismus byl prvokem, uzavřel Hayashi a uvedl: „Dospěl jsem k závěru, že virus choroby je druhem Piroplasma dotyčný [prvoky] ... považuji organismus v Tsutsugamushiho chorobě za dosud nepopsaný druh a na návrh Dr. Henry B. Warda jej označím jako Theileria tsutsugamushi."[12] Objevování podobností s bakterií R. prowazekii, Mataro Nagayo a kolegové dali novou klasifikaci se jménem Rickettsia orientalis v roce 1930.[13][14] (R. prowazekii je příčinná bakterie epidemického tyfu, poprvé objevená americkými lékaři Howard Taylor Ricketts a Russell M. Wilder v roce 1910 a popsal je brazilský lékař Henrique da Rocha Lima v roce 1916.[15])
Taxonomický zmatek se zhoršil. V roce 1931 dal jméno Norio Ogata Rickettsia tsutsugamushi,[16] zatímco Rinya Kawamüra a Yoso Imagawa nezávisle představili jméno Rickettsia akamushi.[17] Kawamüra a Imagawa zjistili, že bakterie jsou uloženy ve slinných žlázách roztočů a že roztoči se živí tělními (lymfatickými) tekutinami, čímž potvrzují skutečnost, že roztoči přenášejí parazity během krmení.[18]
Více než 60 let neexistovala shoda ohledně výběru jména - obojí R. orientalis a R. tsutsugamushi byly stejně použity. Akira Tamura a kolegové v roce 1991 uvedli strukturální rozdíly bakterie od Rickettsia druh, který vyžadoval samostatný rod, a navrhl jméno Orientia tsutsugamushi.[19] Nakonec v roce 1995 vytvořili novou klasifikaci založenou na morfologických a biochemických vlastnostech, formálně vytvořili nový název O. tsutsugamushi.[20]
Biologie

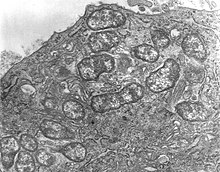
Orientia tsutsugamushi je gramnegativní bakterie a je trvalým (povinným) parazitem u roztočů. V rámci jedné hostitelské buňky O. tsutsugamushi se rychle dělí na mnoho jednotlivců, jak ukazuje Obrázek 1. Jednobuněčný organismus má oválný tvar a měří 0,5 až 0,8 µm široký a 1,2 až 3,0 µm dlouhý. Kvůli podobnosti byl dříve zařazen do rodu Rickettsia mimo jiné bakterie, ale později jim byl přidělen samostatný rod, Orientia,[20] kterou sdílí (od roku 2010) pouze s Candidatus Orientia chuto.[21] Je širší, ale kratší než jiné rickettsiální bakterie, které mají tvar tyčinky a průměrně měří 0,25 až 0,3 µm a 0,8 až 1 µm.[22] Během reprodukce rozdělí (o binární dělení ) do dvou dceřiných buněk procesem pučení. Zatímco prochází pučením, hromadí se na povrchu hostitelské buňky, na rozdíl od jiných bakterií. Jeden úplný začínající cyklus trvá 9 až 18 hodin.[23]
Struktura O. tsutsugamushi (odhalil transmisní elektronová mikroskopie ) je zobrazen v Obrázek 2. Bakterie je obklopena a buněčná stěna na vnější straně a buněčné membráně na vnitřní straně. Pokrytí buněk absorbuje skvrny, jako jsou Giemsa a Gimenezovy skvrny. Ačkoli má její buněčná stěna klasickou bakteriální dvojitou vrstvu, její vnější leták je mnohem silnější než vnitřní, což je v Rickettsia druh.[24] Chybí vrstva tobolky, která tvoří sférické halo v jiných bakteriích. Buněčná stěna je méně tuhá kvůli nepřítomnosti peptidoglykanu, který je jinak charakteristický pro tuhé buněčné stěny jiných bakterií. Klasické bakteriální lipofosfoglykany jako např kyselina muramová, glukosamin, hydroxymastné kyseliny, heptosa a kyselina 2-keto-3-deoxyoktonová také chybí v buněčné stěně. Z důvodu absence peptidoglykan je bakterie přirozeně odolná vůči všem β-laktam antibiotika (např penicilin ), ke kterému Rickettsia druhy jsou obvykle citlivé na.[25] Jeho genom zcela postrádá geny pro syntézu lipofosfoglykanů, ale obsahuje některé pro peptidoglykany. Důležité geny nezbytné pro peptidoglykanovou systézu, jako je alr, dapF a PBP1 chybí: alr kóduje enzym L-alanin racemázu, který převádí L-alanin na D-alanin v prvním kroku syntézy peptidoglykanu; dapF kóduje diaminopimelát epimerázu, která převádí LL-2,6-diaminoheptandioát (L, L-DAP) na meso-diaminoheptandioát (meso-DAP); a PBP1 kóduje protein-1 vázající penicilin (PBP1), který převádí periplazmatický lipid II na peptidoglykan. Bakterie tedy nemůže syntetizovat typickou buněčnou stěnu peptidoglykanu a místo toho vytváří na svém povrchu strukturu podobnou peptidoglykanu.[26] Buněčná membrána je také chemicky odlišná ve svém složení bílkovin a tento rozdíl vede ke změnám kmene uvnitř samotného druhu.[27] Cytoplazma je čirá a vykazuje zřetelnou DNA a ribozomy.
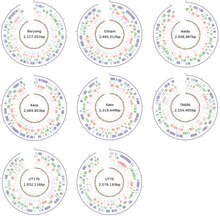
Bakterie je vysoce virulentní, takže její izolace a buněčná kultura se provádí pouze v laboratorním zařízení s úroveň biologické bezpečnosti 3. Na rozdíl od jiných bakterií, které mohou snadno růst na různých kultivačních médiích, lze rickettsiales kultivovat pouze v živých buňkách. O. tsutsugamushi konkrétně lze pěstovat pouze ve žloutkových váčcích vyvíjejících se kuřecích embryí a v kultivovaných buněčných liniích, jako je HeLa, BHK, Vero, a L929.[28] Na rozdíl od Rickettsia druhy, které sídlí v jádru hostitelské buňky, O. tsutsugamushi většinou roste v cytoplazmě hostitelské buňky.[19] Geneticky se liší od ostatních Rickettsia pouze o 9%.[29] I když adaptace na povinný intracelulární parazitismus mezi bakteriemi má obecně za následek redukovaný genom, má velikost genomu asi 2,0–2,7 Mb v závislosti na kmenech (Obrázek 3), který je poměrně větší než u jiných rickettsiales - dvakrát větší než u rickettsiales Rickettsia prowazekii,[30] nejznámější člen. Celý genom je distribuován v jediném kruhovém chromozomu. Sekvence celého genomu jsou k dispozici pouze pro kmeny Ikeda a Boryong, obě z Korejské republiky. Genom kmene Ikeda je dlouhý 2 008 987 párů bází (bp) dlouhý a obsahuje 1 967 genů kódujících proteiny.[31] Kmen Boryong je větší s 2 127 051 bp a 2 179 genů kódujících proteiny.[32]
Porovnání genomu ukazuje pouze 657 základních genů mezi různými kmeny.[33] S přibližně 42–47% opakujících se sekvencí O. tsutsugamushi má nejvíce opakovaný bakteriální genom sekvenovaný od roku 2013.[34] Opakovaná sekvence DNA zahrnuje krátkou opakující se sekvence, transponovatelné prvky (včetně prvků vkládací sekvence, miniaturních transponovatelných prvků s obráceným opakováním, a Intron skupiny II ) a značně zesílený integrační a konjugativní prvek (ICE) nazývaný rickettsiální zesílený genetický prvek (RAGE).[32] RAGE se také nachází v jiných rickettsiálních bakteriích. v O. tsutsugamushiRAGE však obsahuje řadu genů včetně tra geny typické pro sekreční systémy typu IV a gen pro ankyrin opakovat –Obsahující bílkoviny. Ankyrinové proteiny obsahující repetice jsou vylučovány sekrečním systémem typu I do hostitelské buňky. Přesná role sekrečního systému typu IV v O. tsutsugamushi není známo. Může být zapojen do horizontální přenos genů mezi různými kmeny.[1]
Životní cyklus a přenos
Orientia tsutsugamushi se přirozeně přenáší v populaci roztočů patřících do rodu Leptotrombidium. Může být přenášena ženou na vajíčka prostřednictvím procesu zvaného transovariální přenos a z vajíček na larvy a dospělé procesem transstadiálního přenosu. Životní cyklus bakterií je tedy udržován výhradně u roztočů. Infekce u hlodavců a lidí je náhodný přenos z kousnutí larev roztočů a není nutný pro reprodukci nebo přežití bakterie. Ve skutečnosti je u lidí přenos zastaven a bakterie se dostává do slepé uličky.[27] Neinfikovaní roztoči však mohou infekci získat od infikovaných hlodavců.[35]
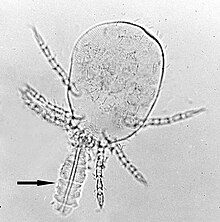
U infekcí hlodavců a lidí Leptotrombidium deliense je nejběžnějším vektorem O. tsutsugamushi. L. pallidum, L. fletcheri a L. scutellare jsou také dopravci v mnoha zemích. Navíc, L. akamushi je endemický dopravce v Japonsku, L. chiangraiensis a L. imphalum v Thajsku, L. gaohuensis v Číně a L. arenicola v Malajsii a Indonésii.[3] V některých částech Indie, jiný druh roztočů, Schoengastiella ligula je také hlavním vektorem.[36] Životní cyklus roztočů se skládá z vajíčka, prelarvy, larvy, protonymph, deutonymph, tritonymph a dospělého. Larvy, běžně označované jako chiggers (Obrázek 4), jsou jediným ektoparazitickým stádiem, které se živí tělními tekutinami hlodavců a jiných oportunistických savců. Jsou tedy jedinou fází v životě roztočů, kteří přenášejí infekci. Divoké krysy rodu Ratus jsou hlavními přirozenými hostiteli chiggerů.[37] Chiggers se živí savčím hostitelem pouze jednou. Krmení obvykle trvá 2 až 4 dny. Na rozdíl od většiny parazitů se nekrmí krví, ale místo toho tělní tekutinou přes vlasové folikuly nebo póry kůže. Mají speciální podávací zařízení zvané a Stylostome, což je trubice tvořená ztuhlými slinami. Jejich sliny mohou rozpustit hostitelskou tkáň kolem místa krmení, takže pohlcují zkapalněnou tkáň. O. tsutsugamushi je přítomen v slinné žlázy roztočů a během tohoto krmení se uvolňuje do hostitelské tkáně.[38]
Buněčná invaze
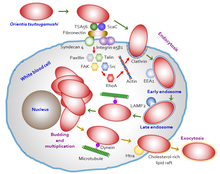
Orientia tsutsugamushi zpočátku útočí na myelocyty (mladé bílé krvinky) v oblasti očkování a poté endoteliální buňky obložení vaskulatura. Proces buněčné invaze je uveden v Obrázek 5. V krevním oběhu se zaměřuje na profesionální fagocyty („pojídače buněk“, bílé krvinky), jako jsou dendritické buňky a makrofágy ve všech orgánech jako sekundární cíle. Parazit se nejprve připojí k cílovým buňkám pomocí povrchových proteoglykanů přítomných na hostitelské buňce a bakteriálních povrchových proteinů, jako je typově specifický protein 56 (nebo typově specifický antigen, TSA56) a antigeny povrchových buněk (ScaA a ScaC, které jsou membránovými transportními proteiny) .[39][40] Tyto proteiny interagují s hostitelem fibronektin vyvolat fagocytóza (proces požití bakterie). Schopnost skutečně vstoupit do hostitelské buňky závisí na integrin - zprostředkovaná signalizace a reorganizace aktin cytoskelet.[41]
Orientia tsutsugamushi má speciální adaptaci pro přežití v hostitelské buňce vyhýbáním se imunitní reakci hostitele. Jakmile interaguje s hostitelskými buňkami, způsobí, že membrána hostitelských buněk vytvoří dopravní bublinu zvanou a vezikul potažený klatrinem kterým se transportuje do cytoplazmy. Uvnitř cytoplazmy vystupuje z vezikuly (nyní známé jako endozom ) před zničením endozomu (tzv autofagie ) podle lysozomy.[42] Poté se pohybuje směrem k jádru, konkrétně v perinukleární oblasti, kde začíná růst a množit se. Na rozdíl od jiných blízce příbuzných bakterií, které využívají pro pohyb v cytoplazmě procesy zprostředkované aktiny (tzv intracelulární obchodování nebo transport ), O. tsutsugamushi je neobvyklé při používání mikrotubul - zprostředkované procesy podobné těm, které používají viry, jako je adenoviry a viry herpes simplex. Dále únik (exocytóza ) z infikované hostitelské buňky je také neobvyklý. Vytváří další vezikul pomocí membrány hostitelské buňky, vytváří malý pupen a uvolňuje se z povrchu hostitelské buňky, zatímco je stále uzavřený ve vezikulu. Bakterie vázaná na membránu je tvořena interakcí mezi lipidovými rafty bohatými na cholesterol a také HtrA, proteinem 47 kDa na bakteriálním povrchu.[43] Proces pučení a význam bakterie vázané na membránu však dosud nejsou známy.
Kmeny
Orientia tsutsugamushi je rozmanitý druh bakterií. Ida A. Bengtson z Služba veřejného zdraví USA byl první, kdo si všiml existence různých kmenů pomocí interakce antigen-protilátka (test fixace komplementu ) v roce 1944.[44] Poznamenala, že různé kmeny mají různý stupeň virulence a že protilátky v krevních sérech pacientů zkříženě reagují na různé kmeny. Do roku 1946 zjistila, že existují tři hlavní kmeny (sérotypy), jmenovitě Karp (z Nové Guineje), Gilliam (z Indie) a Seerangay (z britské Malajska).[45] Akira Shishido popsal kmen Kato, kromě Gilliam a Karp, v Japonsku v roce 1958.[46] Od té doby bylo rozpoznáno šest základních antigenních kmenů, jmenovitě Gilliam, Karp, Kato, Shimokoshi, Kawasaki a Kuroki. Karp je nejhojnější kmen, který představuje asi 50% všech infekcí.[3] V Koreji je hlavním kmenem Boryong.[47] Od roku 2009 bylo u lidí založeno více než 20 různých kmenů na základě antigenní variace pomocí sérologických testů, jako je fixace komplementu a imunofluorescenční test.[3] Počet je mnohem vyšší, pokud se zohlední kmeny hlodavců a roztočů. Například studie v Japonsku v roce 1994 uvádí 32 kmenů, 14 od lidských pacientů, 12 od divokých hlodavců a 6 od roztočů trombiculidů. Různé kmeny vyvíjejí různé úrovně virulence a nejvirulentnější je KN-3, který převládá mezi divokými hlodavci.[48] Další studie z roku 1996 uvádí 40 kmenů.[49] Genetické metody odhalily ještě větší složitost, než byla dříve popsána (například Gilliam se dále dělí na typy Gilliam a JG). Vzhledem k imunologickým rozdílům sérotypů je možná současná a opakovaná infekce různými kmeny.[50][51]
Antigenní variace
Orientia tsutsugamushi má čtyři hlavní proteiny povrchové membrány (antigeny ) mající molekulové hmotnosti 22 kDa, 47 kDa, 56 kDa a 110 kDa. Specifický antigen typu 56 kDa (TSA56) je nejdůležitější, protože jej neprodukují žádné jiné bakterie a je zodpovědný za vytváření genetické rozmanitosti v různých kmenech.[52] Představuje asi 10–15% celkových buněčných bílkovin. Antigeny 22-kDa, 47-kDa nebo 110-kDa nejsou kmenově specifické, takže TSA56 je hlavním cílem sofistikovaných diagnostických testů, jako je imunoblot, ELISA a analýza DNA.[53] Protein pomáhá adhezi a vstupu bakterie do hostitelských buněk, stejně jako vyhýbání se imunitní reakci hostitele. Liší se velikostí od 516 do 540 aminokyselinových zbytků mezi různými kmeny a jeho gen je dlouhý přibližně 1550 párů bází. Jeho gen obsahuje čtyři hypervariabilní oblasti, což naznačuje, že syntetizuje mnoho antigenně odlišných proteinů.[49] Uvnitř bakterie jsou také proteiny 11-kDa a 60-kDa, které jsou si velmi podobné GroES a GroEL bakterie Escherichia coli, ale ne to z Rickettsia druh.[54] GroES a GroEL jsou proteiny tepelného šoku patřící do rodiny molekulární chaperony v bakteriích. Analýzy DNA ukázaly, že GroES a GroEL geny jsou skutečně přítomny v O. tsutsugamushi s mírnou variací v různých kmenech a produkují proteiny 11-kDa a 60-kDa.[55]
Choroba
Orientia tsutsugamushi způsobuje složité a potenciálně život ohrožující onemocnění známé jako tyfus. Infekce začíná, když si chiggers během krmení kousnou na kůži. Bakterie se ukládají v místě krmení (inokulace), kde se množí. Způsobují progresivní poškození tkáně (nekróza ), což vede k tvorbě eschar na kůži. Nekróza postupuje k zánětu cév, tzv vaskulitida. To zase způsobuje zánět lymfatických uzlin, tzv lymfadenopatie. Během několika dnů se vaskulitida rozšíří do různých orgánů včetně jater, mozku, ledvin, mozkové pleny a plíce.[56] Toto onemocnění je zodpovědné za téměř čtvrtinu všech horečnatých onemocnění v endemických oblastech. Úmrtnost v závažných případech nebo v důsledku nesprávného zacházení nebo nesprávné diagnózy může být až 30–70%.[57] Asi 6% infikovaných lidí zemře neléčeno a 1,4% pacientů zemře dokonce i při léčbě. Kromě toho může být míra úmrtí u léčených pacientů až 14% s neurologickými problémy a 24% s multiorgánovou dysfunkcí.[58] V případech chybné diagnózy a selhání léčby se rychle vyvinou systémové komplikace včetně syndrom akutní dechové tísně akutní selhání ledvin, encefalitida, gastrointestinální krvácení, hepatitida, meningitida, myokarditida, pankreatitida, zápal plic, septický šok, subakutní tyroiditida, a syndrom dysfunkce více orgánů.[59] Škodlivé účinky zahrnující selhání více orgánů a neurologické poškození se obtížně léčí a mohou způsobit celoživotní oslabení nebo mohou být přímo smrtelné.[59] Centrální nervový systém je často ovlivněn a vede k různým komplikacím včetně cerebellitis, paralýzy hlavových nervů, meningoencefalitida, plexopatie, příčná myelitida, a Guillan-Barré syndrom.[60] Míra úmrtí v důsledku komplikací může být až 14% u mozkových infekcí a 24% u selhání více orgánů.[58] V Indii se tyfus drhnutí stal hlavní příčinou syndromu akutní encefalitidy, který byl dříve způsoben hlavně virovou infekcí, Japonská encefalitida.[61]
Epidemiologie

The Světová zdravotnická organizace v roce 1999 uvedl, že:
Scrub tyfus je pravděpodobně jedním z nejvíce nedostatečně diagnostikovaných a nedostatečně hlášených horečnatých onemocnění vyžadujících hospitalizaci v regionu. Absence definitivních příznaků a symptomů v kombinaci s obecnou závislostí na sérologických testech činí diferenciaci tyfového drhnutí od jiných běžných horečnatých onemocnění, jako je myší tyfus, tyfus a leptospiróza, docela obtížné.[62]
Scrub týfus je historicky endemický pro asijsko-pacifickou oblast, pokrývající ruský Dálný východ a Koreu na severu, severní Austrálii na jihu a Afghánistán na západě, včetně ostrovů západního Tichého oceánu, jako je Japonsko, Tchaj-wan, Filipíny , Papua Nová Guinea, Indonésie, Srí Lanka a indický subkontinent. Tato zeměpisná oblast se populárně nazývá Tsutsugamushi Triangle, jak je znázorněno na Obrázek 6.[56] Šíří se však do Afriky, Evropy a Jižní Ameriky.[63] Odhaduje se, že miliarda lidí bude kdykoli vystavena riziku infekce a v Tsutsugamushi Triangle se každý rok vyskytne v průměru jeden milion případů.[38] Zátěž drhnout tyfus ve venkovských oblastech Asie je obrovská, představuje až 20% horečnaté nemoci v nemocnici a séroprevalence (pozitivní infekce pomocí krevních testů) u 50% populace.[64] Více než pětina populace nese bakteriální protilátky, tj. Byly infikovány, v endemických oblastech. Nejvyšší míra výskytu je v Jižní Koreji (s nejvyššími 59,7 infekcemi ze 100 000 lidí v roce 2013), následuje Japonsko, Thajsko a Čína v horní části seznamu. Největší riziko infekce je ve věkové skupině 60–69 let. Vyšší infekce (57,3%) je pozorována u žen ve srovnání s muži (42,7%). Nejzranitelnější jsou zemědělci, kteří představují 70% případů v Číně. Toto onemocnění je častější ve venkovských oblastech, ale v městských oblastech dochází k rychlému nárůstu. Například v Koreji se mezi lety 2003 a 2013 v metropolitních oblastech zvýšil roční výskyt 21krát.[58]
Jihilská odrůda
Je známo, že v Chile se nemoc vyskytuje zejména v jižní polovině země Ostrov Chiloé a fjordy a kanály Patagonie. V letech 2015 až 2020 bylo v Chile celkem 40 známých případů, kdy se většina postižených hlásila k práci při sběru a těžbě palivového dřeva.[65] Podle chilského vědce Katie Abarcy nemoc v Chile odpovídá endemické rozmanitosti bakterií, a proto nepřišla z „Tsutsugamushi Triangle“.[65] Skutečnost, že tato choroba byla poprvé hlášena v roce 2006 v Chile, je pravděpodobně výsledkem nedostatečné diagnózy a nedostatečného hlášení.[65] V lednu 2020 byla nemoc poprvé hlášena v roce Nejjižnější region Chile.[65]
Diagnóza
Příznak

Hlavním příznakem O. tsutsugamushi infekce je vysoká (horečnatá) horečka; symptom je však podobný jiným tropickým chorobám přenášeným vektory, jako je malárie, leptospiróza, tyfus, myší tyfus, chikungunya, a horečka dengue.[66][67] Díky tomu je přesná klinická diagnóza obtížná, což často vede k chybné diagnóze. Prvotními indikacemi jsou horečka se zimnicí spojená s bolestmi hlavy, bolesti svalů (myalgie ), pocení a zvracení. Vzhled příznaků (inkubační doba ) trvá mezi 6 a 21 dny.[56] Jednoduchou vizuální diagnózou je přítomnost zapálené jizvy podobné jizvě zvané eschar, která je považována za „nejužitečnější diagnostickou stopu u pacientů s akutním horečnatým onemocněním“. Eschar se tvoří na kůži, kde se infikovaný roztoč kousne, obvykle v podpaží, slabinách nebo v jakékoli břišní oblasti (Obrázek 7). Ve vzácných případech je to vidět na tváři, ušním lalůčku a hřbet nohou.[68] Problém však je, že eschar není vždy přítomen; v nejvyšším záznamu pouze 55% pacientů s křovinatým tyfem mělo během vypuknutí v jižní Indii eschar.[69] Eschar také není specifický pro drhnout tyfus, vyskytující se v další rickettsiální nemoci jako Skalní hora skvrnitá horečka,[70] Brazilská skvrnitá horečka,[71] a tyfus indický.[72][73] Pomocí analýzy DNA pokročilou polymerázovou řetězovou reakcí lze z escharů identifikovat různé rickettsiální infekce.[74][75]
Krevní test
Podezření na infekci je potvrzeno sérologickými testy. O. tsutsugamushi je nejčastěji detekován z krevního séra pomocí Weil – Felixův test. Weil – Felix je nejjednodušší a nejrychlejší test, ale není citlivý ani konkrétní, protože detekuje jakýkoli druh rickettsiální infekce. Používají se citlivější testy, jako je rychlý imunochromatografický test (RICT), imunofluorescenční testy (IFA), ELISA a analýza DNA pomocí polymerázové řetězové reakce (PCR).[37][28] IFA je považován za test zlatého standardu, protože poskytuje spolehlivý výsledek; je však drahý a není specifický pro různé rickettsiální bakterie.[76] ELISA a PCR mohou detekovat O. tsutsugamushi-specifické proteiny, jako jsou TSA56 a GroEL, takže jsou vysoce specifické a citlivé.[77] Na druhou stranu jsou to vysoce sofistikované a drahé techniky.
Léčba
Orientia tsutsugamushi infekce může být léčena antibiotiky, jako je azithromycin, chloramfenikol, doxycyklin, rifampicin, roxithromycin, a tetracyklin. Doxycyklin je nejčastěji používaný a je považován za lék volby díky své vysoké účinnosti a rychlému působení. Ale u těhotných žen a kojenců ano kontraindikováno a azithromycin je lék volby. V jihovýchodní Asii, kde došlo k rezistenci na doxycyklin a chloramfenikol, se doporučuje azithromycin pro všechny pacienty.[78] Randomizovaná kontrolovaná studie a systematický přehled ukázaly, že azithromycin je nejbezpečnější lék.[79][80]
Vakcína
Žádná licence O. tsutsugamushi vakcíny jsou v současné době k dispozici. První vakcíny byly vyvinuty koncem 40. let 20. století, ale v klinických studiích selhaly.[81][82] Samotný jedinečný TSA56, který je považován za ideální cíl, je velmi variabilní svým chemickým složením v různých kmenech. Účinná vakcína pro jeden kmen není pro jiný užitečná. Ideální vakcína by měla chránit všechny místní kmeny. Tato složitost znesnadňuje výrobu použitelné vakcíny.[83] Vakcína zaměřená na protein vnější membrány 47-kDa (OMP) je slibným kandidátem s experimentálním úspěchem u myší proti kmenu Boryong.[84] Kombinované cílení na TSA56 a ScaA je také dobrým kandidátem na infekci smíšeným kmenem.[40]
Imunita
Neexistuje žádná trvalá imunita O. tsutsugamushi infekce. Antigenní variace brání rozvoji zkřížené imunity vůči různým kmenům O. tsutsugamushi. U infikovaného jedince se může krátkodobě vyvinout imunita, která však po několika měsících vymizí a imunita vůči jednomu kmenu neposkytuje imunitu druhému.[83] V roce 1950 byl proveden imunizační experiment, při kterém se u 16 dobrovolníků infekce rozvinula i po 11–25 měsících primární infekce.[85] Nyní je známo, že dlouhověkost imunity závisí na kmenech bakterie. Pokud dojde k reinfekci stejným kmenem jako u předchozí infekce, může u opic existovat imunita po dobu 5–6 let.[86] Ale u lidí imunita klesá po jednom roce a zmizí do dvou let.[87]
Reference
![]() Tento článek byl odeslán na adresu WikiJournal of Medicine pro externí akademické vzájemné hodnocení v roce 2018 (zprávy recenzenta ). Aktualizovaný obsah byl znovu integrován na stránku Wikipedie pod a CC-BY-SA-3.0 licence (2019 ). Verze záznamu v revidovaném znění je: Kholhring Lalchhandama; et al. (2019), „Orientia tsutsugamushi, agentka drhnutí tyfu“, WikiJournal of Medicine, 6 (1): 4, doi:10.15347 / WJM / 2019.004, ISSN 2002-4436, Wikidata Q73054107
Tento článek byl odeslán na adresu WikiJournal of Medicine pro externí akademické vzájemné hodnocení v roce 2018 (zprávy recenzenta ). Aktualizovaný obsah byl znovu integrován na stránku Wikipedie pod a CC-BY-SA-3.0 licence (2019 ). Verze záznamu v revidovaném znění je: Kholhring Lalchhandama; et al. (2019), „Orientia tsutsugamushi, agentka drhnutí tyfu“, WikiJournal of Medicine, 6 (1): 4, doi:10.15347 / WJM / 2019.004, ISSN 2002-4436, Wikidata Q73054107
- ^ A b Salje, J .; Kline, K.A. (2017). "Orientia tsutsugamushi: Zanedbaný, ale fascinující obligátní intracelulární bakteriální patogen “. PLOS patogeny. 13 (12): e1006657. doi:10.1371 / journal.ppat.1006657. PMC 5720522. PMID 29216334.
- ^ Watt, G .; Parola, P. (2003). "Drhnout tyfus a tropické rickettsiosy". Aktuální názor na infekční nemoci. 16 (5): 429–436. doi:10.1097/00001432-200310000-00009. PMID 14501995. S2CID 24087729.
- ^ A b C d Kelly, D.J .; Fuerst, P.A .; Ching, W.M .; Richards, A.L. (2009). "Děrovitý tyfus: zeměpisné rozložení fenotypových a genotypových variant viru Orientia tsutsugamushi". Klinické infekční nemoci. 48 Suppl (Suppl): S203 – S230. doi:10.1086/596576. PMID 19220144.
- ^ Yamamoto, S .; Kawabata, N .; Tamura, A .; Urakami, H .; Ohashi, N .; Murata, M .; Yoshida, Y .; Kawamura A, Jr. (1986). "Imunologické vlastnosti Rickettsia tsutsugamushi, Kmen Kawasaki, izolovaný od pacienta v Kjúšú ". Mikrobiologie a imunologie. 30 (7): 611–620. doi:10.1111 / j.1348-0421.1986.tb02988.x. PMID 3095612. S2CID 2968436.
- ^ Fan, M.Y .; Walker, D.H .; Yu, S.R .; Liu, Q.H. (1987). „Epidemiologie a ekologie rickettsiálních chorob v Čínské lidové republice“. Recenze infekčních nemocí. 9 (4): 823–840. doi:10.1093 / clinids / 9.4.823. PMID 3326129.
- ^ Kawamura, R. (1926). Studie o nemoci tsutsugamushi (japonská horečka krve). Cincinnati, OH (USA): mluvčí tiskové společnosti. p. 2.
- ^ Palm, T.A. (1878). „Někteří obyvatelé Japonska považují chorobu zvanou„ shima-mushi “nebo„ nemoc ostrovního hmyzu “; věří se, že je pro tuto zemi zvláštní a dosud nebyla popsána.“. Edinburgh Medical Journal. 24 (2): 128–132. PMC 5317505. PMID 29640208.
- ^ Miyajima, M .; Okumura, T. (1917). „O životním cyklu„ Akamushi “nositele horečky řeky Nippon.“ Kitasato Archives of Experimental Medicine. 1 (1): 1–14.
- ^ Hirst, S. (1915). „Na Tsutsugamushi (Microtrombidium akamushi, Brumpt), nositel japonské říční horečky ". Journal of Economic Biology. 10 (4): 79–82.
- ^ Nagayo, M. (1917). „Na vílu a prosopon tsutsugamushi, Leptotrombidium akamushi, N. Sp. (Trombidium akamushi Brumpt), nositel tsutsugamushiho choroby “. Journal of Experimental Medicine. 25 (2): 255–272. doi:10.1084 / jem.25.2.255. PMC 2125768. PMID 19868085.
- ^ Lalchhandama, K. (2018). „Sága křovinatého tyfu s poznámkou o ohniscích v Mizoramu“. Vědecká vize. 18 (2): 50–57. doi:10,33493 / scivis.18.02.01.
- ^ Hayashi, N. (1920). "Etiologie tsutsugamushi choroby". Parazitologický časopis. 7 (2): 52–68. doi:10.2307/3270957. JSTOR 3270957.
- ^ Nagayo, M .; Tamiya, T .; Mitamura, T .; Sato, K. (1930). „K viru tsutsugamushiho choroby a jeho prokázání novou metodou“. Jikken Igaku Zasshi (Japonský žurnál experimentální medicíny). 8 (4): 309–318.
- ^ Nagayo, M .; Tamiya, T .; Mitamura, T .; Sato, K. (1930). „Sur le virus de la maladie de Tsutsugamushi [O viru tsutsugamushi]“. Comptes Rendus des Séances de la Société de Biologie. 104: 637–641.
- ^ da Rocha Lima, H. (1916). „Untersuchungen über fleckfleber [Researches on týfus]“. Münchener medizinische Wochenschrift. 63 (39): 1381–1384.
- ^ Ogata, N. (1931). „Aetiologie der Tsutsugamushi-krankheit: Rickettsia tsutsugamushi [Etiologie choroby tsutsugamushi: Rickettsia tsutsugamushi". Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. 122: 249–253.
- ^ Kawamüra, R .; Imagawa, Y. (1931). „Ueber die Proliferation der patogenen Rickettsia im tierischen organismus bei der tsutsugamushi-krankheit [Násobení Rickettsia patogen choroby tsutsugamushi u zvířat] ". Transakce Japonské společnosti pro patologii. 21: 455–461.
- ^ Kawamüra, R .; Imagawa, Y. (1931). „Die feststellung des erregers bei der tsutsugamushikrankheit [Potvrzení infekčního agens u tsutsugamushi choroby]“. Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. 122 (4/5): 253–261.
- ^ A b Tamura, A .; Urakami, H .; Ohashi, N. (1991). "Srovnávací pohled na Rickettsia tsutsugamushi a další skupiny Rickettsiae “. European Journal of Epidemiology. 7 (3): 259–269. doi:10.1007 / BF00145675. PMID 1909244. S2CID 12298540.
- ^ A b Tamura, A .; Ohashi, N .; Urakami, H .; Miyamura, S. (1995). "Klasifikace Rickettsia tsutsugamushi v novém rodu Orientia gen. nov., as Orientia tsutsugamushi Hřeben. listopad". International Journal of Systematic Bacteriology. 45 (3): 589–591. doi:10.1099/00207713-45-3-589. PMID 8590688.
- ^ Izzard, L (2010). „Izolace románu Orientia druh (O. chuto sp. listopadu) od pacienta nakaženého v Dubaji “. Journal of Clinical Microbiology. 48 (12): 4404–4409. doi:10.1128 / JCM.01526-10. PMC 3008486. PMID 20926708.
- ^ Sunyakumthorn, P .; Bourchookarn, A .; Pornwiroon, W .; David, C .; Barker, S. A .; Macaluso, K. R. (2008). "Charakterizace a růst polymorfu Rickettsia felis v klíšťové buněčné linii ". Aplikovaná a environmentální mikrobiologie. 74 (10): 3151–3158. doi:10.1128 / AEM.00025-08. PMC 2394910. PMID 18359823.
- ^ Moree, M.F .; Hanson, B. (1992). "Růstové charakteristiky a proteiny kmenů purifikovaných z plaků Rickettsia tsutsugamushi". Infekce a imunita. 60 (8): 3405–3415. doi:10.1128 / IAI.60.8.3405-3415.1992. PMC 257328. PMID 1379212.
- ^ Silverman, D.J .; Wisseman, C.L. Jr. (1978). "Srovnávací ultrastrukturální studie na obálkách buněk Rickettsia prowazekii, Rickettsia rickettsii, a Rickettsia tsutsugamushi". Infekce a imunita. 21 (3): 1020–1023. doi:10.1128 / IAI.21.3.1020-1023.1978. PMC 422098. PMID 101465.
- ^ Amano, K .; Tamura, A .; Ohashi, N .; Urakami, H .; Kaya, S .; Fukushi, K. (1987). "Nedostatek peptidoglykanových a lipopolysacharidových složek v Rickettsia tsutsugamushi". Infekce a imunita. 55 (9): 2290–2292. doi:10.1128 / IAI.55.9.2290-2292.1987. PMC 260693. PMID 3114150.
- ^ Atwal, S .; Giengkam, S .; Chaemchuen, S .; Dorling, J .; Kosaisawe, N .; VanNieuwenhze, M .; Sampattavanich, S .; Schumann, P .; Salje, J. (2017). "Důkazy o struktuře podobné peptidoglykanu v Orientia tsutsugamushi". Molekulární mikrobiologie. 105 (3): 440–452. doi:10,1111 / mmi.13709. PMC 5523937. PMID 28513097.
- ^ A b Lalchhandama, K. (2017). „Rickettsióza jako kritická nově se objevující infekční nemoc v Indii“. Vědecká vize. 17 (4): 250–259. doi:10,33493 / scivis.17.04.09.
- ^ A b Koh, G.C .; Maude, R.J .; Paris, D.H .; Newton, P.N .; Blacksell, S.D. (2010). „Diagnóza křovinatého tyfu“. American Journal of Tropical Medicine and Hygiene. 82 (3): 368–370. doi:10.4269 / ajtmh.2010.09-0233. PMC 2829893. PMID 20207857.
- ^ Ohashi, N .; Fukuhara, M .; Shimada, M .; Tamura, A. (1995). "Fylogenetická poloha Rickettsia tsutsugamushi a vztah mezi jeho antigenními variantami analýzou genových sekvencí 16S rRNA ". Mikrobiologické dopisy FEMS. 125 (2–3): 299–304. doi:10.1111 / j.1574-6968.1995.tb07372.x. PMID 7875578.
- ^ Bishop-Lilly, K.A .; Ge, H .; Butani, A .; Osborne, B .; Verratti, K .; Mokashi, V .; Nagarajan, N .; Pop, M .; Přečtěte si, T.D .; Richards, A.L. (2013). "Sekvenování genomu čtyř kmenů Rickettsia prowazekii, původce epidemického tyfu, včetně jednoho izolátu létající veverky ". Oznámení o genomu. 1 (3): e00399-13. doi:10.1128 / genomA.00399-13. PMC 3695431. PMID 23814035.
- ^ Nakayama, K .; Yamashita, A .; Kurokawa, K .; Morimoto, T .; Ogawa, M .; Fukuhara, M .; Urakami, H .; Ohnishi, M .; Uchiyama, I .; Ogura, Y .; Ooka, T .; Oshima, K .; Tamura, A .; Hattori, M .; Hayashi, T. (2008). „Celá genomová sekvence obligátní intracelulární bakterie Orientia tsutsugamushi odhalila masivní amplifikaci genů během redukčního vývoje genomu ". Výzkum DNA. 15 (4): 185–199. doi:10.1093 / dnares / dsn011. PMC 2575882. PMID 18508905.
- ^ A b Cho, N.-H .; Kim, H.-R .; Lee, J.-H .; Kim, S.-Y .; Kim, J .; Cha, S .; Kim, S.-Y .; Darby, A. C .; Fuxelius, H.-H .; Yin, J .; Kim, J. H .; Kim, J .; Lee, S. J .; Koh, Y.-S .; Jang, W.-J .; Park, K.-H .; Andersson, S. G. E .; Choi, M.-S .; Kim, I.-S. (2007). „The Orientia tsutsugamushi genom odhaluje masivní množení konjugativního sekrečního systému typu IV a genů interakce hostitel-buňka ". Sborník Národní akademie věd. 104 (19): 7981–7986. Bibcode:2007PNAS..104.7981C. doi:10.1073 / pnas.0611553104. PMC 1876558. PMID 17483455.
- ^ Batty, E.M .; Chaemchuen, S .; Blacksell, S .; Richards, A.L .; Paris, D .; Bowden, R .; Chan, C .; Lachumanan, R .; Den, N .; Donnelly, P .; Chen, S .; Salje, J .; Reck, J. (2018). „Dlouhé čtení celého genomu a srovnávací analýza šesti kmenů lidského patogenu Orientia tsutsugamushi". PLOS opomíjené tropické nemoci. 12 (6): e0006566. doi:10.1371 / journal.pntd.0006566. PMC 6005640. PMID 29874223.
- ^ Viswanathan, S .; Muthu, V .; Iqbal, N .; Remalayam, B .; George, T (2013). „Drhněte tyfusovou meningitidu v jižní Indii - retrospektivní studie“. PLOS ONE. 8 (6): e66595. Bibcode:2013PLoSO ... 866595V. doi:10,1371 / journal.pone.0066595. PMC 3682970. PMID 23799119.
- ^ Takhampunya, R .; Korkusol, A .; Promsathaporn, S .; Tippayachai, B .; Leepitakrat, S .; Richards, A.L .; Davidson, S.A .; Malý, P.L.C. (2018). "Heterogenita Orientia tsutsugamushi genotypy v polních sebraných trombikulidních roztočích od volně žijících malých savců v Thajsku “. PLOS opomíjené tropické nemoci. 12 (7): e0006632. doi:10.1371/journal.pntd.0006632. PMC 6062101. PMID 30011267.
- ^ Tilak, R.; Wankhade, U.; Kunwar, R.; Tilak, V.W. (2011). „Vznik Schoengastiella ligula as the vector of scrub typhus outbreak in Darjeeling: Has Leptotrombidium deliense been replaced?". Indian Journal of Public Health. 55 (2): 92–99. doi:10.4103/0019-557X.85239. PMID 21941043.
- ^ A b Luce-Fedrow, A.; Lehman, M.; Kelly, D.; Mullins, K.; Maina, A.; Stewart, R.; Ge, H.; John, H .; Jiang, J .; Richards, Allen (2018). "A review of scrub typhus (Orientia tsutsugamushi and related organisms): then, now, and tomorrow". Tropická medicína a infekční choroby. 3 (1): 8. doi:10.3390/tropicalmed3010008. PMC 6136631. PMID 30274407.
- ^ A b Xu, G .; Walker, D.H.; Jupiter, D .; Melby, P.C.; Arcari, C.M.; Day, N.P. (2017). "A review of the global epidemiology of scrub typhus". PLOS opomíjené tropické nemoci. 11 (11): e0006062. doi:10.1371/journal.pntd.0006062. PMC 5687757. PMID 29099844.
- ^ Ge, Y .; Rikihisa, Y. (2011). "Subversion of host cell signaling by Orientia tsutsugamushi". Mikroby a infekce. 13 (7): 638–648. doi:10.1016/j.micinf.2011.03.003. PMID 21458586.
- ^ A b Ha, N.Y.; Sharma, P .; Kim, G .; Kim, Y .; Min, C.K.; Choi, M.S.; Kim, I.S.; Cho, N.H. (2015). "Immunization with an autotransporter protein of Orientia tsutsugamushi provides protective immunity against scrub typhus". PLOS opomíjené tropické nemoci. 9 (3): e0003585. doi:10.1371/journal.pntd.0003585. PMC 4359152. PMID 25768004.
- ^ Cho, B. A.; Cho, N. H.; Seong, S. Y.; Choi, M. S.; Kim, I. S. (2010). "Intracellular invasion by Orientia tsutsugamushi is mediated by integrin signaling and actin cytoskeleton rearrangements". Infekce a imunita. 78 (5): 1915–1923. doi:10.1128/IAI.01316-09. PMC 2863532. PMID 20160019.
- ^ Ko, Y.; Choi, J.H .; Ha, N.Y; Kim, I.S.; Cho, N.H.; Choi, M.S.; Bäumler, A. J. (2013). "Active escape of Orientia tsutsugamushi from cellular autophagy". Infekce a imunita. 81 (2): 552–559. doi:10.1128/IAI.00861-12. PMC 3553808. PMID 23230293.
- ^ Kim, M. J.; Kim, M.K.; Kang, J.S. (2013). "Involvement of lipid rafts in the budding-like exit of Orientia tsutsugamushi". Mikrobiální patogeneze. 63: 37–43. doi:10.1016/j.micpath.2013.06.002. PMID 23791848.
- ^ Bengston, I.A. (1945). "Apparent serological heterogeneity among strains of Tsutsugamushi disease (scrub typhus)". Zprávy o veřejném zdraví. 60 (50): 1483–1488. doi:10.2307/4585496. JSTOR 4585496. PMID 21004496.
- ^ Bengston, I.A. (1946). "A serological study of 37 cases of tsutsugamushi disease (scrub typhus) occurring in Burma and the Philippine Islands". Zprávy o veřejném zdraví. 61 (24): 887–894. doi:10.2307/4585717. JSTOR 4585717. PMID 20987857.
- ^ Shishido, A.; Ohtawara, M.; Tateno, S.; Mizuno, S.; Ogura, M.; Kitaoka, M. (1958). "The nature of immunity against scrub typhus in mice i. the resistance of mice, surviving subcutaneous infection of scrub typhus rickettsia, to intraperitoneal reinfection of the same agent". Japonský žurnál lékařské vědy a biologie. 11 (5): 383–399. doi:10.7883/yoken1952.11.383.
- ^ Jang, M.S.; Neupane, G.P.; Lee, Y.M .; Kim, D.M.; Lee, S.H. (2011). "Phylogenetic analysis of the 56 kDa protein genes of Orientia tsutsugamushi in southwest area of Korea". American Journal of Tropical Medicine and Hygiene. 84 (2): 250–254. doi:10.4269/ajtmh.2011.09-0601. PMC 3029177. PMID 21292894.
- ^ Yamashita, T .; Kasuya, S.; Noda, N.; Nagano, I.; Kang, J.S. (1994). "Přenos Rickettsia tsutsugamushi strains among humans, wild rodents, and trombiculid mites in an area of Japan in which tsutsugamushi disease is newly endemic". Journal of Clinical Microbiology. 32 (11): 2780–2785. doi:10.1128/JCM.32.11.2780-2785.1994. PMC 264159. PMID 7852572.
- ^ A b Ohashi, N .; Koyama, Y .; Urakami, H.; Fukuhara, M.; Tamura, A.; Kawamori, F.; Yamamoto, S .; Kasuya, S.; Yoshimura, K. (1996). "Demonstration of antigenic and genotypic variation in Orientia tsutsugamushi which were isolated in Japan, and their classification into type and subtype". Mikrobiologie a imunologie. 40 (9): 627–638. doi:10.1111/j.1348-0421.1996.tb01120.x. PMID 8908607. S2CID 22112557.
- ^ Bakshi, D.; Singhal, P.; Mahajan, S.K.; Subramaniam, P.; Tuteja, U.; Batra, H.V. (2007). "Development of a real-time PCR assay for the diagnosis of scrub typhus cases in India and evidence of the prevalence of new genotype of O. tsutsugamushi". Acta Tropica. 104 (1): 63–71. doi:10.1016/j.actatropica.2007.07.013. PMID 17870041.
- ^ Parola, P.; Blacksell, S.D.; Phetsouvanh, R.; Phongmany, S.; Rolain, J.M.; Day, N.P.; Newton, P.N.; Raoult, D. (2008). "Genotyping of Orientia tsutsugamushi from humans with scrub typhus, Laos". Vznikající infekční nemoci. 14 (9): 1483–1485. doi:10.3201/eid1409.071259. PMC 2603112. PMID 18760027.
- ^ Tamura, A; Ohashi, N; Urakami, H; Takahashi, K; Oyanagi, M (1985). "Analysis of polypeptide composition and antigenic components of Rickettsia tsutsugamushi by polyacrylamide gel electrophoresis and immunoblotting". Infekce a imunita. 48 (3): 671–675. doi:10.1128/IAI.48.3.671-675.1985. PMC 261225. PMID 3922893.
- ^ Stover, CK; Marana, DP; Carter, JM; Roe, BA; Mardis, E; Oaks, EV (1990). "The 56-kilodalton major protein antigen of Rickettsia tsutsugamushi: molecular cloning and sequence analysis of the sta56 gene and precise identification of a strain-specific epitope". Infekce a imunita. 58 (7): 2076–2084. doi:10.1128/IAI.58.7.2076-2084.1990. PMC 258779. PMID 1694818.
- ^ Stover, C.K.; Marana, D.P.; Dasch, G.A.; Oaks, E.V. (1990). "Molecular cloning and sequence analysis of the Sta58 major antigen gene of Rickettsia tsutsugamushi: sequence homology and antigenic comparison of Sta58 to the 60-kilodalton family of stress proteins". Infekce a imunita. 58 (5): 1360–1368. doi:10.1128/IAI.58.5.1360-1368.1990. PMC 258633. PMID 2108930.
- ^ Arai, S .; Tabara, K.; Yamamoto, N .; Fujita, H.; Itagaki, A.; Kon, M.; Satoh, H.; Araki, K.; Tanaka-Taya, K.; Takada, N.; Yoshikawa, Y.; Ishihara, C.; Okabe, N .; Oishi, K. (2013). "Molecular phylogenetic analysis of Orientia tsutsugamushi založeno na groES a groEL genes". Vector Borne and Zoonotic Diseases. 13 (11): 825–829. doi:10.1089/vbz.2012.1155. PMC 3822374. PMID 24107204.
- ^ A b C Peter, J.V.; Sudarsan, T.I.; Prakash, J.A.J.; Varghese, G.M. (2015). "Severe scrub typhus infection: Clinical features, diagnostic challenges and management". World Journal of Critical Care Medicine. 4 (3): 244–250. doi:10.5492/wjccm.v4.i3.244. PMC 4524821. PMID 26261776.
- ^ Taylor, A.J.; Paris, D.H.; Newton, P.N.; Walker, D.H. (2015). "A systematic review of mortality from untreated scrub typhus (Orientia tsutsugamushi)". PLOS opomíjené tropické nemoci. 9 (8): e0003971. doi:10.1371/journal.pntd.0003971. PMC 4537241. PMID 26274584.
- ^ A b C Bonell, A.; Lubell, Y.; Newton, P.N.; Crump, J.A.; Paris, D.H. (2017). "Estimating the burden of scrub typhus: A systematic review". PLOS opomíjené tropické nemoci. 11 (9): e0005838. doi:10.1371/journal.pntd.0005838. PMC 5634655. PMID 28945755.
- ^ A b Rajapakse, S.; Weeratunga, P.; Sivayoganathan, S.; Fernando, S.D. (2017). "Clinical manifestations of scrub typhus". Transakce Královské společnosti pro tropickou medicínu a hygienu. 111 (2): 43–54. doi:10.1093/trstmh/trx017. PMID 28449088. S2CID 3751821.
- ^ Mahajan, S.K.; Mahajan, S.K. (2017). "Neuropsychiatric manifestations of scrub typhus". Journal of Neurosciences in Rural Practice. 8 (3): 421–426. doi:10.4103/jnrp.jnrp_44_17. PMC 5488565. PMID 28694624.
- ^ Jain, P .; Prakash, S .; Tripathi, P.K.; Chauhan, A .; Gupta, S .; Sharma, U.; Jaiswal, A.K.; Sharma, D.; Jain, A. (2018). „Vznik Orientia tsutsugamushi as an important cause of acute encephalitis syndrome in India". PLOS opomíjené tropické nemoci. 12 (3): e0006346. doi:10.1371/journal.pntd.0006346. PMC 5891077. PMID 29590177.
- ^ WHO (1999). "WHO Recommended Surveillance Standards (Second edition)". WHO/CDS/CSR/ISR/99.2. Světová zdravotnická organizace, Ženeva. p. 124.
- ^ Jiang, J .; Richards, A.L. (2018). "Scrub typhus: No longer restricted to the Tsutsugamushi Triangle". Tropická medicína a infekční choroby. 3 (1): 11. doi:10.3390/tropicalmed3010011. PMC 6136623. PMID 30274409.
- ^ Walker, D.H.; Paris, D.H.; Day, N.P.; Shelite, T.R. (2013). "Unresolved problems related to scrub typhus: A seriously neglected life-threatening disease". American Journal of Tropical Medicine and Hygiene. 89 (2): 301–307. doi:10.4269/ajtmh.13-0064. PMC 3741252. PMID 23926142.
- ^ A b C d González, C. (January 29, 2020). "Una bacteria presente en ácaros causa un raro tipo de infección en el sur". El Mercurio (ve španělštině). p. A9.
- ^ Mørch, K.; Manoharan, A.; Chandy, S.; Chacko, N.; Alvarez-Uria, G.; Patil, S .; Henry, A .; Nesaraj, J.; Kuriakose, C.; Singh, A .; Kurian, S.; Gill Haanshuus, C.; Langeland, N .; Blomberg, B.; Vasanthan Antony, G.; Mathai, D. (2017). "Acute undifferentiated fever in India: a multicentre study of aetiology and diagnostic accuracy". Infekční nemoci BMC. 17 (1): 665. doi:10.1186/s12879-017-2764-3. PMC 5628453. PMID 28978319.
- ^ Wangrangsimakul, T.; Althaus, T.; Mukaka, M.; Kantipong, P.; Wuthiekanun, V.; Chierakul, W.; Blacksell, S.D.; Day, N.P.; Laongnualpanich, A.; Paris, D.H. (2018). "Causes of acute undifferentiated fever and the utility of biomarkers in Chiangrai, northern Thailand". PLOS opomíjené tropické nemoci. 12 (5): e0006477. doi:10.1371/journal.pntd.0006477. PMC 5978881. PMID 29852003.
- ^ Kundavaram, A.P.; Jonathan, A.J.; Nathaniel, S.D.; Varghese, G.M. (2013). "Eschar in scrub typhus: a valuable clue to the diagnosis". Journal of Postgraduate Medicine. 59 (3): 177–178. doi:10.4103/0022-3859.118033. PMID 24029193.
- ^ Varghese, G.M.; Janardhanan, J.; Trowbridge, P.; Peter, J.V.; Prakash, J.A.; Sathyendra, S.; Thomas, K .; David, T.S.; Kavitha, M.L.; Abraham, O.C.; Mathai, D. (2013). "Scrub typhus in South India: clinical and laboratory manifestations, genetic variability, and outcome". International Journal of Infection Diseases. 17 (11): e981-987. doi:10.1016/j.ijid.2013.05.017. PMID 23891643.
- ^ Kelman, P.; Thompson, C.W.; Hynes, W.; Bergman, C.; Lenahan, C.; Brenner, J.S.; Brenner, M.G.; Goodman, B.; Borges, D.; Filak, M.; Gaff, H. (2018). "Rickettsia parkeri infections diagnosed by eschar biopsy, Virginia, USA". Infekce. 46 (4): 559–563. doi:10.1007/s15010-018-1120-x. PMID 29383651. S2CID 30442009.
- ^ Silva, N.; Eremeeva, M.E.; Rozental, T.; Ribeiro, G.S.; Paddock, C.D.; Ramos, E.A.; Favacho, A.R.; Reis, M.G.; Dasch, G.A.; de Lemos, E.R.; Ko, A.I. (2011). "Eschar-associated spotted fever rickettsiosis, Bahia, Brazil". Vznikající infekční nemoci. 17 (2): 275–278. doi:10.3201/eid1702.100859. PMC 3204763. PMID 21291605.
- ^ Hulmani, M.; Alekya, P.; Kumar, V.J. (2017). "Indian tick typhus presenting as purpura fulminans with review on rickettsial infections". Indian Journal of Dermatology. 62 (1): 1–6. doi:10.4103/0019-5154.198030. PMC 5286740. PMID 28216718.
- ^ Walker, D.H. (1989). "Rickettsioses of the spotted fever group around the world". The Journal of Dermatology. 16 (3): 169–177. doi:10.1111/j.1346-8138.1989.tb01244.x. PMID 2677080. S2CID 21977403.
- ^ Denison, A.M.; Amin, B.D.; Nicholson, W.L.; Paddock, C.D. (2014). "Detekce Rickettsia rickettsii, Rickettsia parkeri, a Rickettsia akari in skin biopsy specimens using a multiplex real-time polymerase chain reaction assay". Klinické infekční nemoci. 59 (5): 635–642. doi:10.1093/cid/ciu358. PMC 4568984. PMID 24829214.
- ^ Le Viet, N.; Laroche, M .; Thi Pham, H.L.; Viet, N.L.; Mediannikov, O.; Raoult, D .; Parola, P. (2017). "Use of eschar swabbing for the molecular diagnosis and genotyping of Orientia tsutsugamushi causing scrub typhus in Quang Nam province, Vietnam". PLOS opomíjené tropické nemoci. 11 (2): e0005397. doi:10.1371/journal.pntd.0005397. PMC 5344524. PMID 28241043.
- ^ Koraluru, M.; Bairy, I.; Varma, M.; Vidyasagar, S. (2015). "Diagnostic validation of selected serological tests for detecting scrub typhus". Mikrobiologie a imunologie. 59 (7): 371–374. doi:10.1111/1348-0421.12268. PMID 26011315. S2CID 49921.
- ^ Patricia, K.A.; Hoti, S.L.; Kanungo, R.; Jambulingam, P.; Shashikala, N.; Naik, A.C. (2017). "Improving the diagnosis of scrub typhus by combining groEL-based polymerase chain reaction and IgM ELISA". Journal of Clinical and Diagnostic Research. 11 (8): DC27–DC31. doi:10.7860/JCDR/2017/26523.10519. PMC 5620764. PMID 28969124.
- ^ Rahi, M.; Gupte, M.D.; Bhargava, A .; Varghese, G.Mm; Arora, R. (2015). "DHR-ICMR Guidelines for diagnosis & management of rickettsial diseases in India". Indian Journal of Medical Research. 141 (4): 417–22. doi:10.4103/0971-5916.159279. PMC 4510721. PMID 26112842.
- ^ Chanta, C.; Phloenchaiwanit, P. (2015). "Randomized Controlled trial of azithromycin versus doxycycline or chloramphenicol for treatment of uncomplicated pediatric scrub typhus". Journal of the Medical Association of Thailand. 98 (8): 756–760. PMID 26437532.
- ^ Lee, S.C.; Cheng, Y.J.; Lin, C.H.; Lei, W.T.; Chang, H.Y.; Lee, M.D.; Liu, J.M.; Hsu, R.J.; Chiu, N.C.; Chi, H.; Peng, C.C.; Tsai, T.L.; Lin, C.Y. (2017). "Comparative effectiveness of azithromycin for treating scrub typhus". Lék. 96 (36): e7992. doi:10.1097/MD.0000000000007992. PMC 6392745. PMID 28885357.
- ^ Card, W.I.; Walker, J.M. (1947). "Scrub-typhus vaccine; field trial in South-east Asia". Lanceta. 1 (6450): 481–483. doi:10.1016/S0140-6736(47)91989-2. PMID 20294827.
- ^ Berge, T.O.; Gauld, R.L.; Kitaoka, M. (1949). "A field trial of a vaccine prepared from the Volner strain of Rickettsia tsutsugamushi". American Journal of Hygiene. 50 (3): 337–342. doi:10.1093/oxfordjournals.aje.a119366. PMID 15391985.
- ^ A b Valbuena, G.; Walker, D. H. (2013). "Approaches to vaccines against Orientia tsutsugamushi". Hranice v buněčné a infekční mikrobiologii. 2: 127. doi:10.3389/fcimb.2012.00170. PMC 3539663. PMID 23316486.
- ^ Choi, S .; Jeong, H.J.; Hwang, K.J.; Gill, B.; Ju, Y.R.; Lee, Y.S.; Lee, J. (2017). "A recombinant 47-kDa outer membrane protein induces an immune response against Orientia tsutsugamushi strain Boryong". American Journal of Tropical Medicine and Hygiene. 97 (1): 30–37. doi:10.4269/ajtmh.15-0771. PMC 5508880. PMID 28719308.
- ^ Smadel, JE; Ley, H.L.Jr.; Diercks, F.H.; Traub, R. (1950). "Immunity in scrub typhus: resistance to induced reinfection". A.M.A. Archives of Pathology. 50 (6): 847–861. PMID 14789327.
- ^ MacMillan, J.G.; Rice, R.M.; Jerrells, T.R. (1985). "Development of antigen-specific cell-mediated immune responses after infection of cynomolgus monkeys (Macaca fascicularis) s Rickettsia tsutsugamushi". The Journal of Infectious Diseases. 152 (4): 739–749. doi:10.1093/infdis/152.4.739. PMID 2413138.
- ^ Ha, N.Y.; Kim, Y .; Min, C.K.; Kim, H.I.; Yen, N.T.H.; Choi, M.S.; Kang, J.S .; Kim, Y.S .; Cho, N.H. (2017). "Longevity of antibody and T-cell responses against outer membrane antigens of Orientia tsutsugamushi in scrub typhus patients". Vznikající mikroby a infekce. 6 (12): e116. doi:10.1038/emi.2017.106. PMC 5750460. PMID 29259327.