EZH2 - EZH2




Vylepšení homologu zeste 2 (EZH2) je a histon-lysin N-methyltransferáza enzym (ES 2.1.1.43 ) kódováno EZH2 gen, který se účastní methylace histonu a nakonec transkripční represe.[5] EZH2 katalyzuje přidání methylové skupiny na histon H3 na lysin 27,[6] pomocí kofaktor S-adenosyl-L-methionin. Methylační aktivita EZH2 usnadňuje heterochromatin tvorba tak umlčí genovou funkci.[5] Přestavba chromozomů heterochromatin EZH2 je také nutný během buněčné mitózy.
EZH2 je funkční enzymatická složka Polycomb represivní komplex 2 (PRC2 ), který je zodpovědný za zdravé embryonální vývoj skrz epigenetický udržování genů odpovědných za regulaci vývoje a diferenciace.[7] EZH2 je zodpovědný za metylační aktivitu PRC2 a komplex také obsahuje proteiny potřebné pro optimální funkci (EED, SUZ12, JARID2, AEBP2, RbAp46 / 48, a PCL ).[8]
Mutace nebo nadměrná exprese EZH2 byla spojena s mnoha formami rakoviny.[9] EZH2 inhibuje geny odpovědné za potlačení vývoje nádoru a blokování aktivity EZH2 může zpomalit růst nádoru. EZH2 byl zaměřen na inhibici, protože je upregulován u více druhů rakoviny včetně, ale bez omezení na prsa,[10] prostata,[11] melanom,[12] a rakovina močového měchýře.[13] S mutacemi v genu EZH2 jsou také spojeny Weaverův syndrom, vzácný kongenitální porucha,[14] a EZH2 se podílí na vzniku neurodegenerativní příznaky poruchy nervového systému, ataxie telangiektázie.[15]
Funkce
| Histon-lysin N-methyltransferáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 2.1.1.43 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
EZH2 je katalytický podjednotka Polycomb represivního komplexu 2 (PRC2).[16] Katalytická aktivita EZH2 závisí na tvorbě komplexu s alespoň dvěma dalšími složkami PRC2, SUZ12 a EED.[17]
Jako histon methyltransferáza (HMTase), primární funkcí EZH2 je methylát Lys-27 zapnuto histon 3 (H3K27me) přenosem methylové skupiny z kofaktoru S-adenosyl-L-methioninu (SAM). EZH2 je schopen mono-, di- a tri-methylace H3K27 a byla spojována s řadou biologických funkcí, včetně transkripční regulace v krvetvorba, vývoj a diferenciace buněk.[17][18][19][20]
Nedávné studie ukázaly, že EZH2 je také schopen methylovathiston bílkoviny.[17][18]
Transkripční represe
EZH2 jako součást PRC2 katalyzuje trimethylaci H3K27 (H3K27me3 ), což je histonová modifikace, která byla charakterizována jako součást histonový kód.[16][20][21][22] Histonový kód je teorie, že chemické modifikace, jako např methylace, acetylace, a ubikvitinace, z histonových proteinů hraje v epigenetický regulace gen transkripce. EZH2 zprostředkovaná katalýza H3K27me3 je spojena s dlouhodobou transkripční represí.[16][20][21]
EZH2, stejně jako další proteiny skupiny Polycomb, se podílejí na vytváření a udržování genové represe prostřednictvím buněčné dělení.[17][20] Předpokládá se, že tento transkripčně represivní stav je způsoben methylací H3K27 zprostředkovanou PRC2 / EZH2-EED a následným získáváním PRC1 což usnadňuje kondenzaci chromatin a formování heterochromatin.[20] Heterochromatin je pevně zabalený chromatin, který omezuje přístup transkripčního aparátu k podkladové DNA, čímž potlačuje transkripci.[23]
Během buněčného dělení je tvorba heterochromatinu nezbytná segregace chromozomů.[24] Komplex PRC2 / EED-EZH2 může být také zapojen do náboru DNA methyltransferáz (DNMT ), což má za následek zvýšení Methylace DNA, další epigenetická vrstva transkripční represe.[16][17] Mezi specifické geny, které byly identifikovány jako cíle transkripční represe zprostředkované EZH2, patří HOXA9, HOXC8, MYT1, CDKN2A a kyselina retinová cílové geny.[16]
Aktivace přepisu
U rakoviny může EZH2 hrát roli v aktivaci transkripce, nezávisle na PRC2.[17] Bylo prokázáno, že v buňkách rakoviny prsu se EZH2 aktivuje NF-kB cílové geny, které se účastní odpovědí na podněty.[17] Funkční role této činnosti a její mechanismus jsou stále neznámé.
Vývoj a diferenciace buněk
EZH2 hraje při vývoji zásadní roli. Zejména pomáhá kontrolovat transkripční represi genů, které regulují buněčnou diferenciaci.[17][18][20][21] V embryonálních kmenových buňkách se EZH2 zprostředkovaná trimethylace H3K27me3 v oblastech obsahujících vývojové geny jeví jako důležitá pro udržení normální buněčné diferenciace.[20] H3K27me3 je také důležitý při řízení X-inaktivace, umlčení jednoho X-chromozom u žen během vývoje.[22] Během X-inaktivace se má za to, že EZH2 se podílí na zahájení tvorby heterochromatinu trimethylací H3K27 a že další histonmethyltransferázy a histonové značky se mohou podílet na udržování umlčeného stavu.[25]
Dále byl identifikován EZH2 jako esenciální protein podílející se na vývoji a diferenciaci B-buňky a T-buňky.[18] H3K27me3 se podílí na potlačení genů, které podporují diferenciaci, čímž udržuje nediferencovaný stav B- a T-buněk a hraje důležitou roli při regulaci krvetvorba.[18]
Regulace aktivity EZH2
Aktivita EZH2 je regulována posttranslační fosforylace z threonin a serin zbytky na EZH2.[26] Konkrétně fosforylace T350 byla spojena se zvýšením aktivity EZH2, zatímco fosforylace T492 a S21 byla spojena se snížením aktivity EZH2.[21][26] Bylo navrženo, že fosforylace T492 narušuje kontakty mezi lidským EZH2 a jeho vazebnými partnery v komplexu PRC2, čímž brání jeho katalytické aktivitě.[21]
Kromě fosforylace se také ukázalo, že aktivita PRC2 / EZH2-EED je antagonizována transkripčně aktivujícími histonovými značkami, jako je acetylace z H3K27 (H3K27ac ) a methylace H3K36 (H3K36me ).[21][27]
Enzymatická aktivita
Funkce EZH2 je vysoce závislá na jejím náboru komplexem PRC2. Zejména, Vývoj embryonálního ektodermu WD40-opakovaný protein (EED) a supresor bílkovin se zinkovým prstem zeste 12 (SUZ12) jsou potřebné ke stabilizaci interakce EZH2 s jeho histonovým substrátem[28][29] Nedávno byly generovány dvě izoformy EZH2 alternativní sestřih byly identifikovány u lidí: EZH2α a EZH2β.[30] Oba izoformy obsahují prvky, které byly označeny jako důležité pro funkci EZH2, včetně signál jaderné lokalizace, vazebná místa EED a SUZ12 stejně jako konzervovaná doména SET.[30] Většina studií se dosud zaměřila na delší izoformu EZH2α, ale EZH2β, která postrádá exony 4 a 8, je aktivní.[30] Dále komplexy PRC2 / EZH2p působí na odlišné geny od genů jeho protějšku PRC2 / EZH2α, což naznačuje, že každá izoforma může působit k regulaci specifické podskupiny genů.[30] Další důkazy naznačují, že EZH2 může být také schopen metylace lysinu nezávisle na asociaci s PRC2, když je EZH2 vysoce upregulován.[17]
Methylace lysinu

Methylace je přidání -CH3nebo methylová skupina na jinou molekulu. V biologii je methylace typicky katalyzována enzymy a methylové skupiny se běžně přidávají buď k proteinům, nebo k nukleovým kyselinám. Při methylaci katalyzované EZH2 jde o aminokyselinu lysin v histonu je h3 methylován. Tento aminokyselinový zbytek může být na svém konci methylován až třikrát amonný skupina. Tyto methylované lysiny jsou důležité při kontrole exprese genů savců a mají funkční roli v heterochromatin formace, Inaktivace X-chromozomu a transkripční regulace.[31] V savčích chromozomech může methylace histonového lysinu buď aktivovat nebo potlačovat geny v závislosti na místě methylace. Nedávné práce ukázaly, že alespoň částí funkce tlumení komplexu EZH2 je methylace histon H3 na lysinu 27.[32] Methylace a další modifikace probíhají na histonech. Methylové modifikace mohou ovlivnit vazbu proteinů na tyto histony a buď aktivovat, nebo inhibovat transkripce.[24]
Mechanismus katalýzy

EZH2 je členem SET doména rodina lysinu methyltransferázy které fungují pro přidání methylových skupin do lysinových postranních řetězců Podklad bílkoviny.[33] SET methyltransferázy závisí na kofaktoru S-adenosylmethioninu (SAM), který pro svou katalytickou aktivitu působí jako donor methylu. Proteiny domény SET se liší od ostatních SAM závislých methyltransferázy v tom, že vážou svůj substrát a SAM kofaktor na opačných stranách Aktivní stránky enzymu. Tato orientace substrátu a kofaktoru umožňuje SAM disociovat bez narušení vazby substrátu a může vést k několika cyklům methylace lysinu bez disociace substrátu.[33]
Ačkoli pro EZH2 nebyla stanovena ani krystalová struktura vázaná na substrát nebo SAM, vyrovnání struktury STAMP s lidskou SET7 / 9 methyltransferázou ukazuje konzervované tyrosin zbytky v téměř identických pozicích v domnělém aktivním místě EZH2.

Dříve se předpokládalo, že tyrosin 726 v aktivním místě EZH2 působí jako obecná báze k deprotonaci substrátu lysinu, ale kinetické izotopové účinky ukázaly, že zbytky aktivního místa nejsou přímo zapojeny do chemie reakce methyltransferázy.[34] Místo toho tyto experimenty podporují mechanismus, ve kterém zbytky snižují pKa substrátu lysinového zbytku a současně poskytuje kanál pro vodu pro přístup k postrannímu řetězci lysinu uvnitř aktivního místa. Velkoobjemová rozpouštědlová voda pak může snadno deprotonovat lysinový postranní řetězec a aktivuje jej pro nukleofilní útok kofaktoru SAM v SN2 reakce, která vede k přenosu methylové skupiny ze SAM do postranního řetězce lysinu.[34]

EZH2 primárně katalyzuje mono- a di-methylaci H3K27, ale klinicky relevantní mutaci zbytkového tyrosinu 641 na fenylalanin (Y641F) vede k vyšší tri-methylační aktivitě H3K27.[34] Předpokládá se, že odstranění hydroxylové skupiny na Y641 ruší sterické zábrany a umožňuje umístění třetí methylové skupiny na substrátu lysinu.
Klinický význam
Rakovina
EZH2 je atraktivní cíl pro protinádorovou terapii, protože pomáhá dělení a množení rakovinných buněk. Vyskytuje se ve větším množství než ve zdravých buňkách u široké škály druhů rakoviny včetně prsu, prostaty, močového měchýře, děložní, a ledvin rakoviny, stejně jako melanom a lymfom. EZH2 je a gen supresor, takže když dojde k nadměrné expresi, vypne se mnoho genů potlačujících nádor, které jsou normálně zapnuté. Inhibice funkce EZH2 se zmenšuje zhoubný nádory v některých hlášených případech, protože tyto tumor supresorové geny nejsou umlčeny EZH2.[35] EZH2 se obvykle nevyjadřuje u zdravých dospělých; nachází se pouze v aktivně se dělících buňkách, jako jsou ty, které jsou aktivní během vývoje plodu.[36] Kvůli této vlastnosti lze nadexpresi EZH2 použít jako diagnostický marker rakoviny a některých neurodegenerativních poruch.[15] Existují však případy, kdy je obtížné určit, zda je nadměrná exprese EZH2 příčinou onemocnění nebo jen následkem. Pokud je to jen důsledek, cílení na EZH2 pro inhibici nemusí nemoc vyléčit. Jedním příkladem dráhy rakoviny, ve které hraje roli EZH2, je cesta pRB-E2F. Je po proudu od pRB-E2F dráha a signály z této dráhy vedou k nadměrné expresi EZH2.[37] Další důležitou charakteristikou EZH2 je, že když je EZH2 nadměrně exprimován, může aktivovat geny bez tvorby PRC2. To je problém, protože to znamená, že methylační aktivita enzymu není zprostředkována tvorbou komplexu. V buňkách rakoviny prsu EZH2 aktivuje geny, které podporují buněčnou proliferaci a přežití.[17] Může také aktivovat regulační geny jako c-myc a cyklin D1 interakcí s Wnt signalizace faktory.[38] Důležité je, že mutace tyrosinu 641 v aktivní doméně SET na řadu různých aminokyselin je společným rysem některých B-buněčných lymfomů.[39]

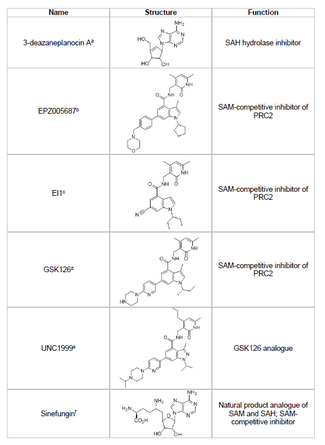
Inhibitory
Vývoj inhibitoru EZH2 a prevence nežádoucí methylace histonových supresorových genů je životaschopnou oblastí výzkumu rakoviny. Vývoj inhibitoru EZH2 se zaměřil na cílení na SET doména aktivní místo proteinu. Od roku 2015 bylo vyvinuto několik inhibitorů EZH2, včetně 3-deazaneplanocin A (DZNep), EPZ005687, EI1, GSK126 a UNC1999. DZNep má potenciální antivirové a protirakovinné vlastnosti, protože snižuje hladiny EZH2 a indukuje apoptóza v buňkách rakoviny prsu a tlustého střeva.[40] DZNep inhibuje hydrolýzu S-adenosyl-L-homocysteinu (SAH), který je produktovým inhibitorem všech proteinových methyltransferáz, což vede ke zvýšení buněčných koncentrací SAH, které zase inhibují EZH2. DZNep však není specifický pro EZH2 a také inhibuje další DNA methyltransferázy.
V roce 2012 společnost s názvem Epizyme odhalila EPZ005687, S-adenosylmethionin (SAM ) kompetitivní inhibitor, který je selektivnější než DZNep; ve srovnání s má 50krát vyšší selektivitu pro EZH2 EZH1. Lék blokuje aktivitu EZH2 vazbou na aktivní místo enzymu v doméně SET. EPZ005687 může také inhibovat Y641 a A677 mutanty EZH2, které mohou být použitelné pro léčbu non-Hodgkinova lymfomu.[41] V roce 2013 zahájil Epizyme klinické studie fáze I s dalším inhibitorem EZH2, tazemetostat (EPZ-6438), pro pacienty s B-buněčným lymfomem.[45] V roce 2020 tazemetostat, s obchodním názvem Tazverik, byl schválen FDA pro léčbu metastatického nebo lokálně pokročilého epitelioidního sarkomu a byl schválen pro léčbu pacientů s relabujícím folikulárním lymfomem později v tomto roce.[46]
Sinefungin je dalším SAM kompetitivním inhibitorem, ale podobně jako DZNep není specifický pro EZH2.[44] Funguje tak, že se váže do kapsy vázající kofaktor DNA methyltransferáz a blokuje přenos methylu. EI1 je další inhibitor vyvinutý společností Novartis, která prokázala EZH2 inhibiční aktivitu v nádorových buňkách lymfomu, včetně buněk s mutací Y641.[42] Mechanismus tohoto inhibitoru také zahrnuje kompetici s kofaktorem SAM o vazbu na EZH2.[42] GSK126 je silný, SAM kompetitivní inhibitor EZH2 vyvinutý společností GlaxoSmithKline, který má 150násobnou selektivitu oproti EZH1 a K.i 0,5 až 3 nM.[43] UNC1999 byl vyvinut jako analog GSK126 a byl prvním orálně biologicky dostupným inhibitorem EZH2, který vykazoval aktivitu. Je však méně selektivní než jeho protějšek GSK126 a váže se také na EZH1, což zvyšuje potenciál pro nežádoucí účinky.
Kombinované terapie jsou studovány jako možné léčby, když primární léčba začíná selhávat. Etoposid, a topoizomeráza Inhibitor v kombinaci s inhibitorem EZH2 se stává účinnějším pro nemalobuněčné rakoviny plic s BRG1 a EGFR mutace.[35] Methylace EZH2 a lysinu však může mít aktivitu potlačující nádor, například v myelodysplastický syndrom,[47] což naznačuje, že inhibice EZH2 nemusí být ve všech případech prospěšná.
Weaverův syndrom
Byly spojeny mutace v genu EZH2 Weaverův syndrom, vzácná porucha charakterizovaná pokročilým kostním věkem, makrocefalie, a hypertelorismus.[14] The histidin rezidua v aktivním místě divoký typ EZH2 byl mutován na tyrosin u pacientů s diagnostikovaným Weaverovým syndromem.[14] Mutace pravděpodobně interferuje kofaktor vazba a způsobuje narušení přirozené funkce proteinu.[14]
Taxonomické rozdělení

Enhancer of zeste (E (z)) byl původně identifikován v Drosophila melanogaster a jeho savčí homology byly následně identifikovány a pojmenovány EZH1 (enhancer zeste homolog 1) a EZH2 (enhancer zeste homolog 2).[49] EZH2 je vysoce konzervovaný vývoj. On a jeho homology hrají zásadní roli ve vývoji, buněčné diferenciaci a dělení buněk v rostlinách, hmyzu, rybách a savcích.[17][21][50][51] Následující taxonomický strom je znázorněním distribuce EZH2 v celé řadě druhů.[52][53]
Viz také
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000106462 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání Ensembl 89: ENSMUSG00000029687 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b Viré E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C a kol. (2006). "Protein skupiny Polycomb EZH2 přímo řídí methylaci DNA". Příroda. 439 (7078): 871–4. doi:10.1038 / nature04431. PMID 16357870. S2CID 4409726.
- ^ Cao R, Wang L, Wang H, Xia L, Erdjument-Bromage H, Tempst P, Jones RS, Zhang Y (2002). "Role methylace histonu H3 lysinu 27 v umlčování skupiny Polycomb". Věda. 298 (5595): 1039–43. doi:10.1126 / science.1076997. PMID 12351676. S2CID 6265267.
- ^ Morey L, Helin K (2010). "Represe transkripce zprostředkovaná proteinem skupiny Polycomb.". Trends Biochem. Sci. 35 (6): 323–32. doi:10.1016 / j.tibs.2010.02.009. PMID 20346678.
- ^ Margueron R, Reinberg D (2011). „Polycomb komplex PRC2 a jeho značka v životě“. Příroda. 469 (7330): 343–9. doi:10.1038 / nature09784. PMC 3760771. PMID 21248841.
- ^ Kim, Kimberly; Roberts, Charles (18. prosince 2015). „Cílení na EZH2 u rakoviny“. Přírodní medicína. 22 (2): 128–134. doi:10,1038 / nm.4036. PMC 4918227. PMID 26845405.
- ^ Yoo KH, Hennighausen L (2012). „EZH2 methyltransferáza a methylace H3K27 u rakoviny prsu“. Int. J. Biol. Sci. 8 (1): 59–65. doi:10,7150 / ijbs.8,59. PMC 3226033. PMID 22211105.
- ^ Varambally S, Dhanasekaran SM, Zhou M, Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt RG, Otte AP, Rubin MA, Chinnaiyan AM (2002). „Polycombový protein skupiny EZH2 se podílí na progresi rakoviny prostaty“ (PDF). Příroda. 419 (6907): 624–9. doi:10.1038 / nature01075. hdl:2027.42/62896. PMID 12374981. S2CID 4414767. Shrnutí ležel – Scientific American.
- ^ Zingg D, Debbache J, Schaefer SM, Tuncer E, Frommel SC, Cheng P a kol. (2015). „Epigenetický modifikátor EZH2 řídí růst melanomu a metastázy prostřednictvím umlčování odlišných nádorových supresorů“. Nat Commun. 6: 6051. doi:10.1038 / ncomms7051. PMID 25609585. Shrnutí ležel – MedicalResearch.com.
- ^ Arisan S, Buyuktuncer ED, Palavan-Unsal N, Caşkurlu T, Cakir OO, Ergenekon E (2005). "Zvýšená exprese EZH2, proteinu skupiny polycombů, v karcinomu močového měchýře". Urol. Int. 75 (3): 252–7. doi:10.1159/000087804. PMID 16215315. S2CID 26843362.
- ^ A b C d Gibson WT, Hood RL, Zhan SH, Bulman DE, Fejes AP, Moore R a kol. (2012). „Mutace v EZH2 způsobují Weaverův syndrom“. Dopoledne. J. Hum. Genet. 90 (1): 110–8. doi:10.1016 / j.ajhg.2011.11.018. PMC 3257956. PMID 22177091.
- ^ A b Li J, Hart RP, Mallimo EM, Swerdel MR, Kusnecov AW, Herrup K (2013). „Trimethylace H3K27 zprostředkovaná EZH2 zprostředkovává neurodegeneraci v ataxii-telangiektázii“. Nat. Neurosci. 16 (12): 1745–53. doi:10.1038 / č. 3564. PMC 3965909. PMID 24162653.
- ^ A b C d E Univerzální přístupové číslo zdroje bílkovin Q15910 na UniProt.
- ^ A b C d E F G h i j k Tan JZ, Yan Y, Wang XX, Jiang Y, Xu HE (2014). „EZH2: biologie, nemoci a objevování léků na základě struktury“. Acta Pharmacol. Hřích. 35 (2): 161–74. doi:10.1038 / aps.2013.161. PMC 3914023. PMID 24362326.
- ^ A b C d E Lund K, Adams PD, Copland M (2014). „EZH2 u normální a maligní krvetvorby“. Leukémie. 28 (1): 44–9. doi:10.1038 / leu.2013.288. PMID 24097338. S2CID 736796.
- ^ „RefSeq“. RefSeq Gene EZH2. Citováno 1. února 2015.
- ^ A b C d E F G Ding X, Wang X, Sontag S, Qin J, Wanek P, Lin Q, Zenke M (2014). „Polycombový protein Ezh2 má dopad na indukovanou generaci pluripotentní kmenové buňky“. Kmenové buňky Dev. 23 (9): 931–40. doi:10.1089 / scd.2013.0267. PMC 3996971. PMID 24325319.
- ^ A b C d E F G O'Meara MM, Simon JA (2012). „Vnitřní fungování a regulační vstupy, které řídí Polycomb represivní komplex 2“. Chromozom. 121 (3): 221–34. doi:10.1007 / s00412-012-0361-1. PMC 3351537. PMID 22349693.
- ^ A b „Histone H3K27“. EpiGenie.
- ^ Grewal SI, Jia S (2007). "Heterochromatin se vrátil". Nat. Genet. 8 (1): 35–46. doi:10.1038 / nrg2008. PMID 17173056. S2CID 31811880.
- ^ A b Stewart MD, Li J, Wong J (2005). „Vztah mezi methylací histonu H3 lysinu 9, transkripční represí a náborem heterochromatinového proteinu 1“. Mol. Buňka. Biol. 25 (7): 2525–2538. doi:10.1128 / MCB.25.7.2525-2538.2005. PMC 1061631. PMID 15767660.
- ^ Jeanteur, Philippe (2008). Epigenetika a chromatin. Springer. ISBN 9783540852360.
- ^ A b Kaneko S, Li G, Son J, Xu CF, Margueron R, Neubert TA, Reinberg D (2010). „Fosforylace PRC2 složky Ezh2 je regulována buněčným cyklem a reguluje její vazbu na ncRNA“. Genes Dev. 24 (23): 2615–20. doi:10.1101 / gad.1983810. PMC 2994035. PMID 21123648.
- ^ Tie F, Banerjee R, Stratton CA, Prasad-Sinha J, Stepanik V, Zlobin A, Diaz MO, Scacheri PC, Harte PJ (2009). "CBP zprostředkovaná acetylace histonového H3 lysinu 27 antagonizuje tlumení Drosophila Polycomb". Rozvoj. 136 (18): 3131–41. doi:10,1242 / dev.037127. PMC 2730368. PMID 19700617.
- ^ Cao R, Zhang Y (2004). „SUZ12 je vyžadován jak pro aktivitu histonmethyltransferázy, tak pro funkci tlumení komplexu EED-EZH2.“ Mol. Buňka. 15 (1): 57–67. doi:10.1016 / j.molcel.2004.06.020. PMID 15225548.
- ^ Denisenko O, Shnyreva M, Suzuki H, Bomsztyk K (1998). „Bodové mutace v doméně WD40 Eed blokují jeho interakci s Ezh2“. Mol. Buňka. Biol. 18 (10): 5634–42. doi:10.1128 / MCB.18.10.5634. PMC 109149. PMID 9742080.
- ^ A b C d Grzenda A, Lomberk G, Svingen P, Mathison A, Calvo E, Iovanna J, Xiong Y, Faubion W, Urrutia R (28. února 2013). „Funkční charakterizace EZH2β odhaluje zvýšenou složitost izoforem EZH2 zapojených do regulace exprese genů savců“. Epigenetika a chromatin. 6 (1): 3. doi:10.1186/1756-8935-6-3. PMC 3606351. PMID 23448518.
- ^ Martin C, Zhang Y (2005). "Různé funkce methylace histon-lysinu". Nat. Rev. Mol. Cell Biol. 6 (11): 838–849. doi:10.1038 / nrm1761. PMID 16261189. S2CID 31300025.
- ^ Brien GL, Gambero G, O'Connell DJ, Jerman E, Turner SA, Egan CM a kol. (2012). "Polycomb PHF19 váže H3K36me3 a rekrutuje PRC2 a demethylázu NO66 na geny embryonálních kmenových buněk během diferenciace". Nat. Struct. Mol. Biol. 19 (12): 1273–1281. doi:10.1038 / nsmb.2449. PMID 23160351. S2CID 1017805.
- ^ A b Dillon SC, Zhang X, Trievel RC, Cheng X (2005). „Nadrodina proteinů domény SET: proteinové lysinmethyltransferázy“ (PDF). Genome Biol. 6 (8): 227. doi:10.1186 / gb-2005-6-8-227. PMC 1273623. PMID 16086857. Citováno 1. února 2015.
- ^ A b C Kipp DR, Quinn CM, Fortin PD (2013). "Enzymaticky závislá deprotonace lysinu v katalýze EZH2". Biochemie. 52 (39): 6866–78. doi:10.1021 / bi400805w. PMID 24000826.
- ^ A b „Okno zranitelnosti“. Harvardská lékařská škola.
- ^ A b Konze KD, Ma A, Li F, Barsyte-Lovejoy D, Parton T, Macnevin CJ, Liu F, Gao C, Huang XP, Kuznetsova E, Rougie M, Jiang A, Pattenden SG, Norris JL, James LI, Roth BL, Brown PJ, Frye SV, Arrowsmith CH, Hahn KM, Wang GG, Vedadi M, Jin J (2013). „Perorálně biologicky dostupná chemická sonda lysinmethyltransferáz EZH2 a EZH1“. ACS Chem. Biol. 8 (6): 1324–34. doi:10.1021 / cb400133j. PMC 3773059. PMID 23614352.
- ^ Bracken AP, Pasini D, Capra M, Prosperini E, Colli E, Helin K (2003). „EZH2 je po směru dráhy pRB-E2F, nezbytný pro proliferaci a zesílený při rakovině“. EMBO J.. 22 (20): 5323–35. doi:10.1093 / emboj / cdg542. PMC 213796. PMID 14532106.
- ^ Shi B, Liang J, Yang X, Wang Y, Zhao Y, Wu H, Sun L, Zhang Y, Chen Y, Li R, Zhang Y, Hong M, Shang Y (2007). „Integrace signalizačních obvodů estrogenu a Wnt proteinem polycombové skupiny EZH2 v buňkách rakoviny prsu“. Mol. Buňka. Biol. 27 (14): 5105–19. doi:10.1128 / MCB.00162-07. PMC 1951944. PMID 17502350.
- ^ Morin RD, Johnson NA, Severson TM, Mungall AJ, An J, Goya R a kol. (2010). „Somatické mutace měnící EZH2 (Tyr641) ve folikulárních a difúzních velkých B-buněčných lymfomech původu z germinálního centra“. Nat. Genet. 42 (2): 181–5. doi:10,1038 / ng.518. PMC 2850970. PMID 20081860.
- ^ A b Tan J, Yang X, Zhuang L, Jiang X, Chen W, Lee PL, Karuturi RK, Tan PB, Liu ET, Yu Q (2007). „Farmakologické narušení represe genu potlačujícího komplex Polycomb 2 zprostředkované selektivně indukuje apoptózu v rakovinných buňkách“. Genes Dev. 21 (9): 1050–63. doi:10.1101 / gad.1524107. PMC 1855231. PMID 17437993.
- ^ A b Knutson SK, Wigle TJ, Warholic NM, Sneeringer CJ, Allain CJ, Klaus CR a kol. (2012). „Selektivní inhibitor EZH2 blokuje methylaci H3K27 a zabíjí buňky mutantního lymfomu“. Nat. Chem. Biol. 8 (11): 890–6. doi:10.1038 / nchembio.1084. PMID 23023262.
- ^ A b C Qi W, Chan H, Teng L, Li L, Chuai S, Zhang R a kol. (2012). „Selektivní inhibice Ezh2 inhibitorem s malou molekulou blokuje proliferaci nádorových buněk“. Proc. Natl. Acad. Sci. USA. 109 (52): 21360–5. doi:10.1073 / pnas.1210371110. PMC 3535655. PMID 23236167.
- ^ A b McCabe MT, Ott HM, Ganji G, Korenchuk S, Thompson C, Van Aller GS a kol. (2012). „Inhibice EZH2 jako terapeutická strategie pro lymfom s mutacemi aktivujícími EZH2“. Příroda. 492 (7427): 108–12. doi:10.1038 / příroda11606. PMID 23051747. S2CID 4385729.
- ^ A b Couture JF, Hauk G, Thompson MJ, Blackburn GM, Trievel RC (2006). „Katalytické role vodíkové vazby uhlík-kyslík v lysinmethyltransferázách v doméně SET“. J. Biol. Chem. 281 (28): 19280–7. doi:10,1074 / jbc.M602257200. PMID 16682405.
- ^ Společnost Epizyme oznámila klinická data ze studie fáze 1 s inhibitorem EZH2 EPZ-6438 (E7438), která bude uvedena na sympoziu EORTC-NCI-AACR. (2014, 1. října).
- ^ https://www.fda.gov/drugs/fda-granted-accelerated-approval-tazemetostat-follicular-lymphoma
- ^ Nikoloski G, Langemeijer SM, Kuiper RP, Knops R, Massop M, Tönnissen ER a kol. (2010). „Somatické mutace genu histonmethyltransferázy EZH2 u myelodysplastických syndromů“. Nat. Genet. 42 (8): 665–7. doi:10,1038 / ng.620. PMID 20601954. S2CID 5814891.
- ^ "Soubor". Genový strom EZH2. Citováno 19. února 2015.
- ^ Laible G, Wolf A, Dorn R, Reuter G, Nislow C, Lebersorger A, Popkin D, Pillus L, Jenuwein T (1997). "Savčí homology genu Polycomb-group Enhancer of zeste zprostředkovávají umlčení genů v heterochromatinu Drosophila a v telomerách S. cerevisiae". EMBO J.. 16 (11): 3219–32. doi:10.1093 / emboj / 16.11.3219. PMC 1169939. PMID 9214638.
- ^ „NCBI UniGene“. Enhancer of zeste homolog 2 (Drosophila) (EZH2). Citováno 1. února 2015.
- ^ „Genové karty“. Enhancer Of Zeste Homolog 2 (Drosophila). Citováno 1. února 2015.
- ^ "Soubor". Genový strom EZH2. Citováno 1. února 2015.
- ^ Flicek P, Amode MR, Barrell D, Beal K, Billis K, Brent S a kol. (2014). „Ensembl 2014“. Nucleic Acids Res. 42 (Problém s databází): D749–55. doi:10.1093 / nar / gkt1196. PMC 3964975. PMID 24316576.
Další čtení
- Zeidler M, Kleer CG (září 2006). „Polycomb group protein Enhancer of Zeste 2: its links to DNA repair and breast cancer“. Journal of Molecular Histology. 37 (5–7): 219–23. doi:10.1007 / s10735-006-9042-9. PMID 16855786. S2CID 2332105.
- De Haan G, Gerrits A (červen 2007). "Epigenetická kontrola stárnutí hematopoetických kmenových buněk v případě Ezh2". Annals of the New York Academy of Sciences. 1106 (1): 233–9. doi:10.1196 / annals.1392.008. PMID 17332078. S2CID 25177748.
- Hobert O, Jallal B, Ullrich A (červen 1996). „Interakce Vav s ENX-1, domnělým transkripčním regulátorem genové exprese homeoboxu“. Molekulární a buněčná biologie. 16 (6): 3066–73. doi:10.1128 / MCB.16.6.3066. PMC 231301. PMID 8649418.
- Bonaldo MF, Lennon G, Soares MB (září 1996). „Normalizace a odčítání: dva přístupy k usnadnění objevování genů“. Výzkum genomu. 6 (9): 791–806. doi:10,1101 / gr. 6.9.791. PMID 8889548.
- Abel KJ, Brody LC, Valdes JM, Erdos MR, McKinley DR, Castilla LH, Merajver SD, Couch FJ, Friedman LS, Ostermeyer EA, Lynch ED, King MC, Welcsh PL, Osborne-Lawrence S, Spillman M, Bowcock AM, Collins FS, Weber BL (říjen 1996). "Charakterizace EZH1, lidského homologu Drosophila Enhancer zeste poblíž BRCA1". Genomika. 37 (2): 161–71. doi:10.1006 / geno.1996.0537. PMID 8921387.
- Laible G, Wolf A, Dorn R, Reuter G, Nislow C, Lebersorger A, Popkin D, Pillus L, Jenuwein T (červen 1997). "Savčí homology genu Polycomb-group Enhancer of zeste zprostředkovávají umlčení genů v heterochromatinu Drosophila a v telomerách S. cerevisiae". Časopis EMBO. 16 (11): 3219–32. doi:10.1093 / emboj / 16.11.3219. PMC 1169939. PMID 9214638.
- Cardoso C, Timsit S, Villard L, Khrestchatisky M, Fontès M, Colleaux L (duben 1998). „Specifická interakce mezi produktem genu XNP / ATR-X a doménou SET lidského proteinu EZH2“. Lidská molekulární genetika. 7 (4): 679–84. doi:10,1093 / hmg / 7,4,679. PMID 9499421.
- van Lohuizen M, Tijms M, Voncken JW, Schumacher A, Magnuson T, Wientjens E (červen 1998). „Interakce proteinů myší polycombové skupiny (Pc-G) Enx1 a Enx2 s Eed: indikace pro samostatné komplexy Pc-G“. Molekulární a buněčná biologie. 18 (6): 3572–9. doi:10.1128 / MCB.18.6.3572. PMC 108938. PMID 9584197.
- Sewalt RG, van der Vlag J, Gunster MJ, Hamer KM, den Blaauwen JL, Satijn DP, Hendrix T, van Driel R, Otte AP (červen 1998). „Charakterizace interakcí mezi savčími proteiny polycombových skupin Enx1 / EZH2 a EED naznačuje existenci různých komplexů proteinů savčích polycombových skupin“. Molekulární a buněčná biologie. 18 (6): 3586–95. doi:10,1128 / mcb.18.6.3586. PMC 108940. PMID 9584199.
- Denisenko O, Shnyreva M, Suzuki H, Bomsztyk K (říjen 1998). „Bodové mutace v doméně WD40 Eed blokují jeho interakci s Ezh2“. Molekulární a buněčná biologie. 18 (10): 5634–42. doi:10.1128 / MCB.18.10.5634. PMC 109149. PMID 9742080.
- van der Vlag J, Otte AP (prosinec 1999). „Transkripční represe zprostředkovaná proteinem lidské polycombové skupiny EED zahrnuje deacetylaci histonu“. Genetika přírody. 23 (4): 474–8. doi:10.1038/70602. PMID 10581039. S2CID 6748531.
- Cardoso C, Mignon C, Hetet G, Grandchamps B, Fontes M, Colleaux L (březen 2000). „Lidský gen EZH2: genomová organizace a revidované mapování v 7q35 v kritické oblasti pro maligní myeloidní poruchy“. European Journal of Human Genetics. 8 (3): 174–80. doi:10.1038 / sj.ejhg.5200439. PMID 10780782.
- Raaphorst FM, Otte AP, van Kemenade FJ, Blokzijl T, Fieret E, Hamer KM, Satijn DP, Meijer CJ (květen 2001). „Výrazné vzorce exprese BMI-1 a EZH2 v thymocytech a zralých T buňkách naznačují roli genů Polycomb v diferenciaci lidských T buněk“. Journal of Immunology. 166 (10): 5925–34. doi:10,4049 / jimmunol.166.10.5925. PMID 11342607.
- O'Connell S, Wang L, Robert S, Jones CA, Saint R, Jones RS (listopad 2001). „Polycomblike PHD prsty zprostředkovávají konzervovanou interakci s enhancerem proteinu zeste“. The Journal of Biological Chemistry. 276 (46): 43065–73. doi:10,1074 / jbc.M104294200. PMID 11571280.
- Varambally S, Dhanasekaran SM, Zhou M, Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt RG, Otte AP, Rubin MA, Chinnaiyan AM (říjen 2002). „Polycombový protein skupiny EZH2 se podílí na progresi rakoviny prostaty“ (PDF). Příroda. 419 (6907): 624–9. doi:10.1038 / nature01075. hdl:2027.42/62896. PMID 12374981. S2CID 4414767.
- Kleer CG, Cao Q, Varambally S, Shen R, Ota I, Tomlins SA, Ghosh D, Sewalt RG, Otte AP, Hayes DF, Sabel MS, Livant D, Weiss SJ, Rubin MA, Chinnaiyan AM (září 2003). „EZH2 je marker agresivního karcinomu prsu a podporuje neoplastickou transformaci buněk epitelu prsu“. Sborník Národní akademie věd Spojených států amerických. 100 (20): 11606–11. doi:10.1073 / pnas.1933744100. PMC 208805. PMID 14500907.