Přestavba chromatinu - Chromatin remodeling - Wikipedia
Přestavba chromatinu je dynamická modifikace chromatin architektura umožňující přístup kondenzované genomové DNA k regulačnímu proteiny transkripčního aparátu, a tím řídit genovou expresi. Taková přestavba se provádí hlavně 1) kovalentně histonové modifikace specifickými enzymy, např. histon acetyltransferázami (HAT), deacetylázami, methyltransferázami a kinázami a 2) komplexy remodelace chromatinu závislé na ATP, které se buď pohybují, vysouvají nebo restrukturalizují nukleosomy.[1] Kromě aktivní regulace genové exprese dodává dynamická remodelace chromatinu epigenetickou regulační roli v několika klíčových biologických procesech, replikaci a opravě DNA vaječných buněk; apoptóza; segregace chromozomů, stejně jako vývoj a pluripotence. Bylo zjištěno, že odchylky v proteinech remodelačních chromatinu jsou spojeny s lidskými chorobami, včetně rakoviny. Cílení na cesty remodelace chromatinu se v současné době vyvíjí jako hlavní terapeutická strategie v léčbě několika druhů rakoviny.
Přehled

Transkripční regulace genomu je řízena primárně na preiniciační fáze vazbou jaderných transkripčních strojních proteinů (jmenovitě RNA polymerázy, transkripčních faktorů a aktivátorů a represorů) na sekvenci promotoru jádra v kódující oblasti DNA. DNA je však v jádru pevně zabalena pomocí balicích proteinů, zejména histonových proteinů, které tvoří opakující se jednotky nukleosomy které se dále spojují a tvoří kondenzovanou strukturu chromatinu. Taková kondenzovaná struktura uzavírá mnoho regulačních oblastí DNA, což jim neumožňuje interagovat s proteiny transkripčního aparátu a regulovat genovou expresi. Abychom tento problém překonali a umožnili dynamický přístup ke kondenzované DNA, proces známý jako remodelace chromatinu mění architekturu nukleosomů, aby vystavil nebo skryl oblasti DNA pro regulaci transkripce.
Podle definice„Remodelace chromatinu je proces podporovaný enzymy, který usnadňuje přístup k nukleosomální DNA remodelací struktury, složení a umístění nukleosomů.
Klasifikace
Přístup k nukleosomální DNA je řízen dvěma hlavními třídami proteinových komplexů:
- Kovalentní komplexy modifikující histon.
- Komplexy remodelace chromatinu závislé na ATP.
Kovalentní komplexy modifikující histon
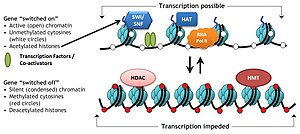
Specifické proteinové komplexy známé jako komplexy modifikující histon katalyzují přidání nebo odstranění různých chemických prvků na histonech. Tyto enzymatické modifikace zahrnují acetylace, methylace, fosforylace, a ubikvitinace a primárně se vyskytují na N-koncových histonových koncích. Takové modifikace ovlivňují vazebnou afinitu mezi histony a DNA, a tak uvolňují nebo utahují kondenzovanou DNA obalenou kolem histonů, např. Methylace specifických lysinových zbytků v H3 a H4 způsobuje další kondenzaci DNA kolem histonů, a tím brání vazbě transkripčních faktorů na DNA, která vede k genové represi. Naopak, acetylace histonu uvolňuje kondenzaci chromatinu a vystavuje DNA vazbě TF, což vede ke zvýšené genové expresi.[3]
Známé úpravy
Dobře charakterizované modifikace histonů zahrnují:[4]
Je známo, že jak lysinové, tak argininové zbytky jsou methylované. Methylované lysiny jsou nejlépe pochopitelné značky histonového kódu, protože specifický methylovaný lysin dobře odpovídá stavům genové exprese. Methylace lysinů H3K4 a H3K36 koreluje s transkripční aktivací, zatímco demethylace H3K4 koreluje s umlčením genomové oblasti. Methylace lysinů H3K9 a H3K27 koreluje s transkripční represí.[5] Zejména H3K9me3 vysoce koreluje s konstitutivním heterochromatinem.[6]
Acetylace má tendenci definovat „otevřenost“ chromatin protože acetylované histony se nemohou sbalit tak dobře jako deacetylované histony.
Existuje však mnohem více histonových modifikací a citlivých hmotnostní spektrometrie přístupy v poslední době výrazně rozšířily katalog.[7]
Histonový kód hypotéza
The histonový kód je hypotéza, že transkripce genetické informace kódované v DNA je částečně regulována chemickými modifikacemi histonových proteinů, zejména na jejich nestrukturovaných koncích. Spolu s podobnými úpravami, jako je Methylace DNA je součástí epigenetický kód.
Kumulativní důkazy naznačují, že takový kód je napsán specifickými enzymy, které mohou (například) methylátovou nebo acetylátovou DNA („spisovatelé“), odstraněny jinými enzymy, které mají aktivitu demetylázy nebo deacetylázy („gumy“), a nakonec je snadno identifikovatelný proteiny („ čtenáři), kteří jsou přijímáni k takovým histonovým modifikacím a váží se přes specifické domény, např. bromodoména, chromodoména. Tato trojitá akce „psaní“, „čtení“ a „mazání“ vytvářejí příznivé místní prostředí pro regulaci transkripce, opravu poškození DNA atd.[8]
Kritický koncept hypotéza histonového kódu je, že histonové modifikace slouží k náboru dalších proteinů specifickým rozpoznáním modifikovaného histonu prostřednictvím proteinové domény specializované pro tyto účely, spíše než pouhou stabilizací nebo destabilizací interakce mezi histonem a podkladovou DNA. Tyto přijaté proteiny pak aktivně mění strukturu chromatinu nebo podporují transkripci.
Níže je uveden velmi základní souhrn histonového kódu pro stav genové exprese (je popsána histonová nomenklatura tady ):
| Typ modifikace | Histone | ||||||
|---|---|---|---|---|---|---|---|
| H3K4 | H3K9 | H3K14 | H3K27 | H3K79 | H4K20 | H2BK5 | |
| mono-methylace | aktivace[9] | aktivace[10] | aktivace[10] | aktivace[10][11] | aktivace[10] | aktivace[10] | |
| di-methylace | represe[5] | represe[5] | aktivace[11] | ||||
| tri-methylace | aktivace[12] | represe[10] | represe[10] | aktivace,[11] represe[10] | represe[5] | ||
| acetylace | aktivace[12] | aktivace[12] | |||||
Remodelace chromatinu závislá na ATP
Komplexy remodelace chromatinu závislé na ATP regulují genovou expresi buď pohybem, vysunutím nebo restrukturalizací nukleosomů. Tyto proteinové komplexy mají společnou doménu ATPázy a energie z hydrolýzy ATP umožňuje těmto remodelačním komplexům přemístit nukleosomy (často označované jako „klouzání nukleosomů“) podél DNA, vysunout nebo sestavit histony na / z DNA nebo usnadnit výměnu histonu varianty, a tím vytvoření oblastí DNA bez nukleosomů pro aktivaci genů.[13] Několik předělávačů má také aktivitu translokace DNA k provádění konkrétních úkolů předělávání.[14]
Všechny komplexy remodelace chromatinu závislé na ATP mají podjednotku ATPázy, která patří do nadrodiny proteinů SNF2. V souvislosti s identitou podjednotky byly pro tyto proteiny klasifikovány dvě hlavní skupiny. Tito jsou známí jako skupina SWI2 / SNF2 a skupina imitace SWI (ISWI). Třetí třída komplexů závislých na ATP, která byla nedávno popsána, obsahuje ATPázu podobnou Snf2 a také demonstruje aktivitu deacetylázy.[15]
Známé komplexy remodelace chromatinu
U eukaryot existuje nejméně pět rodin remodelátorů chromatinu: SWI / SNF, ISWI, NuRD / Mi-2 /CHD „INO80 a SWR1, přičemž první dva remodelátory jsou zatím velmi dobře studovány, zejména u modelu kvasinek. Ačkoli všichni remodelátoři sdílejí společnou doménu ATPázy, jejich funkce jsou specifické na základě několika biologických procesů (oprava DNA, apoptóza atd.). To je způsobeno skutečností, že každý remodelační komplex má jedinečné proteinové domény (Helicase, bromodoména atd.) ve své katalytické oblasti ATPázy a má také různé přijaté podjednotky.
Specifické funkce
- Několik experimentů in vitro naznačuje, že remodelační systémy ISWI organizují nukleosomy do správné formy svazku a vytvářejí stejné mezery mezi nukleosomy, zatímco remodelační systémy SWI / SNF narušují nukleosomy.
- Ukázalo se, že remodelovače rodiny ISWI hrají ústřední roli v sestavování chromatinu po replikaci DNA a udržování struktur chromatinu vyššího řádu.
- Remodelátory rodiny INO80 a SWI / SNF se účastní opravy dvouvláknového zlomu DNA (DSB) a opravy nukleotidové excize (NER), a tím hrají klíčovou roli v reakci na poškození DNA zprostředkované TP53.
- NuRD / Mi-2 /CHD remodelační komplexy primárně zprostředkovávají transkripční represi v jádře a jsou nutné pro udržení pluripotence embryonálních kmenových buněk.[13]
Význam
V normálních biologických procesech
Remodelace chromatinu hraje ústřední roli v regulaci genové exprese tím, že poskytuje transkripčnímu aparátu dynamický přístup k jinak těsně zabalenému genomu. Pohyb nukleosomů remodelátory chromatinu je dále nezbytný pro několik důležitých biologických procesů, včetně sestavení a segregace chromozomů, replikace a opravy DNA, embryonálního vývoje a pluripotence a progrese buněčného cyklu. Deregulace remodelace chromatinu způsobuje ztrátu transkripční regulace v těchto kritických kontrolních bodech vyžadovaných pro správné buněčné funkce, a tím způsobuje různé syndromy onemocnění, včetně rakoviny.
Reakce na poškození DNA
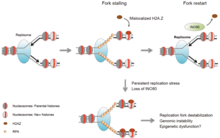
Relaxace chromatinu je jednou z prvních buněčných odpovědí na poškození DNA.[16] Uvolnění se zdá být iniciováno PARP1, jehož akumulace při poškození DNA je napůl úplná do 1,6 sekundy po poškození DNA.[17] Poté rychle následuje akumulace remodelovače chromatinu Alc1, který má ADP-ribóza –Vázací doména, která umožňuje rychlé přilákání k produktu PARP1. K maximálnímu náboru Alc1 dojde do 10 sekund od poškození DNA.[16] Asi polovina maximální relaxace chromatinu, pravděpodobně v důsledku působení Alc1, nastane o 10 sekund.[16] Akce PARP1 v místě dvouvláknového zlomu umožňuje nábor dvou DNA opravných enzymů MRE11 a NBS1. Poloviční maximální nábor těchto dvou DNA opravných enzymů trvá 13 sekund pro MRE11 a 28 sekund pro NBS1.[17]
Další proces relaxace chromatinu po vytvoření dvouřetězcového zlomu DNA využívá γH2AX, fosforylovanou formu H2AX protein. The histon varianta H2AX tvoří asi 10% histonů H2A v lidském chromatinu.[18] γH2AX (fosforylovaný na serinu 139 H2AX) byl detekován 20 sekund po ozáření buněk (s tvorbou dvouřetězcového zlomu DNA) a poloviční maximální akumulace yH2AX nastala za jednu minutu.[18] Rozsah chromatinu s fosforylovaným yH2AX je asi dva miliony párů bází v místě dvouřetězcového zlomu DNA.[18]
γH2AX sám o sobě nezpůsobuje dekondenzaci chromatinu, ale během několika sekund po ozařování protein „Mediator of the DNA damage checkpoint 1“ (MDC1 ) konkrétně se váže na γH2AX.[19][20] To je doprovázeno současnou akumulací RNF8 protein a protein pro opravu DNA NBS1 které se vážou k MDC1 protože MDC1 se váže na γH2AX.[21] RNF8 zprostředkovává rozsáhlou dekondenzaci chromatinu prostřednictvím své následné interakce s CHD4 protein,[22] součást komplexu remodelace nukleosomů a deacetylázy NuRD. Akumulace CHD4 v místě dvouvláknového zlomu je rychlá, k poloviční maximální akumulaci dochází do 40 sekund po ozáření.[23]
Po rychlé relaxaci počátečního chromatinu po poškození DNA (s rychlým zahájením opravy DNA) následuje pomalá rekondenzace, přičemž chromatin obnoví stav zhutnění blízko své úrovně před poškozením za ~ 20 minut.[16]
Rakovina
Remodelace chromatinu poskytuje jemné doladění v rozhodujících krocích buněčného růstu a dělení, jako je progrese buněčného cyklu, oprava DNA a segregace chromozomů, a proto vykonává funkci potlačující nádor. Mutace v takových remodelačních systémech chromatinu a deregulované kovalentní modifikace histonů potenciálně upřednostňují soběstačnost v buněčném růstu a únik z růstově regulačních buněčných signálů - dva důležité znaky rakovina.[24]
- Inaktivační mutace v systému SMARCB1, dříve známý jako hSNF5 / INI1 a součást člověka SWI / SNF remodelační komplex byly nalezeny ve velkém počtu rhabdoidní nádory, běžně postihující dětskou populaci.[25] Podobné mutace jsou také přítomny u jiných dětských rakovin, jako jsou karcinom plexus choroid, meduloblastom a u některých akutních leukémií. Dále studie knock-out myší silně podporují SMARCB1 jako tumor supresorový protein. Od původního pozorování mutací SMARCB1 v rhabdoidních nádorech bylo nalezeno několik dalších podjednotek komplexu remodelace lidského SWI / SNF chromatinu v široké škále novotvarů.[26]
- The SWI / SNF ATPase BRG1 (nebo 4 ) je nejčastěji mutovanou ATPázou remodelující chromatin u rakoviny.[27] Mutace v tomto genu byly poprvé rozpoznány v lidských rakovinných buněčných liniích získaných z nadledvin[28] a plíce.[29] U rakoviny vykazují mutace v BRG1 neobvykle vysokou preferenci missense mutací, které cílí na doménu ATPázy.[30][27] Mutace jsou obohaceny o vysoce konzervované sekvence ATPázy,[31] které leží na důležitých funkčních površích, jako je kapsa ATP nebo povrch vázající DNA.[30] Tyto mutace působí geneticky dominantním způsobem a mění regulační funkci chromatinu u enhancerů[30] a promotéry.[31]
- Fúzní protein PML-RAR v Akutní myeloidní leukémie rekrutuje histonové deacetylázy. To vede k represi genu odpovědného za diferenciaci myelocytů, což vede k leukémii.
- Tumor supresorový Rb protein funguje prostřednictvím náboru lidských homologů SWI / SNF enzymů BRG1, histondeacetylázy a DNA methyltransferázy. Mutace v BRG1 jsou hlášeny u několika druhů rakoviny způsobujících ztrátu tumor-supresorového působení Rb.[32]
- Nedávné zprávy naznačují hypermethylaci DNA v promotorové oblasti hlavních tumor supresorových genů u několika druhů rakoviny. Přestože u histonmethyltransferáz je dosud hlášeno několik mutací, byla u několika typů rakoviny, zejména u rakoviny tlustého střeva a prsu, hlášena korelace hypermethylace DNA a methylace histonu H3 lysinu-9.
- Mutace v histonacetyltransferázách (HAT) p300 (missense a zkrácený typ) jsou nejčastěji hlášeny u kolorektálních, pankreatických, karcinomů prsu a žaludku. Ztráta heterozygotnosti v kódující oblasti p300 (chromozom 22q13) je přítomna u velkého počtu glioblastomů.
- Kromě toho HAT mají různou roli jako transkripční faktory kromě toho, že mají aktivitu histonacetylázy, např. HAT podjednotka, hADA3 může působit jako adaptační protein spojující transkripční faktory s jinými HAT komplexy. Při absenci hADA3 je transkripční aktivita TP53 významně snížena, což naznačuje roli hADA3 při aktivaci funkce TP53 v reakci na poškození DNA.
- Podobně bylo prokázáno, že TRRAP, lidský homolog s kvasinkami Tra1, přímo interaguje s c-Myc a E2F1 - známými onkoproteiny.
Genomika rakoviny
Rychlý postup dovnitř genomika rakoviny a vysokou propustností Čip ChIP, ChIP-sekv a Bisulfitové sekvenování metody poskytují lepší vhled do role remodelace chromatinu v regulaci transkripce a role v rakovině.
Terapeutický zásah
Epigenetická nestabilita způsobená deregulací při remodelaci chromatinu je studována u několika druhů rakoviny, včetně rakoviny prsu, rakoviny tlustého střeva a konečníku, rakoviny slinivky břišní. Taková nestabilita do značné míry způsobuje rozsáhlé umlčování genů s primárním dopadem na geny potlačující nádory. Proto se nyní zkouší strategie k překonání epigenetického umlčování pomocí synergické kombinace Inhibitory HDAC nebo HDI a DNA-demetylační činidla.HDI se primárně používají jako doplňková léčba u několika typů rakoviny.[33][34] Inhibitory HDAC mohou indukovat p21 (WAF1) výraz, regulátor p53 je supresivní aktivita nádoru. HDAC jsou zapojeny do cesty, kterou retinoblastomový protein (pRb) potlačuje proliferace buněk.[35] Estrogen je dobře zavedený jako a mitogenní faktor podílí se na tumorigenezi a progresi rakovina prsu prostřednictvím jeho vazby na estrogenový receptor alfa (ERα). Nedávná data naznačují, že inaktivace chromatinu zprostředkovaná HDAC a methylací DNA je kritickou složkou umlčování ERα v lidských buňkách rakoviny prsu.[36]
- Schválené použití:
- Vorinostat byl licencován Americký FDA v říjnu 2006 k léčbě kožní T-lymfom (CTCL).
- Romidepsin (obchodní název Istodax) byl licencován americkým FDA v listopadu 2009 pro kožní T-buněčný lymfom (CTCL).
- Fáze III Klinické studie:
- Panobinostat (LBH589) je v klinických studiích pro různé druhy rakoviny, včetně studie fáze III pro kožní T buněčný lymfom (CTCL).
- Kyselina valproová (jako Mg valproát) ve studiích fáze III pro rakovina děložního hrdla a rakovina vaječníků.
- Zahájeny stěžejní klinické studie fáze II
- Belinostat (PXD101) podstoupil studii fáze II pro relabující rakovina vaječníků a uvedl dobré výsledky pro T-lymfom.
- Inhibitory HDAC
Současní kandidáti na nové cíle v oblasti drog jsou Histon lysin methyltransferázy (KMT) a protein arginin methyltransferázy (PRMT).[37]
Jiné syndromy nemoci
- ATRX syndrom (mentální retardace spojená s α-thalassemií X) a syndrom myelodysplasie α-thalassemie jsou způsobeny mutacemi v ATRX, ATPáza související s SNF2 s PHD.
- Syndrom CHARGE, autosomálně dominantní porucha, byla nedávno spojena s haploinsufficiencí CHD7, který kóduje Rodina CHD ATPáza CHD7.[38]
Stárnutí
Chromatinová architektonická přestavba se podílí na procesu buněčná stárnutí, který souvisí a přesto se liší od, stárnutí organismu. Replikativní buněčná senescence označuje trvalou buněčný cyklus zatknout, kde pomitotický buňky nadále existují jako metabolicky aktivní buňky, ale nedokáží množit se.[39][40] Senescence může vzniknout kvůli degradace související s věkem n, opotřebení telomer, progerias, před malignitami a další formy poškození nebo nemoc. Senescentní buňky procházejí zřetelnými represivními fenotypovými změnami, potenciálně k zabránění množení poškozených nebo rakovinných buněk, s upravenými organizace chromatinu fluktuace v hojnosti remodelátorů a změny v epigenetické modifikace.[41][42][39] Stárnoucí buňky procházejí chromatinová krajina úpravy jako konstitutivní heterochromatin migruje do středu jádra a přemisťuje se euchromatin a fakultativní heterochromatin do oblastí na okraji jádra. To narušuje chromatinlamin interakce a inverze vzoru typicky pozorovaného v mitoticky aktivní buňce.[43][41] Jednotlivé domény spojené s laminem (LAD) a Topologicky sdružující domény (TAD) jsou touto migrací narušeny, což může ovlivnit cis interakce napříč genomem.[44] Kromě toho existuje obecný vzor kanonického histon ztráta, zejména pokud jde o nukleosom histony H3 a H4 a linker histon H1.[43] Varianty histonu se dvěma exony jsou upregulovány v senescentních buňkách za vzniku modifikované sestavy nukleosomů, což přispívá k permisivitě chromatinu k senescentním změnám.[44] Ačkoli transkripce variantních histonových proteinů může být zvýšena, kanonické histonové proteiny nejsou exprimovány, protože jsou vytvářeny pouze během S fáze buněčného cyklu a senescentní buňky jsou postmitotické.[43] Během stárnutí, části chromozomy lze exportovat z jádra pro lysozomální degradace což má za následek větší organizační zmatek a narušení interakcí chromatinu.[42]
Množství remodelátoru chromatinu může být zapleteno do buněčné stárnutí jako srazit nebo knokaut ATP závislých remodelátorů, jako jsou NuRD, ACF1 a SWI / SNP, může mít za následek poškození DNA a senescentní fenotypy v kvasinkách, C. elegans, myši a lidské buněčné kultury.[45][42][46] ACF1 a NuRD jsou downregulovány v senescentních buňkách, což naznačuje, že remodelace chromatinu je nezbytná pro udržení mitotického fenotypu.[45][46] Geny podílející se na signalizaci stárnutí mohou být umlčeny potvrzením chromatinu a polykombovými represivními komplexy, jak je patrné v umlčování PRC1 / PCR2 p16.[47][48] Konkrétní deplece remodelovače vede k aktivaci proliferativních genů v důsledku nedodržení umlčování.[42] Někteří remodelátoři působí spíše na oblasti zesilovače genů než na konkrétní lokusy, aby zabránili opětovnému vstupu do buněčného cyklu tvorbou oblastí hustého heterochromatinu kolem regulačních oblastí.[48]
Senescentní buňky procházejí rozsáhlými fluktuacemi epigenetických modifikací ve specifických oblastech chromatinu ve srovnání s mitotickými buňkami. Lidské a myší buňky procházející replikačním stárnutím zažívají obecný globální pokles methylace; konkrétní loci se však mohou lišit od obecného trendu.[49][44][42][47] Specifické oblasti chromatinu, zejména ty kolem promotorů nebo zesilovačů proliferativních lokusů, mohou vykazovat zvýšené methylační stavy s celkovou nerovnováhou represivních a aktivujících histonových modifikací.[41] Proliferativní geny mohou vykazovat zvýšení represivní značky H3K27me3 zatímco geny podílející se na umlčení nebo aberantní histonové produkty mohou být obohaceny aktivační modifikací H3K4me3.[44] Dále upregulování histonových deacetyláz, jako jsou členové sirtuin rodina, může oddálit stárnutí odstraněním acetylových skupin, které přispívají k větší dostupnosti chromatinu.[50] Obecná ztráta methylace v kombinaci s přidáním acetylových skupin vede k přístupnější konformaci chromatinu s tendencí k dezorganizaci ve srovnání s mitoticky aktivními buňkami.[42] Obecná ztráta histonů vylučuje přidání histonových modifikací a přispívá ke změnám v obohacení v některých oblastech chromatinu během stárnutí.[43]
Viz také
- Epigenetika
- Histone
- Nukleosomy
- Chromatin
- Histon acetyltransferáza
- Transkripční faktory
- CAF-1 (Chromatinový montážní faktor-1) - histonový chaperon, který vykonává koordinační roli při remodelaci chromatinu.
Reference
- ^ Teif VB, Rippe K (září 2009). „Predikce pozic nukleosomů na DNA: kombinace preferencí vnitřní sekvence a aktivit remodelace“. Výzkum nukleových kyselin. 37 (17): 5641–55. doi:10.1093 / nar / gkp610. PMC 2761276. PMID 19625488.
- ^ Boyer (2009). "Podpis chromatinu pluripotentních buněk". Stembook. doi:10.3824 / stembook.1.45.1. PMID 20614601.
- ^ Wang GG, Allis CD, Chi P (září 2007). "Remodelace chromatinu a rakovina, Část I: Kovalentní histonové modifikace". Trendy v molekulární medicíně. 13 (9): 363–72. doi:10.1016 / j.molmed.2007.07.003. PMID 17822958.
- ^ Strahl BD, Allis CD (leden 2000). "Jazyk kovalentních modifikací histonu". Příroda. 403 (6765): 41–5. Bibcode:2000Natur.403 ... 41S. doi:10.1038/47412. PMID 10638745. S2CID 4418993.
- ^ A b C d Rosenfeld JA, Wang Z, Schones DE, Zhao K, DeSalle R, Zhang MQ (březen 2009). „Stanovení obohacených modifikací histonu v negenických částech lidského genomu“. BMC Genomics. 10: 143. doi:10.1186/1471-2164-10-143. PMC 2667539. PMID 19335899.
- ^ Hublitz P, Albert M, Peters A (28. dubna 2009). "Mechanismy transkripční represe methylací histonového lysinu". International Journal of Developmental Biology. 10 (1387): 335–354. doi:10,1387 / ijdb.082717ph. ISSN 1696-3547. PMID 19412890.
- ^ Tan M, Luo H, Lee S, Jin F, Yang JS, Montellier E, Buchou T, Cheng Z, Rousseaux S, Rajagopal N, Lu Z, Ye Z, Zhu Q, Wysocka J, Ye Y, Khochbin S, Ren B , Zhao Y (září 2011). „Identifikace 67 histonových značek a krotononylace histon-lysinu jako nový typ modifikace histonu“. Buňka. 146 (6): 1016–28. doi:10.1016 / j.cell.2011.08.008. PMC 3176443. PMID 21925322.
- ^ Jenuwein T, Allis CD (srpen 2001). "Překlad histonového kódu". Věda. 293 (5532): 1074–80. CiteSeerX 10.1.1.453.900. doi:10.1126 / science.1063127. PMID 11498575. S2CID 1883924.
- ^ Benevolenskaya EV (srpen 2007). „Histonové H3K4 demethylázy jsou nezbytné pro vývoj a diferenciaci“. Biochemie a buněčná biologie. 85 (4): 435–43. doi:10.1139 / o07-057. PMID 17713579.
- ^ A b C d E F G h Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K (květen 2007). „Profilování methylací histonu v lidském genomu s vysokým rozlišením“. Buňka. 129 (4): 823–37. doi:10.1016 / j.cell.2007.05.009. PMID 17512414. S2CID 6326093.
- ^ A b C Steger DJ, Lefterova MI, Ying L, Stonestrom AJ, Schupp M, Zhuo D, Vakoc AL, Kim JE, Chen J, Lazar MA, Blobel GA, Vakoc CR (duben 2008). „Nábor DOT1L / KMT4 a methylace H3K79 jsou všudypřítomně spojeny s genovou transkripcí v savčích buňkách“. Molekulární a buněčná biologie. 28 (8): 2825–39. doi:10.1128 / MCB.02076-07. PMC 2293113. PMID 18285465.
- ^ A b C Koch CM, Andrews RM, Flicek P, Dillon SC, Karaöz U, Clelland GK, Wilcox S, Beare DM, Fowler JC, Couttet P, James KD, Lefebvre GC, Bruce AW, Dovey OM, Ellis PD, Dhami P, Langford CF , Weng Z, Birney E, Carter NP, Vetrie D, Dunham I (červen 2007). „Krajina modifikací histonu přes 1% lidského genomu v pěti lidských buněčných liniích“. Výzkum genomu. 17 (6): 691–707. doi:10,1101 / gr. 5704207. PMC 1891331. PMID 17567990.
- ^ A b Wang GG, Allis CD, Chi P (září 2007). „Remodelace a rakovina chromatinu, část II: Remodelace chromatinu závislá na ATP“. Trendy v molekulární medicíně. 13 (9): 373–80. doi:10.1016 / j.molmed.2007.07.004. PMC 4337864. PMID 17822959.
- ^ Saha A, Wittmeyer J, Cairns BR (červen 2006). „Chromatinová přestavba: průmyslová revoluce DNA kolem histonů“. Nature Reviews Molecular Cell Biology. 7 (6): 437–47. doi:10.1038 / nrm1945. PMID 16723979. S2CID 6180120.
- ^ Vignali, M .; Hassan, A. H .; Neely, K. E .; Workman, J. L. (2000-03-15). „Komplexy remodelační na chromatinu závislé na ATP“. Molekulární a buněčná biologie. 20 (6): 1899–1910. doi:10.1128 / mcb.20.6.1899-1910.2000. ISSN 0270-7306. PMC 110808. PMID 10688638.
- ^ A b C d Sellou H, Lebeaupin T, Chapuis C, Smith R, Hegele A, Singh HR, Kozlowski M, Bultmann S, Ladurner AG, Timinszky G, Huet S (2016). „Poly (ADP-ribóza) závislý remodelovač chromatinu Alc1 indukuje lokální relaxaci chromatinu při poškození DNA“. Mol. Biol. Buňka. 27 (24): 3791–3799. doi:10,1091 / mbc.E16-05-0269. PMC 5170603. PMID 27733626.
- ^ A b Haince JF, McDonald D, Rodrigue A, Déry U, Masson JY, Hendzel MJ, Poirier GG (2008). „PARP1-dependentní kinetika náboru proteinů MRE11 a NBS1 do více míst poškození DNA“. J. Biol. Chem. 283 (2): 1197–208. doi:10,1074 / jbc.M706734200. PMID 18025084.
- ^ A b C Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM (1998). „Dvouřetězcové zlomy DNA indukují fosforylaci histonu H2AX na serinu 139“. J. Biol. Chem. 273 (10): 5858–68. doi:10.1074 / jbc.273.10.5858. PMID 9488723.
- ^ Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C, Lukas J (2007). „RNF8 ubikvityláty histonů při dvouřetězcových zlomech DNA a podporuje shromažďování opravných proteinů“. Buňka. 131 (5): 887–900. doi:10.1016 / j.cell.2007.09.040. PMID 18001824. S2CID 14232192.
- ^ Stucki M, Clapperton JA, Mohammad D, Yaffe MB, Smerdon SJ, Jackson SP (2005). „MDC1 přímo váže fosforylovaný histon H2AX k regulaci buněčných odpovědí na dvouřetězcové zlomy DNA“. Buňka. 123 (7): 1213–26. doi:10.1016 / j.cell.2005.09.038. PMID 16377563.
- ^ Chapman JR, Jackson SP (2008). „Fosfo-závislé interakce mezi NBS1 a MDC1 zprostředkovávají retenci chromatinu komplexu MRN v místech poškození DNA“. EMBO Rep. 9 (8): 795–801. doi:10.1038 / embor.2008.103. PMC 2442910. PMID 18583988.
- ^ Luijsterburg MS, Acs K, Ackermann L, Wiegant WW, Bekker-Jensen S, Larsen DH, Khanna KK, van Attikum H, Mailand N, Dantuma NP (2012). „Nová nekatalytická role ubikvitin ligázy RNF8 při rozvíjení struktury chromatinu vyššího řádu“. EMBO J.. 31 (11): 2511–27. doi:10.1038 / emboj.2012.104. PMC 3365417. PMID 22531782.
- ^ Smeenk G, Wiegant WW, Vrolijk H, Solari AP, Pastink A, van Attikum H (2010). „Komplex remodelace NuRD chromatinu reguluje signalizaci a opravu poškození DNA“. J. Cell Biol. 190 (5): 741–9. doi:10.1083 / jcb.201001048. PMC 2935570. PMID 20805320.
- ^ Hanahan D, Weinberg RA (leden 2000). "Charakteristické znaky rakoviny". Buňka. 100 (1): 57–70. doi:10.1016 / S0092-8674 (00) 81683-9. PMID 10647931. S2CID 1478778.
- ^ Versteege I, Sévenet N, Lange J, Rousseau-Merck MF, Ambros P, Handgretinger R, Aurias A, Delattre O (červenec 1998). "Zkrácení mutací hSNF5 / INI1 u agresivní dětské rakoviny". Příroda. 394 (6689): 203–6. Bibcode:1998 Natur.394..203V. doi:10.1038/28212. PMID 9671307. S2CID 6019090.
- ^ Shain AH, Pollack JR (2013). "Spektrum mutací SWI / SNF, všudypřítomné u lidských rakovin". PLOS ONE. 8 (1): e55119. Bibcode:2013PLoSO ... 855119S. doi:10.1371 / journal.pone.0055119. PMC 3552954. PMID 23355908.
- ^ A b Hodges C, Kirkland JG, Crabtree GR (srpen 2016). „Mnoho rolí komplexů BAF (mSWI / SNF) a PBAF v rakovině“. Perspektivy Cold Spring Harbor v medicíně. 6 (8): a026930. doi:10.1101 / cshperspect.a026930. PMC 4968166. PMID 27413115.
- ^ Dunaief JL, Strober BE, Guha S, Khavari PA, Alin K, Luban J, Begemann M, Crabtree GR, Goff SP (říjen 1994). „Protein retinoblastomu a BRG1 tvoří komplex a spolupracují při indukci zástavy buněčného cyklu.“ Buňka. 79 (1): 119–30. doi:10.1016/0092-8674(94)90405-7. PMID 7923370. S2CID 7058539.
- ^ Medina PP, Romero OA, Kohno T, Montuenga LM, Pio R, Yokota J, Sanchez-Cespedes M (květen 2008). „Časté mutace inaktivující BRG1 / SMARCA4 v buněčných liniích lidské rakoviny plic“. Lidská mutace. 29 (5): 617–22. doi:10.1002 / humu.20730. PMID 18386774.
- ^ A b C Hodges HC, Stanton BZ, Cermakova K, Chang CY, Miller EL, Kirkland JG, Ku WL, Veverka V, Zhao K, Crabtree GR (leden 2018). „Dominantní negativní mutanti SMARCA4 mění prostředí přístupnosti zesilovačů bez omezení tkáně“. Přírodní strukturní a molekulární biologie. 25 (1): 61–72. doi:10.1038 / s41594-017-0007-3. PMC 5909405. PMID 29323272.
- ^ A b Stanton BZ, Hodges C, Calarco JP, Braun SM, Ku WL, Kadoch C, Zhao K, Crabtree GR (únor 2017). "Smarca4 ATPázové mutace narušují přímé vystěhování PRC1 z chromatinu". Genetika přírody. 49 (2): 282–288. doi:10.1038 / ng.3735. PMC 5373480. PMID 27941795.
- ^ Wolffe AP (květen 2001). „Přestavba chromatinu: proč je důležitá u rakoviny“. Onkogen. 20 (24): 2988–90. doi:10.1038 / sj.onc.1204322. PMID 11420713.
- ^ Marks PA, Dokmanovic M (prosinec 2005). "Inhibitory histon-deacetylázy: objev a vývoj jako protirakovinné látky". Znalecký posudek na vyšetřované léky. 14 (12): 1497–511. doi:10.1517/13543784.14.12.1497. PMID 16307490. S2CID 1235026.
- ^ Richon VM, O'Brien JP (2002). „Inhibitory histon-deacetylázy: nová třída potenciálních terapeutických látek pro léčbu rakoviny“ (PDF). Klinický výzkum rakoviny. 8 (3): 662–4. PMID 11895892.
- ^ Richon VM, Sandhoff TW, Rifkind RA, Marks PA (srpen 2000). „Inhibitor histon-deacetylázy selektivně indukuje expresi p21WAF1 a acetylaci histonu související s genem“. Sborník Národní akademie věd Spojených států amerických. 97 (18): 10014–9. Bibcode:2000PNAS ... 9710014R. doi:10.1073 / pnas.180316197. PMC 27656. PMID 10954755.
- ^ Zhang Z, Yamashita H, Toyama T, Sugiura H, Ando Y, Mita K, Hamaguchi M, Hara Y, Kobayashi S, Iwase H (listopad 2005). "Kvantifikace exprese mRNA HDAC1 v invazivním karcinomu prsu *". Výzkum a léčba rakoviny prsu. 94 (1): 11–6. doi:10.1007 / s10549-005-6001-1. PMID 16172792. S2CID 27550683.
- ^ Dowden J, Hong W, Parry RV, Pike RA, Ward SG (duben 2010). „Směrem k vývoji silných a selektivních inhibitorů bisubstrátů protein arginin methyltransferáz“. Dopisy o bioorganické a léčivé chemii. 20 (7): 2103–5. doi:10.1016 / j.bmcl.2010.02.069. PMID 20219369.
- ^ Clapier CR, Cairns BR (2009). "Biologie komplexů remodelovajících chromatin". Roční přehled biochemie. 78: 273–304. doi:10,1146 / annurev.biochem.77.062706.153223. PMID 19355820.
- ^ A b Parry, Aled John; Narita, Masashi (2016). "Staré buňky, nové triky: struktura chromatinu v senescence". Savčí genom. 27 (7–8): 320–331. doi:10.1007 / s00335-016-9628-9. ISSN 0938-8990. PMC 4935760. PMID 27021489.
- ^ Hayflick, L .; Moorhead, P. S. (01.12.1961). "Sériová kultivace kmenů lidských diploidních buněk". Experimentální výzkum buněk. 25 (3): 585–621. doi:10.1016/0014-4827(61)90192-6. ISSN 0014-4827. PMID 13905658.
- ^ A b C Chandra, Tamir; Ewels, Philip Andrew; Schoenfelder, Stefan; Furlan-Magaril, Mayra; Wingett, Steven William; Kirschner, Kristina; Thuret, Jean-Yves; Andrews, Simon; Fraser, Peter; Reik, Vlk (2015-01-29). „Globální reorganizace jaderné krajiny v senescentních buňkách“. Zprávy buněk. 10 (4): 471–483. doi:10.1016 / j.celrep.2014.12.055. ISSN 2211-1247. PMC 4542308. PMID 25640177.
- ^ A b C d E F Sun, Luyang; Yu, Ruofan; Dang, Weiwei (2018-04-16). "Chromatinové architektonické změny během buněčného stárnutí a stárnutí". Geny. 9 (4): 211. doi:10,3390 / geny9040211. ISSN 2073-4425. PMC 5924553. PMID 29659513.
- ^ A b C d Criscione, Steven W .; Teo, Yee Voan; Neretti, Nicola (2016). „Chromatinová krajina buněčné stárnutí“. Trendy v genetice. 32 (11): 751–761. doi:10.1016 / j.tig.2016.09.005. ISSN 0168-9525. PMC 5235059. PMID 27692431.
- ^ A b C d Yang, Na; Sen, Payel (03.11.2018). "Senescentní buněčný epigenom". Stárnutí (Albany NY). 10 (11): 3590–3609. doi:10.18632 / stárnutí.101617. ISSN 1945-4589. PMC 6286853. PMID 30391936.
- ^ A b Basta, Jeannine; Rauchman, Michael (2015). „Komplex remodelace a deacetylázy nukleosomů (NuRD) ve vývoji a nemoci“. Translační výzkum. 165 (1): 36–47. doi:10.1016 / j.trsl.2014.05.003. ISSN 1931-5244. PMC 4793962. PMID 24880148.
- ^ A b Li, Xueping; Ding, Dong; Yao, červen; Zhou, Bin; Shen, Ting; Qi, Yun; Ni, Ting; Wei, Gang (2019-07-15). „Faktor remodelace chromatinu BAZ1A reguluje stárnutí buněk jak v rakovině, tak v normálních buňkách“. Humanitní vědy. 229: 225–232. doi:10.1016 / j.lfs.2019.05.023. ISSN 1879-0631. PMID 31085244.
- ^ A b López-Otín, Carlos; Blasco, Maria A .; Partridge, Linda; Serrano, Manuel; Kroemer, Guido (06.06.2013). „Charakteristické znaky stárnutí“. Buňka. 153 (6): 1194–1217. doi:10.1016 / j.cell.2013.05.039. ISSN 0092-8674. PMC 3836174. PMID 23746838.
- ^ A b Tasdemir, Nilgun; Banito, Ana; Roe, Jae-Seok; Alonso-Curbelo, Direna; Camiolo, Matthew; Tschaharganeh, Darjus F .; Huang, Chun-Hao; Aksoy, Ozlem; Bolden, Jessica E .; Chen, Chi-Chao; Fennell, Myles (2016). „BRD4 připojuje remodelaci zesilovače k imunitnímu sledování Senescence“. Objev rakoviny. 6 (6): 612–629. doi:10.1158 / 2159-8290.CD-16-0217. ISSN 2159-8290. PMC 4893996. PMID 27099234.
- ^ Wilson, V. L .; Jones, P. A. (06.06.1983). „Methylace DNA klesá ve stárnutí, ale ne v nesmrtelných buňkách“. Věda. 220 (4601): 1055–1057. Bibcode:1983Sci ... 220,1055 W.. doi:10.1126 / science.6844925. ISSN 0036-8075. PMID 6844925.
- ^ Kaeberlein, Matt; McVey, Mitch; Guarente, Leonard (01.10.1999). „Samotný komplex SIR2 / 3/4 a SIR2 podporují dlouhověkost v Saccharomyces cerevisiae dvěma různými mechanismy“. Geny a vývoj. 13 (19): 2570–2580. doi:10.1101 / gad.13.19.2570. ISSN 0890-9369. PMC 317077. PMID 10521401.
Další čtení
- Chen T, Dent SY (únor 2014). "Chromatinové modifikátory a remodelátory: regulátory buněčné diferenciace". Genetika hodnocení přírody. 15 (2): 93–106. doi:10.1038 / nrg3607. PMC 3999985. PMID 24366184.
externí odkazy
- MBInfo - Chromatin
- MBInfo - balení DNA
- YouTube - Chromatin, histony a úpravy
- YouTube - přehled epigenetiky
- Chromatin + remodelace v americké národní lékařské knihovně Lékařské předměty (Pletivo)