Inositol-trisfosfát 3-kináza - Inositol-trisphosphate 3-kinase - Wikipedia
| Inositol trisfosfát 3-kináza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Inositol-trisfosfát 3-kináza A Katalytické jádro. 1 TZD | |||||||||
| Identifikátory | |||||||||
| EC číslo | 2.7.1.127 | ||||||||
| Číslo CAS | 106283-10-7 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
Inositol (1,4,5) trifosfát 3-kináza (ES 2.7.1.127 ), zkráceně zde ITP3K, je enzym, který usnadňuje přenos fosfoskupiny z adenosintrifosfát na 1D-myo-inositol 1,4,5-trisfosfát. Tento enzym patří do rodiny transferázy, konkrétně ty, které přenášejí skupiny obsahující fosfor (fosfotransferázy ) s alkoholovou skupinou jako akceptorem. The systematické jméno z této třídy enzymů je ATP: 1D-myo-inositol-1,4,5-trisfosfát-3-fosfotransferáza. ITP3K katalyzuje přenos gama-fosfátu z ATP do 3 polohy inositol 1,4,5-trisfosfát za vzniku inositol 1,3,4,5-tetrakisfosfátu.[1] ITP3K je vysoce specifický pro 1,4,5-izomer IP3, a to výhradně fosforyláty pozice 3-OH, produkující Ins (1,3,4,5) P4, také známý jako inositol tetrakisphosphate nebo IP4.
V biologii je enzym ITP3K zkrácen řadou různých způsobů, včetně 1D-myo-inositol-trisfosfát 3-kináza, ITP3K, ITPK, IP3-kináza, IP3-3-kináza, Ins (1,4,5) P3 3-kináza. Kromě toho lze enzym pojmenovat jako produkt jednoho ze 3 genů u lidí ITPKA, ITPKB, a ITPKC nebo jeden ze dvou v ovocných muškách, IP3K1 a IP3K2 —Mutant známý genetikům jako vlnitý.[2] Genom hlístic má jednu formu enzymu, kódovanou Gen LFE-2. Enzymy ITP3K jsou exprimovány pouze v metazoans; nejsou exprimovány v kvasinkách nebo rostlinách.
Všechny ITP3K patří do větší konstrukční rodiny, inositol polyfosfát kinázy nebo IPK. Všimněte si však, že lidský genom také obsahuje gen pro jinou kinázu známý jako ITPK1, což je inositol 1, 3, 4-trisfosfát 5/6-kináza a není členem rodiny IPK.
Rodina enzymů ITP3K je někdy zaměňována s jinou rodinou enzymů, která má podobný název, tj fosfatidyl inositol 3-kinázy nebo fosfoinositid 3-kináza (PI3-K), jehož substráty jsou lipidy inositolu, nikoli rozpustný druhý posel inositol trisphosphate.
Objev a charakterizace
Vědecký zájem o inositol fosfáty zesílil v letech následujících po objevu roku 1983 inositol trisphosphate byl intracelulární posel, který uvolňuje vápník z intracelulárních zásob v endoplazmatickém retikulu.[3] Na konci tohoto desetiletí bylo objeveno velké množství inositolfosfátkináz a fosfatáz, včetně ITP3K v roce 1986.[4][5]Biochemické a molekulární studie v 90. letech vedly k čištění enzymu z mozku krysy a jeho molekulární klonování a tyto studie odhalily různé mechanismy zpětné vazby, kterými je enzym regulován vápníkovými a proteinovými kinázami.[6] V roce 1999 byl ITP3K identifikován jako člen větší rodiny Inositol polyfosfát kinázy, které sdílejí podobnou strukturu a katalytický mechanismus.[7][8] Enzymy ITP3K sdílejí společné strukturní rysy včetně konzervovaného katalytického jádra, které váže ATP umístěné blízko C-konce, a různých regulačních domén blíže k N-konci.[9]
Katalytická doména
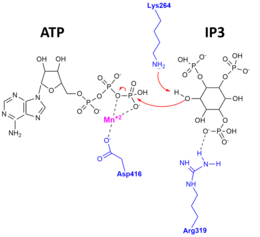
Důkazy pro tuto vynikající specificitu a pro katalytický mechanismus byly nalezeny, když apoenzym, komplex vázaný na substrát a komplex vázaný na produkt Rentgenové krystalové struktury z ITPKA byly určeny.[10][11] Na obrázku vpravo je znázorněn katalytický mechanismus, kterým je 3'OH IP3 napadá gama-fosfát ATP a aminokyselinové zbytky ITPK důležité pro stabilizaci substrátů a produktů v aktivním místě.
Ukázalo se, že struktura katalytické domény lidské ITP3KA je rozdělena do tří subdomén. Tyto subdomény jsou zobrazeny jako N laloku, což je N-terminální doména, C laloku, který je C-terminální subdoménou a třetí alfa-pouze subdoménou. Katalytická doména ITP3K se poněkud liší od nadrodiny proteinových kináz a má novou doménu vázající substrát se čtyřmi šroubovicemi. V tomhle kináza, jsou dvě domény v otevřené konformaci, což naznačuje, že obě domény jsou přístupné současně. To naznačuje Podklad uznání a katalýza ITP3K zahrnuje dynamický konformační cyklus. Navíc tato jedinečná šroubovicová doména ITPK blokuje přístup k Aktivní stránky fosfoinositidy vázanými na membránu, vysvětlující strukturní základ pro rozpustný specificita inositol polyfosfátu. Dalším znakem katalytického jádra je vazebné místo ATP. Tady, jedna molekula ADP je vázán v rozštěpu hlavní domény, což naznačuje aktivní místo kinázy.
Podrobněji, větší doména proteinová struktura má strukturu třídy α / β. Doména má N-terminál a C-terminální lalok s rozštěpem mezi nimi a každý z těchto laloků je postaven kolem antiparalelní β-list. Na N-konci má list tři vlákna, zatímco na C-konci je pětivláknový list. Druhá doména je α-helikální a skládá se ze čtyř α helixů spojených dlouhými smyčkami. Spirály jsou volně zabalené proti sobě a celá doména je ve srovnání s velkou doménou α / β vysoce mobilní. Spirálovitá doména je umístěna vedle sebe na jednom konci rozštěpu ve velké doméně.
Nařízení
ITP3K je regulován různými posttranslační mechanismy. ITP3K jsou stimulovány přímo vápník / kalmodulin (Ca2 + / CaM) vazba.[12] Obecně jsou savčí ITP3K aktivovány vápníkem a kalmodulinem v různé míře. Metoda, ve které to funguje, je kalmodulin rozpoznává sekvence, které obsahují amfifilní alfa-helixy se shluky kladně nabitých a hydrofobní aminokyseliny.[13] Určité sekvence jsou vyžadovány pro vazbu CaM a enzym aktivace a tato úroveň stimulace se jeví jako specifická pro buňku, tkáň nebo izoformu. ITP3K z hlístice a Arabidopsis thaliana postrádají vazebná místa pro CaM, a proto jsou necitlivé na vápník a kalmodulin.[14] Další významnou posttranslační modifikací, která je důležitá pro regulaci ITP3K, je fosforylace. Aktivita ITP3K je nepřímo stimulována fosforylací pomocí kináza II závislá na vápníku / kalmodulinu (CaMKII). Kromě toho existují důkazy, že ITP3K mohou být aktivovány při fosforylaci pomocí protein kináza C (PKC) a inhibována při fosforylaci pomocí protein kináza A (PKA).
Izoformy
Existují tři ITP3K, které jsou kódovány lidským genomem: ITPKA, ITPKB, a ITPKC. Všechny sdílejí konzervovanou C-koncovou katalytickou doménu, ale liší se v mechanismech regulace i v tkáňové expresi. ITPKA převládá v neurony a v testy. Je lokalizován na dendritické trny sdružením s vlákny aktin což odpovídá jeho pravděpodobné roli v paměťových funkcích. ITPKB je exprimován častěji, ale je často obohacen v imunitních tkáních a má různé intracelulární lokalizace, které závisí na tkáni, interakci s aktinovými vlákny a proteolýze v N-koncových oblastech. ITPKC je také exprimován v mnoha různých tkáních a je více obohacen v jádře ve srovnání s jinými izoformami.
Funkce při signalizaci vápníku
ITP3K hraje roli při regulaci nebo spolupráci s intracelulárními buňkami vápníkové signály které nastanou po osvobození inositol trisphosphate. V této cestě buď Receptor spojený s G-proteinem (GPCR) nebo receptor tyrosinkináza (RTK) je aktivován extracelulární vazbou ligandu. Zahájení cesty vede k aktivaci G-alfa podjednotka a heterotrimerní G protein (v případě zprostředkování GPCR signální transdukce ) nebo autofosorylace RTK cytoplazmatických domén (v případě RTK zprostředkované signální transdukce). Tyto intracelulární události nakonec vedou k aktivaci fosfolipáza C (PLC), který štěpí fosfolipid PIP2 do diacylglycerol (DAG) a inositol 1,4,5-trisfosfát (IP3). DAG zůstává spojena s plazmatická membrána, zatímco IP3 se uvolňuje do cytoplazma. IP3 pak difunduje cytosolem a váže se na IP3 receptory na endoplazmatické retikulum nebo sarkoplazmatické retikulum, což má za následek otevření membránového kanálu a příliv vápník ionty do cytoplazmy.[15] Vápník slouží jako a druhý posel pro různé následné buněčné události včetně glykogen metabolismus, svalová kontrakce, uvolňování neurotransmiteru, a transkripční regulace.[15] Proto je homeostáza vápníku nezbytná pro správnou funkci buněk a reakci na extracelulární signály.[16]
Aby se buňka připravila na budoucí signalizační událost, musí být dráha vápníku přísně regulována. Zdá se, že ITP3K hraje důležitou roli při ukončení signálu. Jak již bylo zmíněno, ITP3K katalyzuje fosforylaci IP3 udělat IP4. Na rozdíl od IP3, IP4 nezpůsobuje otevírání vápníkových kanálů na endoplazmatickém retikulu nebo sarkoplazmatickém retikulu.[17] Snížením koncentrace IP3 v cytoplazmě ITP3K ukončuje šíření vápníkové signální dráhy.[14]

Další role
ITP3K není jediným enzymem odpovědným za čištění IP3 z cytoplazmy. Volal druhý enzym inositol 5-fosfatáza katalyzuje defosforylace IP3 vytvořit IP2.[18] Příroda obvykle neupřednostňuje vývoj druhého enzymu, který by vykonával již existující identickou funkci.[19] Bližší zkoumání evoluční historie inositol 5-fosfatázy a ITP3K vede k několika zajímavým hypotézám o rolích těchto enzymů v buňce.
Inositol 5-fosfatáza existovala předtím, než se v buňce savců vyvinul ITP3K. Jako ostatní fosfatázy, inositol 5-fosfatáza je energeticky nezávislý enzym, který štěpí a fosfát skupina mimo a Podklad.[20] Naproti tomu ITP3K (jako všechny kinázy ) je energeticky závislý, což znamená, že k provádění chemie přenosu fosforylu vyžaduje molekulu ATP.[21] Pokud už příroda měla energeticky nezávislý mechanismus pro ukončení signální dráhy vápníku, proč byl vývoj ITP3K výhodný? To je zřejmé redundance funkce nebo „plýtvání“ energií buňkou naznačuje, že ITP3K může mít v buňce důležitější funkci než pouhé vymazání IP3 druhý posel z cytoplazmy.[20] Aktuální hypotézy o dalších rolích pro ITPK jsou vysvětleny v následujících dvou podsekcích.
Produktem ITPK může být druhý posel
Jak již bylo zmíněno dříve, ITP3K katalyzuje reakci přenosu fosforylu, která převádí IP3 do IP4. IP4 nestimuluje příliv vápníku prostřednictvím IP3 receptorové kanály na endoplazmatickém nebo sarkoplazmatickém retikulu. Ukázalo se však, že IP4 stimuluje otevření kalciového kanálu na plazmatické membráně. Tímto způsobem IP4 může ve skutečnosti sloužit k prodloužení signálu vápníku aktivací přílivu zásob vápníku z extracelulárního prostoru. Kromě toho existují důkazy, že IP4 váže dva Proteiny aktivující GTPázu, GAP1IP4BP a GAP1m.[18] GAP se často používají při přenosu signálu jako vypínače. IP4 vazba na GAP naznačuje, že ITPK může být zapojen do paralelní signální transdukční dráhy. Přesná role IP4 vazba na tyto GAP však nebyla stanovena, takže k získání úplnějšího porozumění bude zapotřebí dalšího výzkumu v této oblasti.[22]
Role v metabolismu inositolfosfátu
Kromě svých potenciálních rolí jako druhý posel, IP4 může také fungovat jako zásadní předchůdce pro jiné více fosforylované fosfáty inositolu, jako je IP5, IP6, IP7a IP8. Taková údržba je nezbytná pro přípravu buňky na budoucí příchozí signál.[22]
Význam pro fyziologii a lidské choroby
ITPKA protein je vysoce obohacen o dendritické trny.[23] ITPKA se účastní procesu učení a paměti v neuronálních buňkách, a to jak svou katalytickou aktivitou, tak interakcí s vláknitým aktinem.
Ačkoli ITPKA je fyziologicky exprimován v neuronech a varlatech, gen se stává exprimován v řadě typů rakovinných buněk. Ve většině případů exprese ITP3K způsobuje, že rakovina je agresivnější.[24]
ITPKB se podílí na fyziologické imunitní funkci.[25]
ITPKC byl spojován s Kawasakiho chorobou, autoimunitní poruchou.[26][27]
Reference
- ^ „UniProtKB - P23677 (IP3KA_HUMAN)“. Citováno 19. února 2015.
- ^ Dean DM, Maroja LS, Cottrill S, Bomkamp BE, Westervelt KA, Deitcher DL (listopad 2015). „Zvlněné mutační mapy genu inositol 1,4,5-trisfosfát 3-kinázy 2 (IP3K2) Drosophila a interakce s IP3R ovlivňují vývoj křídel“. G3. 6 (2): 299–310. doi:10,1534 / g3,115,024307. PMC 4751550. PMID 26613949.
- ^ Streb H, Irvine RF, Berridge MJ, Schulz I (1983). "Uvolňování Ca2 + z nonmitochondriálního intracelulárního úložiště v pankreatických acinárních buňkách inositol-1,4,5-trisfosfátem". Příroda. 306 (5938): 67–9. Bibcode:1983 Natur.306 ... 67S. doi:10.1038 / 306067a0. PMID 6605482. S2CID 4359904.
- ^ Irvine RF, Letcher AJ, Heslop JP, Berridge MJ (1986). „Inositol tris / tetrakisfosfátová dráha - demonstrace aktivity Ins (1,4,5) P3 3-kinázy ve zvířecích tkáních“. Příroda. 320 (6063): 631–4. Bibcode:1986 Natur.320..631I. doi:10.1038 / 320631a0. PMID 3010126. S2CID 4249596.
- ^ Hansen CA, Mah S, Williamson JR (červen 1986). "Tvorba a metabolismus inositol 1,3,4,5-tetrakisfosfátu v játrech". The Journal of Biological Chemistry. 261 (18): 8100–3. PMID 3487541.
- ^ Takazawa K, Vandekerckhove J, Dumont JE, Erneux C (listopad 1990). „Klonování a exprese cDNA mozků potkanů kódujících Ca2 + / kalmodulin-senzitivní inositol 1,4,5-trisfosfát 3-kinázu v Escherichia coli“. The Biochemical Journal. 272 (1): 107–12. doi:10.1042 / bj2720107. PMC 1149663. PMID 2176078.
- ^ Saiardi A, Erdjument-Bromage H, Snowman AM, Tempst P, Snyder SH (listopad 1999). "Syntéza difosfoinositol pentakisfosfátu nově identifikovanou rodinou vyšších inositol polyfosfát kináz". Aktuální biologie. 9 (22): 1323–6. doi:10.1016 / s0960-9822 (00) 80055-x. PMID 10574768. S2CID 15311443.
- ^ Odom AR, Stahlberg A, Wente SR, York JD (březen 2000). „Role jaderné inositol 1,4,5-trisfosfátkinázy v transkripční kontrole“. Věda. 287 (5460): 2026–9. Bibcode:2000Sci ... 287.2026O. doi:10.1126 / science.287.5460.2026. PMID 10720331.
- ^ Schell MJ (červen 2010). „Inositol trisfosfát 3-kinázy: zaměření na imunitní a neuronální signalizaci“. Buněčné a molekulární biologické vědy. 67 (11): 1755–78. doi:10.1007 / s00018-009-0238-5. PMID 20066467. S2CID 25121695.
- ^ González B, Schell MJ, Letcher AJ, Veprintsev DB, Irvine RF, Williams RL (září 2004). „Struktura lidské inositol 1,4,5-trisfosfát 3-kinázy: vazba na substrát odhaluje, proč nejde o fosfoinositid 3-kinázu“. Molekulární buňka. 15 (5): 689–701. doi:10.1016 / j.molcel.2004.08.004. PMID 15350214.
- ^ Miller GJ, Hurley JH (září 2004). "Krystalová struktura katalytického jádra inositol 1,4,5-trisfosfát 3-kinázy". Molekulární buňka. 15 (5): 703–11. doi:10.1016 / j.molcel.2004.08.005. PMID 15350215.
- ^ Lloyd-Burton SM, Yu JC, Irvine RF, Schell MJ (březen 2007). „Regulace inositol 1,4,5-trisfosfát 3-kináz vápníkem a lokalizace v buňkách“. The Journal of Biological Chemistry. 282 (13): 9526–35. doi:10,1074 / jbc.M610253200. PMID 17284449.
- ^ Franco-Echevarría E, Baños-Sanz JI, Monterroso B, kolo A, Sanz-Aparicio J, González B (listopad 2014). „Nový motiv vázající kalmodulin pro regulaci inositol 1,4,5-trisfosfát 3-kinázy“. The Biochemical Journal. 463 (3): 319–28. doi:10.1042 / BJ20140757. PMID 25101901.
- ^ A b Xia HJ, Yang G (únor 2005). „Inositol 1,4,5-trisfosfát 3-kinázy: funkce a předpisy“. Cell Research. 15 (2): 83–91. doi:10.1038 / sj.cr.7290270. PMID 15740635.
- ^ A b Berridge MJ (leden 1993). "Signalizace inositoltrisfosfátu a vápníku". Příroda. 361 (6410): 315–25. Bibcode:1993 Natur.361..315B. doi:10.1038 / 361315a0. PMID 8381210. S2CID 4362262.
- ^ Voet, Donald Voet, Judith G. (2011). Biochemie (4. vydání). Hoboken, NJ: John Wiley & Sons. ISBN 978-0-470-57095-1.
- ^ Havas N (srpen 2011). „Zpátky ve vodě“. Journal of Paliative Medicine. 14 (8): 971–2. doi:10.1089 / jpm.2011.0043. PMID 21809925.
- ^ A b Pattni K, Banting G (červen 2004). "Ins (1,4,5) P3 metabolismus a rodina IP3-3Kinas". Mobilní signalizace. 16 (6): 643–54. doi:10.1016 / j.cellsig.2003.10.009. PMID 15093605.
- ^ „Pochopení evoluce“. Citováno 19. února 2015.
- ^ A b Irvine RF, Lloyd-Burton SM, Yu JC, Letcher AJ, Schell MJ (2006). „Regulace a funkce inositol 1,4,5-trisfosfát 3-kináz“. Pokroky v regulaci enzymů. 46 (1): 314–23. doi:10.1016 / j.advenzreg.2006.01.009. PMC 1820747. PMID 16857241.
- ^ „WikiKinome“. Kinase.com. Citováno 19. února 2015.
- ^ A b Irvine RF, Schell MJ (květen 2001). „Zpátky ve vodě: návrat inositol fosfátů“. Recenze přírody. Molekulární buněčná biologie. 2 (5): 327–38. doi:10.1038/35073015. PMID 11331907. S2CID 2259401.
- ^ Yamada M, Kakita A, Mizuguchi M, Rhee SG, Kim SU, Ikuta F (březen 1993). „Specifická exprese inositol 1,4,5-trisfosfát 3-kinázy v dendritických trnech“. Výzkum mozku. 606 (2): 335–40. doi:10.1016 / 0006-8993 (93) 91004-c. PMID 8387863. S2CID 10790958.
- ^ Windhorst S, Fliegert R, Blechner C, Möllmann K, Hosseini Z, Günther T, Eiben M, Chang L, Lin HY, Fanick W, Schumacher U, Brandt B, Mayr GW (únor 2010). „Inositol 1,4,5-trisfosfát 3-kináza-A je nový protein podporující motilitu buněk, který zvyšuje metastatický potenciál nádorových buněk o dvě funkční aktivity“. The Journal of Biological Chemistry. 285 (8): 5541–54. doi:10.1074 / jbc.M109.047050. PMC 2820782. PMID 20022963.
- ^ Miller AT, Dahlberg C, Sandberg ML, Wen BG, Beisner DR, Hoerter JA, Parker A, Schmedt C, Stinson M, Avis J, Cienfuegos C, McPate M, Tranter P, Gosling M, Groot-Kormelink PJ, Dawson J, Pan S, Tian SS, Seidel HM, Cooke MP (2015). „Inhibice Inositol kinázy Itpkb zvyšuje signalizaci vápníku v lymfocytech a odhaluje novou strategii léčby autoimunitních onemocnění“. PLOS ONE. 10 (6): e0131071. Bibcode:2015PLoSO..1031071M. doi:10.1371 / journal.pone.0131071. PMC 4488288. PMID 26121493.
- ^ Onouchi Y, Gunji T, Burns JC, Shimizu C, Newburger JW, Yashiro M, Nakamura Y, Yanagawa H, Wakui K, Fukushima Y, Kishi F, Hamamoto K, Terai M, Sato Y, Ouchi K, Saji T, Nariai A Kaburagi Y, Yoshikawa T, Suzuki K, Tanaka T, Nagai T, Cho H, Fujino A, Sekine A, Nakamichi R, Tsunoda T, Kawasaki T, Nakamura Y, Hata A (leden 2008). „ITPKC funkční polymorfismus spojený s náchylností k Kawasakiho chorobě a tvorbou aneuryzmatu koronárních tepen“. Genetika přírody. 40 (1): 35–42. doi:10.1038 / ng.2007.59. PMC 2876982. PMID 18084290.
- ^ Alphonse MP, Duong TT, Shumitzu C, Hoang TL, McCrindle BW, Franco A, Schurmans S, Philpott DJ, Hibberd ML, Burns J, Kuijpers TW, Yeung RS (listopad 2016). „Inositol-trifosfát 3-kináza C zprostředkovává aktivaci zánětlivých buněk a reakci na léčbu u Kawasakiho choroby“. Journal of Immunology. 197 (9): 3481–3489. doi:10,4049 / jimmunol. 1600388. PMID 27694492.