Polyadenylace - Polyadenylation

Polyadenylace je přidání a poly (A) ocas k transkriptu RNA, obvykle a messenger RNA (mRNA). Poly (A) ocas se skládá z několika adenosinmonofosfáty; jinými slovy, je to úsek RNA, který má pouze adenin základny. v eukaryoty, polyadenylace je součástí procesu, který produkuje zralou mRNA pro překlad. V mnoha bakterie, poly (A) ocas podporuje degradaci mRNA. Je tedy součástí širšího procesu genová exprese.
Proces polyadenylace začíná jako transkripce a gen končí. The 3'-nejvíce segment nově vyrobené pre-mRNA je nejprve odštěpen a sada bílkovin; tyto proteiny pak syntetizují poly (A) ocas na 3 'konci RNA. V některých genech tyto proteiny přidávají poly (A) ocas na jednom z několika možných míst. Polyadenylace proto může produkovat více než jeden transkript z jednoho genu (alternativní polyadenylace), podobný alternativní sestřih.[1]
Poly (A) ocas je důležitý pro jaderný export, translaci a stabilitu mRNA. Ocas se časem zkracuje a když je dostatečně krátký, mRNA se enzymaticky odbourává.[2] U několika typů buněk jsou však mRNA s krátkými poly (A) konci uloženy pro pozdější aktivaci re-polyadenylací v cytosolu.[3] Naproti tomu, když k polyadenylaci dochází u bakterií, podporuje degradaci RNA.[4] To je také někdy případ eukaryotů nekódující RNA.[5][6]
molekuly mRNA jak u prokaryot, tak u eukaryot mají polyadenylované 3'-konce, přičemž prokaryotické poly (A) konce jsou obecně kratší a méně molekul mRNA polyadenylované.[7]
Pozadí RNA
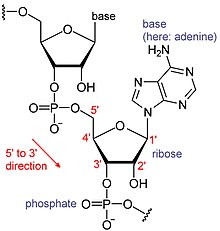
- Další informace viz RNA a Messenger RNA
RNA jsou typem velkých biologických molekul, jejichž jednotlivé stavební kameny se nazývají nukleotidy. Název poly (A) ocas (pro ocas polyadenylové kyseliny)[8] odráží způsob zkrácení RNA nukleotidů, s písmenem pro bázi, kterou nukleotid obsahuje (A pro adenin, C pro cytosin, G pro guanin a U pro uracil ). RNA jsou produkovány (přepsal ) z a DNA šablona. Podle konvence jsou sekvence RNA psány ve směru 5 'až 3'. 5 'konec je část molekuly RNA, která je přepsána jako první, a 3' konec je přepsán jako poslední. 3 'konec je také místem, kde se poly (A) ocas nachází na polyadenylovaných RNA.[1][9]
Messenger RNA (mRNA) je RNA, která má kódující oblast, která funguje jako templát pro syntézu proteinů (překlad ). Zbytek mRNA, nepřeložené regiony, naladit, jak aktivní je mRNA.[10] Existuje také mnoho RNA, které nejsou překládány, nazývaných nekódující RNA. Stejně jako nepřekládané oblasti má mnoho z těchto nekódujících RNA regulační role.[11]
Jaderná polyadenylace
Funkce
V nukleární polyadenylaci je poly (A) ocas přidán k RNA na konci transkripce. Na mRNA chrání poly (A) ocas molekulu mRNA před enzymatickou degradací v cytoplazma a pomáhá při ukončení transkripce, exportu mRNA z jádra a translaci.[2] Téměř všechny eukaryotické mRNA jsou polyadenylované,[12] s výjimkou závislých na replikaci zvířat histon mRNA.[13] Toto jsou jediné mRNA v eukaryotech, které postrádají poly (A) ocas, končící místo a kmenová smyčka Struktura následovaná sekvencí bohatou na puriny, nazývanou histonový downstream prvek, která řídí, kde je štěpena RNA, takže je vytvořen 3 'konec histonové mRNA.[14]
Mnoho eukaryotických nekódujících RNA je vždy na konci transkripce polyadenylováno. Existují malé RNA, kde poly (A) ocas je vidět pouze ve zprostředkujících formách a ne ve zralé RNA, protože konce jsou během zpracování odstraněny, ty pozoruhodné jsou mikroRNA.[15][16] Ale pro mnohé dlouhé nekódující RNA - zdánlivě velká skupina regulační RNA, které například obsahují RNA Xist, který zprostředkovává Inaktivace chromozomu X - poly (A) ocas je součástí zralé RNA.[17]
Mechanismus
| Zapojené proteiny:[12][18] CPSF: faktor specificity štěpení / polyadenylace |
The procesivní polyadenylační komplex v jádru eukaryot pracuje na produktech RNA polymeráza II, jako prekurzor mRNA. Zde multi-proteinový komplex (viz komponenty vpravo)[18] štěpí 3'-největší část nově vyrobené RNA a polyadenyláty konce produkovaného tímto štěpením. Štěpení je katalyzováno enzymem CPSF[13][18] a vyskytuje se 10–30 nukleotidů pod jeho vazebným místem.[19] Toto místo má často na RNA polyadenylační signální sekvenci AAUAAA, ale její varianty, které se na CPSF existovat.[18][20] Dva další proteiny dodávají specificitu vazbě na RNA: CstF a CFI. CstF se váže na oblast bohatou na GU dále po proudu od místa CPSF.[21] CFI rozpoznává třetí místo na RNA (soubor sekvencí UGUAA u savců[22][23][24]) a může získat CPSF, i když chybí sekvence AAUAAA.[25][26] Polyadenylační signál - sekvenční motiv rozpoznávaný komplexem štěpení RNA - se mezi skupinami eukaryot liší. Většina lidských polyadenylačních míst obsahuje sekvenci AAUAAA,[21] ale tato sekvence je u rostlin a hub méně obvyklá.[27]
RNA je typicky štěpena před ukončením transkripce, protože CstF se také váže na RNA polymerázu II.[28] Prostřednictvím špatně pochopeného mechanismu (od roku 2002) signalizuje, že RNA polymeráza II vyklouzla z transkriptu.[29] Štěpení také zahrnuje protein CFII, i když není známo jak.[30] Místo štěpení spojené s polyadenylačním signálem se může lišit až do přibližně 50 nukleotidů.[31]
Když je RNA štěpena, začíná polyadenylace, katalyzovaná polyadenylát polymerázou. Polyadenylát polymeráza vytvoří poly (A) ocas přidáním adenosinmonofosfát jednotky z adenosintrifosfát odštěpením RNA pyrofosfát.[32] Další protein, PAB2, se váže na nový, krátký poly (A) konec a zvyšuje afinitu polyadenylát polymerázy k RNA. Když je poly (A) ocas přibližně 250 nukleotidy enzym se již nemůže vázat na CPSF a polyadenylační zastávky se zastaví, čímž se určí délka poly (A) ocasu.[33][34] CPSF je v kontaktu s RNA polymerázou II, což jí umožňuje signalizovat polymerázu k ukončení transkripce.[35][36] Když RNA polymeráza II dosáhne „terminační sekvence“ (⁵'TTTATT³ 'na templátu DNA a ⁵'AAUAAA³' na primárním transkriptu), je signalizován konec transkripce.[37] Polyadenylační aparát je také fyzicky spojen s spliceosome, komplex, který odstraňuje introny z RNA.[26]
Následné účinky
Poly (A) ocas funguje jako vazebné místo pro protein vázající poly (A). Protein vázající poly (A) podporuje export z jádra a translaci a inhibuje degradaci.[38] Tento protein se váže na poly (A) ocas před exportem mRNA z jádra a v kvasinkách také rekrutuje poly (A) nukleázu, enzym, který zkracuje poly (A) ocas a umožňuje export mRNA. Poly (A) -binding protein je exportován do cytoplazmy s RNA. mRNA, které nejsou exportovány, jsou degradovány exozom.[39][40] Poly (A) -binding protein se také může vázat na, a tak získávat, několik proteinů, které ovlivňují translaci,[39] jedním z nich je iniciační faktor -4G, což zase rekrutuje 40S ribozomální podjednotka.[41] Poly (A) ocas však není vyžadován pro translaci všech mRNA.[42] Poly (A) tailing (oligo-adenylation) can determine the osud of RNA addresses that are usually not poly (A) -tailed (such as (small) non-coding (sn) RNAs etc.), and indukce their RNA rozklad.[43]
Deadenylation
V eukaryotice somatické buňky, poly (A) ocasy většiny mRNA v cytoplazmě se postupně zkracují a mRNA s kratšími poly (A) ocasy jsou méně přeloženy a degradovány dříve.[44] Může však trvat mnoho hodin, než se mRNA degraduje.[45] Tento proces deadenylace a degradace může být urychlen pomocí mikroRNA doplňujících se k 3 'nepřekládaná oblast mRNA.[46] v nezralé vaječné buňky mRNA se zkrácenými poly (A) konci nejsou degradovány, ale jsou místo toho uloženy a translačně neaktivní. Tyto krátce sledované mRNA jsou aktivovány cytoplazmatickou polyadenylací po oplodnění během aktivace vajec.[47]
U zvířat poly (A) ribonukleáza (PARN ) se může vázat na 5 'čepice a odstranit nukleotidy z poly (A) ocasu. Úroveň přístupu k 5 'čepici a poly (A) ocasu je důležitá pro řízení, jak brzy je mRNA degradována. PARN deadenylates méně, pokud je RNA vázána iniciačními faktory 4E (na 5 'čepici) a 4G (na poly (A) ocasu), což je důvod, proč translace snižuje Deadenylation. Rychlost deadenylace může být také regulována proteiny vázajícími RNA. Jakmile je poly (A) ocas odstraněn, dekappingový komplex odstraní 5 'čepičku, což vede k degradaci RNA. Několik dalších proteinů se účastní deadenylace v nadějné droždí a lidské buňky, zejména CCR4-ne komplex.[48]
Cytoplazmatická polyadenylace
V cytosolu některých typů živočišných buněk, zejména v zárodečná linie, během rané embryogeneze a v post-synaptický stránky nervové buňky. Tím se prodlouží poly (A) ocas mRNA se zkráceným poly (A) ocasem, takže mRNA bude přeloženo.[44][49] Tyto zkrácené poly (A) ocasy jsou často menší než 20 nukleotidů a jsou prodlouženy na přibližně 80–150 nukleotidů.[3]
U raného myšího embrya umožňuje cytoplazmatická polyadenylace mateřských RNA z vaječné buňky buňce přežít a růst, i když transkripce nezačne až do poloviny 2-buněčného stádia (4-buněčného stadia u člověka).[50][51] V mozku je cytoplazmatická polyadenylace aktivní během učení a mohla by hrát roli v dlouhodobé potenciace, což je posílení přenosu signálu z nervové buňky do jiné v reakci na nervové impulsy a je důležité pro učení a formování paměti.[3][52]
Cytoplazmatická polyadenylace vyžaduje proteiny vázající RNA CPSF a CPEB a může zahrnovat další proteiny vázající RNA, jako je Pumilio.[53] V závislosti na typu buňky může být polymeráza stejný typ polyadenylát polymerázy (PAP), který se používá v jaderném procesu, nebo cytoplazmatická polymeráza GLD-2.[54]
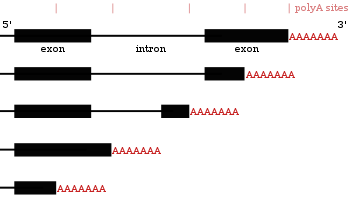
Alternativní polyadenylace
Mnoho genů kódujících proteiny má více než jedno polyadenylační místo, takže gen může kódovat několik mRNA, které se liší v jejich 3 'konec.[27][55][56] 3 'oblast přepisu obsahuje mnoho polyadenylačních signálů (PAS). Když se používají proximálnější (blíže k 5 'konci) PAS místa, zkrátí to délku 3' nepřekládané oblasti (3 'UTR) transkriptu.[57] Studie u lidí i much ukázaly tkáňově specifické APA. S neuronálními tkáněmi preferujícími distální PAS, což vede k delšímu 3 'UTR a tkáně varlat preferující proximální PAS vedoucí k kratším 3' UTR.[58][59] Studie ukázaly, že existuje korelace mezi úrovní zachování genu a jeho tendencí k alternativní polyadenylaci, přičemž vysoce konzervované geny vykazují více APA. Podobně vysoce exprimované geny sledují stejný vzorec.[60] Ribo-sekvenování údaje (sekvenování pouze mRNA uvnitř ribozomů) ukázaly, že izoformy mRNA s kratšími 3 'UTR jsou pravděpodobně přeloženy.[57]
Protože alternativní polyadenylace mění délku 3 'UTR,[61] může také změnit, pro která vazebná místa jsou k dispozici mikroRNA ve 3 'UTR.[19][62] MicroRNA mají tendenci potlačovat translaci a podporovat degradaci mRNA, na které se vážou, i když existují příklady mikroRNA, které stabilizují transkripty.[63][64] Alternativní polyadenylace může také zkrátit kódující oblast, čímž se vytvoří mRNA kód pro jiný protein,[65][66] ale to je mnohem méně časté než pouhé zkrácení 3 'nepřekládané oblasti.[27]
Volba poly (A) místa může být ovlivněna extracelulárními podněty a závisí na expresi proteinů, které se účastní polyadenylace.[67][68] Například výraz CstF-64, podjednotka stimulační faktor štěpení (CstF), zvyšuje se makrofágy v reakci na lipopolysacharidy (skupina bakteriálních sloučenin, které vyvolávají imunitní odpověď). To má za následek výběr slabých poly (A) míst a tedy kratších transkriptů. Tím se odstraní regulační prvky ve 3 'nepřekládaných oblastech mRNA u produktů souvisejících s obranou lysozym a TNF-α. Tyto mRNA pak mají delší poločasy a produkují více těchto proteinů.[67] Jiné proteiny vázající RNA než ty, které jsou v polyadenylačním aparátu, mohou také ovlivnit, zda je použito polyadenylační místo,[69][70][71][72] jak může Methylace DNA blízko polyadenylačního signálu.[73]
Označení pro degradaci u eukaryot
Pro mnoho nekódující RNA, počítaje v to tRNA, rRNA, snRNA, a snoRNA, polyadenylace je způsob značení RNA pro degradaci, alespoň v droždí.[74] Tato polyadenylace se v jádře provádí pomocí Komplex TRAMP, který udržuje ocas dlouhý přibližně 4 nukleotidy až k 3 'konci.[75][76] RNA je poté degradována exozom.[77] Poly (A) ocasy byly také nalezeny na fragmentech lidské rRNA, a to jak ve formě homopolymerních (pouze A), tak heterpolymerních (většinou A) ocasů.[78]
U prokaryot a organel

V mnoha bakteriích mohou být polyadenylovány jak mRNA, tak nekódující RNA. Tento poly (A) ocas podporuje degradaci degradosom, který obsahuje dva enzymy degradující RNA: polynukleotid fosforyláza a RNáza E. Polynukleotid fosforyláza se váže na 3 'konec RNA a 3' prodloužení poskytované poly (A) ocasem mu umožňuje vázat se na RNA, jejichž sekundární struktura jinak by zablokoval 3 'konec. Postupná kola polyadenylace a degradace 3 'konce polynukleotid fosforylázou umožňují degradosom překonat tyto sekundární struktury. Poly (A) ocas může také rekrutovat RNasy, které štěpí RNA na dvě části.[79] Tyto bakteriální poly (A) ocasy jsou dlouhé asi 30 nukleotidů.[80]
V tak odlišných skupinách jako zvířata a trypanosomy, mitochondrie obsahují stabilizační i destabilizující poly (A) ocasy. Destabilizující polyadenylace cílí na mRNA i nekódující RNA. Poly (A) ocasy mají průměrně 43 nukleotidů. Stabilizační ty začínají na stop kodonu a bez nich není stop kodon (UAA) úplný, protože genom kóduje pouze U nebo UA část. Rostlinné mitochondrie mají pouze destabilizující polyadenylaci. Mitochondriální polyadenylace nebyla nikdy pozorována u začínajících ani štěpných kvasinek. [81][82]
Zatímco mnoho bakterií a mitochondrií má polyadenylátové polymerázy, mají také jiný typ polyadenylace prováděné polynukleotid fosforyláza sám. Tento enzym se nachází v bakteriích,[83] mitochondrie,[84] plastidy[85] a jako složka archaeal exosome (v těch archaea které mají exozom ).[86] Může syntetizovat 3 'rozšíření, kde převážnou většinu bází tvoří adeniny. Podobně jako u bakterií podporuje polyadenylace polynukleotid fosforylázou degradaci RNA v plastidech[87] a pravděpodobně také archaea.[81]
Vývoj
Přestože je polyadenylace vidět téměř ve všech organismech, není univerzální.[7][88] Široká distribuce této modifikace a skutečnost, že je přítomna v organismech ze všech tří domén života znamená, že poslední univerzální společný předek Předpokládá se, že všech živých organismů měla nějakou formu polyadenylačního systému.[80] Několik organismů polyadenylát mRNA neimplikuje, což znamená, že během evoluce ztratily své polyadenylační stroje. Ačkoli nejsou známy žádné příklady eukaryot, které postrádají polyadenylaci, mRNA z bakterie Mycoplasma gallisepticum a sůl-tolerantní archaean Haloferax volcanii chybí tato modifikace.[89][90]
Nejstarší polyadenylační enzym je polynukleotid fosforyláza. Tento enzym je součástí bakterie degradosom a archaeal exozom,[91] dva blízce příbuzné komplexy, které recyklují RNA na nukleotidy. Tento enzym degraduje RNA napadením vazby mezi 3'-nejvíce nukleotidy fosforečnanem a odštěpením difosfátového nukleotidu. Tato reakce je reverzibilní, a proto může enzym také rozšiřovat RNA o více nukleotidů. Heteropolymerní ocas přidaný polynukleotid fosforylázou je velmi bohatý na adenin. Volba adeninu je s největší pravděpodobností výsledkem vyššího ADP koncentrace než jiné nukleotidy v důsledku použití ATP jako energetická měna, takže je pravděpodobnější, že bude začleněna do tohoto ocasu v časných formách života. Bylo navrženo, že zapojení ocasů bohatých na adenin při degradaci RNA vedlo k pozdějšímu vývoji polyadenylátových polymeráz (enzymů, které produkují poly (A) ocasy bez dalších nukleotidů).[92]
Polyadenylátové polymerázy nejsou tak staré. Samostatně se vyvinuli jak z bakterií, tak z eukaryot Enzym přidávající CCA, což je enzym, který dokončuje 3 'konce tRNA. Jeho katalytická doména je homologní s doménou ostatních polymerázy.[77] Předpokládá se, že horizontální přenos bakteriálního enzymu přidávajícího CCA na eukaryota umožnil archaealnímu enzymu přidávajícímu CCA přepnout funkci na poly (A) polymerázu.[80] Některé linie, jako archaea a sinice, se nikdy nevyvinula polyadenylát polymeráza.[92]
Polyadenylátové ocasy jsou pozorovány u několika RNA viry, počítaje v to Chřipka A,[93] Koronavirus[94], Virus mozaiky vojtěšky,[95] a Kachní hepatitida A.[96] Některé viry, jako např HIV-1 a Poliovirus, inhibují buněčný poly-A vazebný protein (PABPC1 ), aby zdůraznili expresi svých vlastních genů nad hostitelskou buňkou.[97]
Dějiny
Poly (A) polymeráza byla poprvé identifikována v roce 1960 jako enzymatická aktivita v extraktech vyrobených z buněčných jader, které by mohly polymerovat ATP, ale ne ADP, na polyadenin.[98][99] Ačkoli byla tato aktivita identifikována v mnoha typech buněk, neměla žádnou známou funkci až do roku 1971, kdy byly v mRNA nalezeny sekvence poly (A).[100][101] Jedinou funkcí těchto sekvencí byla původně myšlenka ochrana 3 'konce RNA před nukleázami, ale později byly identifikovány specifické role polyadenylace v exportu a translaci jader. Polymerázy odpovědné za polyadenylaci byly poprvé purifikovány a charakterizovány v šedesátých a sedmdesátých letech minulého století, ale velké množství pomocných proteinů, které řídí tento proces, bylo objeveno až na počátku 90. let.[100]
Viz také
Reference
- ^ A b Proudfoot NJ, Furger A, Dye MJ (únor 2002). Msgstr "Integrace zpracování mRNA s transkripcí". Buňka. 108 (4): 501–12. doi:10.1016 / S0092-8674 (02) 00617-7. PMID 11909521. S2CID 478260.
- ^ A b Guhaniyogi J, Brewer G (březen 2001). "Regulace stability mRNA v buňkách savců". Gen. 265 (1–2): 11–23. doi:10.1016 / S0378-1119 (01) 00350-X. PMC 3340483. PMID 11255003.
- ^ A b C Richter JD (červen 1999). „Cytoplazmatická polyadenylace ve vývoji i mimo něj“. Recenze mikrobiologie a molekulární biologie. 63 (2): 446–56. doi:10.1128 / MMBR.63.2.446-456.1999. PMC 98972. PMID 10357857.
- ^ Steege DA (srpen 2000). „Nové rysy rozpadu mRNA v bakteriích“. RNA. 6 (8): 1079–90. doi:10.1017 / S1355838200001023. PMC 1369983. PMID 10943888.
- ^ Zhuang Y, Zhang H, Lin S (červen 2013). "Polyadenylace 18S rRNA v řasách (1)". Journal of Phycology. 49 (3): 570–9. doi:10.1111 / jpy.12068. PMID 27007045. S2CID 19863143.
- ^ Anderson JT (srpen 2005). „Obrat RNA: neočekávané důsledky sledování“. Aktuální biologie. 15 (16): R635-8. doi:10.1016 / j.cub.2005.08.002. PMID 16111937. S2CID 19003617.
- ^ A b Sarkar N (červen 1997). "Polyadenylace mRNA u prokaryot". Roční přehled biochemie. 66 (1): 173–97. doi:10,1146 / annurev.biochem. 66.1.173. PMID 9242905.
- ^ Stevens A (1963). "Ribonukleové kyseliny - biosyntéza a degradace". Roční přehled biochemie. 32: 15–42. doi:10.1146 / annurev.bi.32.070163.000311. PMID 14140701.
- ^ Lehninger AL, Nelson DL, Cox MM, eds. (1993). Základy biochemie (2. vyd.). New York: Worth. ISBN 978-0-87901-500-8.[stránka potřebná ]
- ^ Abaza I, Gebauer F (březen 2008). „Obchodní překlad s proteiny vázajícími RNA“. RNA. 14 (3): 404–9. doi:10,1261 / rna.848208. PMC 2248257. PMID 18212021.
- ^ Mattick JS, Makunin IV (duben 2006). „Nekódující RNA“. Lidská molekulární genetika. 15 Spec No 1 (90001): R17-29. doi:10,1093 / hmg / ddl046. PMID 16651366.
- ^ A b Hunt AG, Xu R, Addepalli B, Rao S, Forbes KP, Meeks LR, Xing D, Mo M, Zhao H, Bandyopadhyay A, Dampanaboina L, Marion A, Von Lanken C, Li QQ (květen 2008). „Polyadenylační stroj Arabidopsis mRNA: komplexní analýza interakcí protein-protein a profilování genové exprese“. BMC Genomics. 9: 220. doi:10.1186/1471-2164-9-220. PMC 2391170. PMID 18479511.
- ^ A b Dávila López M, Samuelsson T (leden 2008). „Časný vývoj zpracování histonové mRNA 3“. RNA. 14 (1): 1–10. doi:10,1261 / rna.782308. PMC 2151031. PMID 17998288.
- ^ Marzluff WF, Gongidi P, Woods KR, Jin J, Maltais LJ (listopad 2002). "Lidské a myší replikačně závislé histonové geny". Genomika. 80 (5): 487–98. doi:10.1016 / S0888-7543 (02) 96850-3. PMID 12408966.
- ^ Saini HK, Griffiths-Jones S, Enright AJ (listopad 2007). „Genomická analýza transkriptů lidské mikroRNA“. Sborník Národní akademie věd Spojených států amerických. 104 (45): 17719–24. Bibcode:2007PNAS..10417719S. doi:10.1073 / pnas.0703890104. PMC 2077053. PMID 17965236.
- ^ Yoshikawa M, Peragine A, Park MY, Poethig RS (září 2005). „Cesta k biogenezi trans-působících siRNA v Arabidopsis“. Geny a vývoj. 19 (18): 2164–75. doi:10.1101 / gad.1352605. PMC 1221887. PMID 16131612.
- ^ Amaral PP, Mattick JS (srpen 2008). "Nekódující RNA ve vývoji". Savčí genom. 19 (7–8): 454–92. doi:10.1007 / s00335-008-9136-7. PMID 18839252. S2CID 206956408.
- ^ A b C d Bienroth S, Keller W, Wahle E (únor 1993). „Sestavení polyadenylačního komplexu procesivního messengerového RNA“. Časopis EMBO. 12 (2): 585–94. doi:10.1002 / j.1460-2075.1993.tb05690.x. PMC 413241. PMID 8440247.
- ^ A b Liu D, Brockman JM, Dass B, Hutchins LN, Singh P, McCarrey JR, MacDonald CC, Graber JH (2006). „Systematické variace signálů mRNA 3′-zpracování během spermatogeneze myší“. Výzkum nukleových kyselin. 35 (1): 234–46. doi:10.1093 / nar / gkl919. PMC 1802579. PMID 17158511.
- ^ Lutz CS (říjen 2008). "Alternativní polyadenylace: zkroucení tvorby konce mRNA 3 '". ACS Chemická biologie. 3 (10): 609–17. doi:10.1021 / cb800138w. PMID 18817380.
- ^ A b Beaudoing E, Freier S, Wyatt JR, Claverie JM, Gautheret D (červenec 2000). „Vzory využití variantního polyadenylačního signálu v lidských genech“. Výzkum genomu. 10 (7): 1001–10. doi:10.1101 / gr.10.7.1001. PMC 310884. PMID 10899149.
- ^ Brown KM, GM Gilmartin (prosinec 2003). "Mechanismus pro regulaci zpracování pre-mRNA 3 'lidským štěpným faktorem Im". Molekulární buňka. 12 (6): 1467–76. doi:10.1016 / S1097-2765 (03) 00453-2. PMID 14690600.
- ^ Yang Q, Gilmartin GM, Doublié S (červen 2010). „Strukturální základ rozpoznávání UGUA proteinem Nudix CFI (m) 25 a důsledky pro regulační roli při zpracování mRNA 3 '“. Sborník Národní akademie věd Spojených států amerických. 107 (22): 10062–7. Bibcode:2010PNAS..10710062Y. doi:10.1073 / pnas.1000848107. PMC 2890493. PMID 20479262.
- ^ Yang Q, Coseno M, GM Gilmartin, Doublié S (březen 2011). „Krystalová struktura komplexu lidského štěpného faktoru CFI (m) 25 / CFI (m) 68 / RNA poskytuje pohled na rozpoznávání poly (A) místa a RNA smyčky“. Struktura. 19 (3): 368–77. doi:10.1016 / j.str.2010.12.021. PMC 3056899. PMID 21295486.
- ^ Venkataraman K, Brown KM, Gilmartin GM (červen 2005). „Analýza nekanonického poly (A) místa odhaluje tripartitní mechanismus pro rozpoznávání poly (A) místa obratlovců“. Geny a vývoj. 19 (11): 1315–27. doi:10.1101 / gad.1298605. PMC 1142555. PMID 15937220.
- ^ A b Millevoi S, Loulergue C, Dettwiler S, Karaa SZ, Keller W, Antoniou M, Vagner S (říjen 2006). „Interakce mezi U2AF 65 a CF I (m) spojuje sestřih a stroj na zpracování na 3 ′ konci“. Časopis EMBO. 25 (20): 4854–64. doi:10.1038 / sj.emboj.7601331. PMC 1618107. PMID 17024186.
- ^ A b C Shen Y, Ji G, Haas BJ, Wu X, Zheng J, Reese GJ, Li QQ (květen 2008). „Analýza genomové úrovně signálů zpracování mRNA 3'-konce rýže a alternativní polyadenylace“. Výzkum nukleových kyselin. 36 (9): 3150–61. doi:10.1093 / nar / gkn158. PMC 2396415. PMID 18411206.
- ^ Glover-Cutter K, Kim S, Espinosa J, Bentley DL (leden 2008). „RNA polymeráza II se pozastavuje a asociuje s faktory zpracování pre-mRNA na obou koncích genů“. Přírodní strukturní a molekulární biologie. 15 (1): 71–8. doi:10.1038 / nsmb1352. PMC 2836588. PMID 18157150.
- ^ Molekulární biologie buňky, kapitola 6, „Od DNA k RNA“. 4. vydání. Alberts B, Johnson A, Lewis J a kol. New York: Garland Science; 2002.
- ^ Stumpf G, Domdey H (listopad 1996). "Závislost zpracování kvasinkové pre-mRNA 3'-konce na CFT1: sekvenční homolog savčího AAUAAA vazebného faktoru". Věda. 274 (5292): 1517–20. Bibcode:1996Sci ... 274.1517S. doi:10.1126 / science.274.5292.1517. PMID 8929410. S2CID 34840144.
- ^ Iseli C, Stevenson BJ, de Souza SJ, Samaia HB, Camargo AA, Buetow KH, Strausberg RL, Simpson AJ, Bucher P, Jongeneel CV (červenec 2002). „Heterogenita dlouhého dosahu na 3 'koncích lidských mRNA“. Výzkum genomu. 12 (7): 1068–74. doi:10,1101 / gr. 62002. PMC 186619. PMID 12097343.
- ^ Balbo PB, Bohm A (září 2007). "Mechanism of poly (A) polymerase: structure of the enzym-MgATP-RNA ternary complex and kinetical analysis". Struktura. 15 (9): 1117–31. doi:10.1016 / j.str.2007.07.010. PMC 2032019. PMID 17850751.
- ^ Viphakone N, Voisinet-Hakil F, Minvielle-Sebastia L (duben 2008). "Molekulární disekce kontroly délky ocasu mRNA poly (A) v kvasinkách". Výzkum nukleových kyselin. 36 (7): 2418–33. doi:10.1093 / nar / gkn080. PMC 2367721. PMID 18304944.
- ^ Wahle E (únor 1995). „Řízení délky ocasu Poly (A) je způsobeno ukončením procesivní syntézy“. The Journal of Biological Chemistry. 270 (6): 2800–8. doi:10.1074 / jbc.270.6.2800. PMID 7852352.
- ^ Dichtl B, Blank D, Sadowski M, Hübner W, Weiser S, Keller W (srpen 2002). „Yhh1p / Cft1p přímo spojuje rozpoznávání poly (A) místa a ukončení transkripce RNA polymerázy II“. Časopis EMBO. 21 (15): 4125–35. doi:10.1093 / emboj / cdf390. PMC 126137. PMID 12145212.
- ^ Nag A, Narsinh K, Martinson HG (červenec 2007). „Poly (A) -dependentní transkripční pauza je zprostředkována CPSF působícím na tělo polymerázy“. Přírodní strukturní a molekulární biologie. 14 (7): 662–9. doi:10.1038 / nsmb1253. PMID 17572685. S2CID 5777074.
- ^ Tefferi A, Wieben ED, Dewald GW, Whiteman DA, Bernard ME, Spelsberg TC (srpen 2002). „Primer on medical genomics part II: Background Principles and Methods in molekulární genetiky“. Mayo Clinic Proceedings. 77 (8): 785–808. doi:10.4065/77.8.785. PMID 12173714. S2CID 2237085.
- ^ Coller JM, Gray NK, Wickens MP (říjen 1998). "Stabilizace mRNA poly (A) vazebným proteinem je nezávislá na poly (A) a vyžaduje translaci". Geny a vývoj. 12 (20): 3226–35. doi:10.1101 / gad.12.20.3226. PMC 317214. PMID 9784497.
- ^ A b Siddiqui N, Mangus DA, Chang TC, Palermino JM, Shyu AB, Gehring K (srpen 2007). "Poly (A) nukleáza interaguje s C-koncovou doménou polyadenylát vázající proteinové domény z poly (A) -vázajícího proteinu". The Journal of Biological Chemistry. 282 (34): 25067–75. doi:10,1074 / jbc.M701256200. PMID 17595167.
- ^ Vinciguerra P, Stutz F (červen 2004). "export mRNA: montážní linka z genů do jaderných pórů". Současný názor na buněčnou biologii. 16 (3): 285–92. doi:10.1016 / j.ceb.2004.03.013. PMID 15145353.
- ^ Gray NK, Coller JM, Dickson KS, Wickens M (září 2000). „Více částí proteinu vázajícího poly (A) stimuluje translaci in vivo“. Časopis EMBO. 19 (17): 4723–33. doi:10.1093 / emboj / 19.17.4723. PMC 302064. PMID 10970864.
- ^ Meaux S, Van Hoof A (červenec 2006). „Kvasinkové transkripty štěpené vnitřním ribozymem poskytují nový pohled na roli víčka a poly (A) ocasu v translaci a rozpadu mRNA“. RNA. 12 (7): 1323–37. doi:10,1261 / rna.46306. PMC 1484436. PMID 16714281.
- ^ Kargapolova Y, Levin M, Lackner K, Danckwardt S (červen 2017). „sCLIP - integrovaná platforma ke studiu interakcí RNA-protein v biomedicínském výzkumu: identifikace CSTF2tau v alternativním zpracování malých jaderných RNA“. Výzkum nukleových kyselin. 45 (10): 6074–6086. doi:10.1093 / nar / gkx152. PMC 5449641. PMID 28334977.
- ^ A b Meijer HA, Bushell M, Hill K, Gant TW, Willis AE, Jones P, de Moor CH (2007). „Nová metoda frakcionace poly (A) odhaluje velkou populaci mRNA s krátkým poly (A) ocasem v savčích buňkách“. Výzkum nukleových kyselin. 35 (19): e132. doi:10.1093 / nar / gkm 830. PMC 2095794. PMID 17933768.
- ^ Lehner B, Sanderson CM (červenec 2004). „Rámec interakce proteinů pro degradaci lidské mRNA“. Výzkum genomu. 14 (7): 1315–23. doi:10,1101 / gr. 2122004. PMC 442147. PMID 15231747.
- ^ Wu L, Fan J, Belasco JG (březen 2006). „MicroRNAs direct quick deadenylation of mRNA“. Sborník Národní akademie věd Spojených států amerických. 103 (11): 4034–9. Bibcode:2006PNAS..103,4034W. doi:10.1073 / pnas.0510928103. PMC 1449641. PMID 16495412.
- ^ Cui J, Sackton KL, Horner VL, Kumar KE, Wolfner MF (duben 2008). „Wispy, homolog Drosophila GLD-2, je vyžadován během oogeneze a aktivace vajíčka“. Genetika. 178 (4): 2017–29. doi:10.1534 / genetika.107.084558. PMC 2323793. PMID 18430932.
- ^ Wilusz CJ, Wormington M, Peltz SW (duben 2001). "Průvodce„ cap-to-tail “k obratu mRNA“. Nature Reviews Molecular Cell Biology. 2 (4): 237–46. doi:10.1038/35067025. PMID 11283721. S2CID 9734550.
- ^ Jung MY, Lorenz L, Richter JD (červen 2006). „Translační kontrola neuroguidinem, eukaryotickým iniciačním faktorem 4E a vazebným proteinem CPEB“. Molekulární a buněčná biologie. 26 (11): 4277–87. doi:10.1128 / MCB.02470-05. PMC 1489097. PMID 16705177.
- ^ Sakurai T, Sato M, Kimura M (listopad 2005). "Různorodé vzorce prodloužení ocasu poly (A) a zkrácení myší mateřské mRNA z plně dospělého oocytu do 2-buněčných stadií embrya". Sdělení o biochemickém a biofyzikálním výzkumu. 336 (4): 1181–9. doi:10.1016 / j.bbrc.2005.08.250. PMID 16169522.
- ^ Taft RA (leden 2008). „Cnosti a omezení preimplantačního myšího embrya jako modelového systému“. Theriogenologie. 69 (1): 10–6. doi:10.1016 / j.theriogenology.2007.09.032. PMC 2239213. PMID 18023855.
- ^ Richter JD (červen 2007). "CPEB: život v překladu". Trendy v biochemických vědách. 32 (6): 279–85. doi:10.1016 / j.tibs.2007.04.004. PMID 17481902.
- ^ Piqué M, López JM, Foissac S, Guigó R, Méndez R (únor 2008). "Kombinatorický kód pro překladovou kontrolu zprostředkovanou CPE". Buňka. 132 (3): 434–48. doi:10.1016 / j.cell.2007.12.038. PMID 18267074. S2CID 16092673.
- ^ Benoit P, Papin C, Kwak JE, Wickens M, Simonelig M (červen 2008). „Poly (A) polymerázy typu PAP a GLD-2 jsou vyžadovány postupně u cytoplazmatické polyadenylace a oogeneze u Drosophila“. Rozvoj. 135 (11): 1969–79. doi:10.1242 / dev.021444. PMID 18434412.
- ^ Tian B, Hu J, Zhang H, Lutz CS (2005). „Rozsáhlá analýza polyadenylace mRNA lidských a myších genů“. Výzkum nukleových kyselin. 33 (1): 201–12. doi:10.1093 / nar / gki158. PMC 546146. PMID 15647503.
- ^ Danckwardt S, Hentze MW, Kulozik AE (únor 2008). „Zpracování mRNA na 3 konci: molekulární mechanismy a důsledky pro zdraví a nemoci“. Časopis EMBO. 27 (3): 482–98. doi:10.1038 / sj.emboj.7601932. PMC 2241648. PMID 18256699.
- ^ A b Tian, Bin; Manley, James L. (2017). „Alternativní polyadenylace prekurzorů mRNA“. Recenze přírody. Molekulární buněčná biologie. 18 (1): 18–30. doi:10.1038 / nrm.2016.116. ISSN 1471-0080. PMC 5483950. PMID 27677860.
- ^ Zhang, Haibo; Lee, Ju Youn; Tian, Bin (2005). „Předpojatá alternativní polyadenylace v lidských tkáních“. Genome Biology. 6 (12): R100. doi:10.1186 / gb-2005-6-12-r100. ISSN 1474-760X. PMC 1414089. PMID 16356263.
- ^ Smibert, Peter; Miura, Pedro; Westholm, Jakub O .; Shenker, Sol; May, Gemma; Duff, Michael O .; Zhang, Dayu; Eads, Brian D .; Carlson, Joe; Brown, James B .; Eisman, Robert C. (2012). „Globální vzorce alternativní polyadenylace specifické pro tkáně v Drosophile“. Zprávy buněk. 1 (3): 277–289. doi:10.1016 / j.celrep.2012.01.001. ISSN 2211-1247. PMC 3368434. PMID 22685694.
- ^ Lee, Ju Youn; Ji, Zhe; Tian, Bin (2008). „Fylogenetická analýza polyadenylačních míst mRNA odhaluje roli transponovatelných prvků ve vývoji 3'-konce genů“. Výzkum nukleových kyselin. 36 (17): 5581–5590. doi:10.1093 / nar / gkn540. ISSN 1362-4962. PMC 2553571. PMID 18757892.
- ^ Ogorodnikov A, Kargapolova Y, Danckwardt S (červen 2016). „Zpracování a expanze transkriptomu na konci mRNA 3 've zdraví a nemoci: nalezení správného konce“. Archiv Pflügers. 468 (6): 993–1012. doi:10.1007 / s00424-016-1828-3. PMC 4893057. PMID 27220521.
- ^ Sandberg R, Neilson JR, Sarma A, Sharp PA, Burge CB (červen 2008). „Proliferující buňky exprimují mRNA se zkrácenými 3 'nepřekládanými oblastmi a méně cílovými místy pro mikroRNA“. Věda. 320 (5883): 1643–7. Bibcode:2008Sci ... 320.1643S. doi:10.1126 / science.1155390. PMC 2587246. PMID 18566288.
- ^ Tili E, Michaille JJ, Calin GA (duben 2008). "Exprese a funkce mikro-RNA v imunitních buňkách během normálního nebo chorobného stavu". International Journal of Medical Sciences. 5 (2): 73–9. doi:10,7150 / ijms. 5.73. PMC 2288788. PMID 18392144.
- ^ Ghosh T, Soni K, Scaria V, Halimani M, Bhattacharjee C, Pillai B (listopad 2008). „MicroRNA zprostředkovaná up-regulace alternativně polyadenylované varianty myšího cytoplazmatického {beta} -aktinového genu“. Výzkum nukleových kyselin. 36 (19): 6318–32. doi:10.1093 / nar / gkn624. PMC 2577349. PMID 18835850.
- ^ Alt FW, Bothwell AL, Knapp M, Siden E, Mather E, Koshland M, Baltimore D (červen 1980). „Syntéza secernovaných a na membránu vázaných imunoglobulinových těžkých řetězců mu je řízena mRNA, které se liší na svých 3 'koncích“. Buňka. 20 (2): 293–301. doi:10.1016/0092-8674(80)90615-7. PMID 6771018. S2CID 7448467.
- ^ Tian B, Pan Z, Lee JY (únor 2007). „Rozšířené polyadenylační události mRNA v intronech naznačují dynamickou souhru mezi polyadenylací a sestřihem“. Výzkum genomu. 17 (2): 156–65. doi:10,1101 / gr. 5532707. PMC 1781347. PMID 17210931.
- ^ A b Shell SA, Hesse C, Morris SM, Milcarek C (prosinec 2005). „Zvýšené hladiny stimulačního faktoru štěpení 64-kDa (CstF-64) v makrofágech stimulovaných lipopolysacharidem ovlivňují genovou expresi a indukují alternativní výběr poly (A) místa“. The Journal of Biological Chemistry. 280 (48): 39950–61. doi:10,1074 / jbc.M508848200. PMID 16207706.
- ^ Ogorodnikov A, Levin M, Tattikota S, Tokalov S, Hoque M, Scherzinger D, Marini F, Poetsch A, Binder H, Macher-Göppinger S, Probst HC, Tian B, Schaefer M, Lackner KJ, Westermann F, Danckwardt S ( Prosince 2018). "Organizace transkriptomu 3 'pomocí PCF11 spojuje alternativní polyadenylaci s tvorbou a neuronální diferenciací neuroblastomu". Příroda komunikace. 9 (1): 5331. Bibcode:2018NatCo ... 9.5331O. doi:10.1038 / s41467-018-07580-5. PMC 6294251. PMID 30552333.
- ^ Licatalosi DD, Mele A, Fak JJ, Ule J, Kayikci M, Chi SW, Clark TA, Schweitzer AC, Blume JE, Wang X, Darnell JC, Darnell RB (listopad 2008). „HITS-CLIP přináší genomové poznatky o alternativním zpracování RNA v mozku“. Příroda. 456 (7221): 464–9. Bibcode:2008 Natur.456..464L. doi:10.1038 / nature07488. PMC 2597294. PMID 18978773.
- ^ Hall-Pogar T, Liang S, Haag LK, Lutz CS (červenec 2007). „Specifické trans-působící proteiny interagují s pomocnými polyadenylačními prvky RNA v COX-2 3′-UTR“. RNA. 13 (7): 1103–15. doi:10,1261 / rna.577707. PMC 1894925. PMID 17507659.
- ^ Danckwardt S, Kaufmann I, Gentzel M, Foerstner KU, Gantzert AS, Gehring NH, Neu-Yilik G, Bork P, Keller W, Wilm M, Hentze MW, Kulozik AE (červen 2007). „Faktory sestřihu stimulují polyadenylaci pomocí USE při nekanonických 3 'konci formujících signálů“. Časopis EMBO. 26 (11): 2658–69. doi:10.1038 / sj.emboj.7601699. PMC 1888663. PMID 17464285.
- ^ Danckwardt S, Gantzert AS, Macher-Goeppinger S, Probst HC, Gentzel M, Wilm M, Gröne HJ, Schirmacher P, Hentze MW, Kulozik AE (únor 2011). "p38 MAPK řídí expresi protrombinu regulovaným zpracováním konce 3 'konce RNA". Molekulární buňka. 41 (3): 298–310. doi:10.1016 / j.molcel.2010.12.032. PMID 21292162.
- ^ Wood AJ, Schulz R, Woodfine K, Koltowska K, Beechey CV, Peters J, Bourc'his D, Oakey RJ (květen 2008). „Regulace alternativní polyadenylace pomocí genomového otisku“. Geny a vývoj. 22 (9): 1141–6. doi:10,1101 / gad.473408. PMC 2335310. PMID 18451104.
- ^ Reinisch KM, Wolin SL (duben 2007). "Vznikající témata v nekódující kontrole kvality RNA". Aktuální názor na strukturní biologii. 17 (2): 209–14. doi:10.1016 / j.sbi.2007.03.012. PMID 17395456.
- ^ Jia H, Wang X, Liu F, Guenther UP, Srinivasan S, Anderson JT, Jankowsky E (červen 2011). „RNA helikáza Mtr4p moduluje polyadenylaci v komplexu TRAMP“. Buňka. 145 (6): 890–901. doi:10.1016 / j.cell.2011.05.010. PMC 3115544. PMID 21663793.
- ^ LaCava J, Houseley J, Saveanu C, Petfalski E, Thompson E, Jacquier A, Tollervey D (červen 2005). „Odbourávání RNA exosomem je podporováno nukleárním polyadenylačním komplexem“. Buňka. 121 (5): 713–24. doi:10.1016 / j.cell.2005.04.029. PMID 15935758. S2CID 14898055.
- ^ A b Martin G, Keller W (listopad 2007). "RNA-specifické ribonukleotidyltransferázy". RNA. 13 (11): 1834–49. doi:10,1261 / rna.652807. PMC 2040100. PMID 17872511.
- ^ Slomovic S, Laufer D, Geiger D, Schuster G (2006). "Polyadenylace ribozomální RNA v lidských buňkách". Výzkum nukleových kyselin. 34 (10): 2966–75. doi:10.1093 / nar / gkl357. PMC 1474067. PMID 16738135.
- ^ Régnier P, Arraiano CM (březen 2000). „Degradace mRNA v bakteriích: vznik všudypřítomných znaků“. BioEssays. 22 (3): 235–44. doi:10.1002 / (SICI) 1521-1878 (200003) 22: 3 <235 :: AID-BIES5> 3.0.CO; 2-2. PMID 10684583.
- ^ A b C Anantharaman V, Koonin EV, Aravind L (duben 2002). „Srovnávací genomika a vývoj proteinů podílejících se na metabolismu RNA“. Výzkum nukleových kyselin. 30 (7): 1427–64. doi:10.1093 / nar / 30.7.1427. PMC 101826. PMID 11917006.
- ^ A b Slomovic S, Portnoy V, Liveanu V, Schuster G (2006). „Polyadenylace RNA u prokaryot a organel; různé ocasy vyprávějí různé příběhy“. Kritické recenze v rostlinných vědách. 25: 65–77. doi:10.1080/07352680500391337. S2CID 86607431.
- ^ Chang, Jeong Ho; Tong, Liang (2012). „Mitochondriální poly (A) polymeráza a polyadenylace“. Biochimica et Biophysica Acta (BBA) - regulační mechanismy genů. 1819 (9–10): 992–997. doi:10.1016 / j.bbagrm.2011.10.012. ISSN 0006-3002. PMC 3307840. PMID 22172994.
- ^ Chang SA, Cozad M, Mackie GA, Jones GH (leden 2008). „Kinetika polynukleotid fosforylázy: srovnání enzymů ze Streptomyces a Escherichia coli a účinky nukleosid difosfátů“. Journal of Bacteriology. 190 (1): 98–106. doi:10.1128 / JB.00327-07. PMC 2223728. PMID 17965156.
- ^ Nagaike T, Suzuki T, Ueda T (duben 2008). „Polyadenylace v savčích mitochondriích: poznatky z nedávných studií“. Biochimica et Biophysica Acta (BBA) - regulační mechanismy genů. 1779 (4): 266–9. doi:10.1016 / j.bbagrm.2008.02.001. PMID 18312863.
- ^ Walter M, Kilian J, Kudla J (prosinec 2002). „Aktivita PNPázy určuje účinnost zpracování mRNA 3'-end, degradaci tRNA a rozsah polyadenylace v chloroplastech“. Časopis EMBO. 21 (24): 6905–14. doi:10.1093 / emboj / cdf686. PMC 139106. PMID 12486011.
- ^ Portnoy V, Schuster G (2006). „Polyadenylace a degradace RNA v různých Archaeách; role exosomu a RNázy R“. Výzkum nukleových kyselin. 34 (20): 5923–31. doi:10.1093 / nar / gkl763. PMC 1635327. PMID 17065466.
- ^ Yehudai-Resheff S, Portnoy V, Yogev S, Adir N, Schuster G (září 2003). „Analýza domény chloroplastové polynukleotid fosforylázy odhaluje diskrétní funkce při degradaci RNA, polyadenylaci a sekvenční homologii s exosomovými proteiny“. Rostlinná buňka. 15 (9): 2003–19. doi:10.1105 / tpc.013326. PMC 181327. PMID 12953107.
- ^ Slomovic S, Portnoy V, Schuster G (2008). RNA Turnover in Prokaryotes, Archaea and Organelles: Chapter 24 Detection and Characterization of Polyadenylated RNA in Eukarya, Bacteria, Archaea, and Organelles. Metody v enzymologii. 447. pp. 501–20. doi:10.1016/S0076-6879(08)02224-6. ISBN 978-0-12-374377-0. PMID 19161858.
- ^ Portnoy V, Evguenieva-Hackenberg E, Klein F, Walter P, Lorentzen E, Klug G, Schuster G (December 2005). "RNA polyadenylation in Archaea: not observed in Haloferax while the exosome polynucleotidylates RNA in Sulfolobus". Zprávy EMBO. 6 (12): 1188–93. doi:10.1038/sj.embor.7400571. PMC 1369208. PMID 16282984.
- ^ Portnoy V, Schuster G (June 2008). "Mycoplasma gallisepticum as the first analyzed bacterium in which RNA is not polyadenylated". Mikrobiologické dopisy FEMS. 283 (1): 97–103. doi:10.1111/j.1574-6968.2008.01157.x. PMID 18399989.
- ^ Evguenieva-Hackenberg E, Roppelt V, Finsterseifer P, Klug G (December 2008). "Rrp4 and Csl4 are needed for efficient degradation but not for polyadenylation of synthetic and natural RNA by the archaeal exosome". Biochemie. 47 (50): 13158–68. doi:10.1021/bi8012214. PMID 19053279.
- ^ A b Slomovic S, Portnoy V, Yehudai-Resheff S, Bronshtein E, Schuster G (April 2008). "Polynucleotide phosphorylase and the archaeal exosome as poly(A)-polymerases". Biochimica et Biophysica Acta (BBA) - regulační mechanismy genů. 1779 (4): 247–55. doi:10.1016/j.bbagrm.2007.12.004. PMID 18177749.
- ^ Poon, Leo L. M.; Pritlove, David C.; Fodor, Ervin; Brownlee, George G. (1 April 1999). "Direct Evidence that the Poly(A) Tail of Influenza A Virus mRNA Is Synthesized by Reiterative Copying of a U Track in the Virion RNA Template". Journal of Virology. 73 (4): 3473–3476. doi:10.1128/JVI.73.4.3473-3476.1999.
- ^ Wu, Hung-Yi; Ke, Ting-Yung; Liao, Wei-Yu; Chang, Nai-Yun (2013). "Regulation of Coronaviral Poly(A) Tail Length during Infection". PLOS ONE. 8 (7): e70548. Bibcode:2013PLoSO...870548W. doi:10.1371/journal.pone.0070548. PMC 3726627. PMID 23923003.
- ^ Neeleman, Lyda; Olsthoorn, René C. L.; Linthorst, Huub J. M.; Bol, John F. (4 December 2001). "Translation of a nonpolyadenylated viral RNA is enhanced by binding of viral coat protein or polyadenylation of the RNA". Sborník Národní akademie věd. 98 (25): 14286–14291. Bibcode:2001PNAS...9814286N. doi:10.1073/pnas.251542798. PMC 64674. PMID 11717411.
- ^ Chen, Jun-Hao; Zhang, Rui-Hua; Lin, Shao-Li; Li, Peng-Fei; Lan, Jing-Jing; Song, Sha-Sha; Gao, Ji-Ming; Wang, Yu; Xie, Zhi-Jing; Li, Fu-Chang; Jiang, Shi-Jin (2018). "The Functional Role of the 3′ Untranslated Region and Poly(A) Tail of Duck Hepatitis a Virus Type 1 in Viral Replication and Regulation of IRES-Mediated Translation". Hranice v mikrobiologii. 9: 2250. doi:10.3389/fmicb.2018.02250. PMC 6167517. PMID 30319572.
- ^ https://viralzone.expasy.org/909?outline=all_by_species
- ^ Edmonds M, Abrams R (April 1960). "Polynucleotide biosynthesis: formation of a sequence of adenylate units from adenosine triphosphate by an enzyme from thymus nuclei". The Journal of Biological Chemistry. 235 (4): 1142–9. PMID 13819354.
- ^ Colgan DF, Manley JL (November 1997). "Mechanism and regulation of mRNA polyadenylation". Geny a vývoj. 11 (21): 2755–66. doi:10.1101/gad.11.21.2755. PMID 9353246.
- ^ A b Edmonds, M (2002). A history of poly A sequences: from formation to factors to function. Pokrok ve výzkumu nukleových kyselin a molekulární biologie. 71. pp. 285–389. doi:10.1016/S0079-6603(02)71046-5. ISBN 978-0-12-540071-8. PMID 12102557.
- ^ Edmonds M, Vaughan MH, Nakazato H (June 1971). "Polyadenylic acid sequences in the heterogeneous nuclear RNA and rapidly-labeled polyribosomal RNA of HeLa cells: possible evidence for a precursor relationship". Sborník Národní akademie věd Spojených států amerických. 68 (6): 1336–40. Bibcode:1971PNAS...68.1336E. doi:10.1073/pnas.68.6.1336. PMC 389184. PMID 5288383.
Další čtení
- Danckwardt S, Hentze MW, Kulozik AE (February 2008). "3′ end mRNA processing: molecular mechanisms and implications for health and disease". Časopis EMBO. 27 (3): 482–98. doi:10.1038/sj.emboj.7601932. PMC 2241648. PMID 18256699.
externí odkazy
 Média související s Polyadenylace na Wikimedia Commons
Média související s Polyadenylace na Wikimedia Commons