ALOX12 - ALOX12
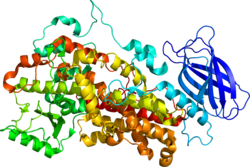




ALOX12 (ES 1.13.11.31 ), také známý jako arachidonát 12-lipoxygenáza, 12-lipoxygenáza, 12S-Lipoxygenáza, 12-LOX, a 12S-LOX je lipoxygenáza -typ enzym že u lidí je kódován ALOX12 gen který se nachází spolu s dalšími lipoyxgenázami na chromozomu 17p13.3.[5][6] ALOX12 je 75 kilodalton protein složený z 663 aminokyselin.
Nomenklatura
| arachidonát 12-lipoxygenáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 1.13.11.31 | ||||||||
| Číslo CAS | 82391-43-3 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
jiný systematické názvy pro ALOX12 zahrnují 12S-lipoxygenázu, destičkovou 12-lipoxygenázu, arachidonát: kyslík 12-oxidoreduktázu, Delta12-lipoxygenázu, 12 Delta-lipoxygenázu a C-12 lipoxygenázu. ALOX12, často označovaný jako 12-lipoxygenáza destičkových destiček, se odlišuje od 12-lipoxygenázy leukocytového typu, která se vyskytuje u myší, potkanů, krav a prasat, ale ne u lidí. 12-lipoxygenáza leukocytového typu u těchto druhů zvířat sdílí 73-86% aminokyselinovou identitu s člověkem ALOX15 ale pouze 57-66% identita s lidskou destičkovou 12-lipoxygenázou a podobně jako ALOX15 metabolizuje kyselinu arachidonovou primárně na 15 (S) -hydroperoxy-5Z,8Z,11Z,13Ekyselina eikosatetraenová (tj. 15 (S) -HpETE; vidět Kyselina 15-hydroxyeikosatetraenová ).[7] V souladu s tím je hlodavčí leukocyt 12-lipoxygenáza považován za ortolog ALOX15 a je označen jako Alox15.[8]
Lidský ALOX12 a ALOX15 spolu s leukocytovým typem hlodavců Alox12 a Alox15 se běžně nazývají 12/15-lipoxygenázy na základě jejich schopnosti metabolizovat kyselinu arachidonovou na obě 12 (S) -HpETE a 15 (S) -HpETE a provádět stejný metabolismus na kyselině arachidonové, která je esterifikovaný na membránu fosfolipidy; lidský ALOX15B dělá 15 (S) -HpETE, ale ne 12 (S) -HpETE, a proto se nepovažuje za 12/15-lipoxygenázu.[9] Studie o roli ALOX12 v patofyziologii využívající hlavní modely těchto funkčních studií, potkanů a myší, jsou komplikované, protože žádný z těchto druhů nemá lipoxygenáza což činí převahu 12 (S) -HETE, a proto je metabolicky ekvivalentní ALOX12.[7][9] Například odvozené funkce Alox12 u myší s nedostatkem Alox12 použitím knokaut metody nemusí označovat podobnou funkci pro ALOX12 u lidí kvůli rozdílům v metabolických aktivitách těchto dvou enzymů. Funkce ALOX12 je dále zakalena lidským ALOX15, který primárně metabolizuje kyselinu arachidonovou na 15 (S) -HpETE, ale také dělá menší, ale stále významná množství 12 (S) -HpETE (viz ALOX15 ).
ALOX12 se také odlišuje od arachidonátu 12-lipoxygenázy typu 12R (ALOX12B), který metabolizuje kyselinu arachidonovou na R stereoizomer z 12 (S) -HpETE viz., 12 (R) -hydroperoxy-5Z,8Z,10E,14Z-ikosatetraenová kyselina (12 (R) -HpETE), produkt s velmi odlišnými patofyziologickými rolemi než produkt 12 (S) -HpETE (viz ALOX12B ).
Objev
ALOX12, původně nazývaný arachidonát 12-lipoxygenáza, byl poprvé charakterizován laureátem Nobelovy ceny, Bengt I. Samuelsson a jeho slavný kolega Mats Hamberg v roce 1974 tím, že ukázal toho člověka krevní destičky metabolizovat kyselina arachidonová nejen známým cyklooxygenáza cesta do prostaglandiny a Kyselina 12-hydroxyheptadekatrienová ale také cestou nezávislou na cyklooxygenáze k 12 (S) -hydroperoxy-5,8,10,14-eikosatetraenová kyselina; tato aktivita byla první savčí lipoxygenázovou aktivitou, která byla charakterizována.[10] V roce 1975 byla k tomuto metabolitu připojena první biologická aktivita ve studiích, které prokázaly, že simuloval chemotaxe člověka neutrofily.[11] Během několika následujících let byl lidský ALOX12 čištěn, charakterizován biochemicky a měl svůj gen molekulárně naklonovaný.[7][12]
Distribuce tkání
Založeno převážně na přítomnosti jeho mRNA, lidský ALOX12 je distribuován převážně v krevních destičkách a leukocyty a na nižších úrovních v bazální vrstvě epidermis (zejména v kožních lézích psoriáza ), Langerhansových ostrůvků uvnitř slinivky břišní a některých druhů rakoviny.[13]
Enzymové aktivity
Zdá se, že řízení aktivity ALOX12 spočívá hlavně na jeho dostupnosti polynenasycená mastná kyselina (PUFA) substráty, které se uvolňují z membrány fosfolipidy buněčnou stimulací.[14] Enzym se účastní metabolismus kyseliny arachidonové provedením následujícího chemická reakce kde jeho substráty jsou kyselina arachidonová (také označovaný jako arachidonát nebo chemicky jako 5Z,8Z,11Z,14Zkyselina eikosatetraenová) a O2 (tj. kyslík ) a jeho produkt je 12S-hydroperoxy-5Z,8Z,10E,14Z-eikosatetraenová kyselina (tj. 12S-hydroperoxyeikosatetraenová kyselina nebo 12S-HpETE):[10][15]
- arachidonát + O.2 → 12S-hydroperoxy-5Z,8Z,10E,14Z-eikosatetraenová kyselina
V buňkách, 12SHpETE může být dále metabolizován samotným ALOX12 ALOXE3 nebo případně jiné, dosud neidentifikované, hepoxilin syntázy do hepoxilin A3 (8R / S-hydroxy-11,12-oxido-5Z,9E,14Zkyselina eikosatrienová) a B3 (10R / S-hydroxy-11,12-oxido-5Z,8Z,14Z-eikosatrienová kyselina):[16][17][18]
- 12S-hydroperoxy-5Z, 8Z, 10E, 14Z-eikosatetraenová kyselina → 8R / S-hydroxy-11,12-oxido-5Z,9E,14Z-eikosatrienová kyselina + 10R / S-hydroxy-11,12-oxido-5Z,8Z,14Z-eikosatrienová kyselina
Hepoxiliny mohou podporovat určité zánět reakce, zvyšují vnímání bolesti (tj. hmatové alodynie ), regulují regionální průtok krve a přispívají k regulaci krevního tlaku na zvířecích modelech (viz Hepoxiliny ). Mnohem častěji však 12S-HpETE se rychle redukuje na své hydroxyl produkt všudypřítomnými aktivitami buněčné peroxidázy, čímž se tvoří 12S-hydroxy-5Z,8Z,10E,14Z-eikosatetraenová kyselina, tj. Kyselina 12-hydroxyeikosatetraenová nebo 12S-HETE:[19]
- 12S-hydroperoxy-5 (Z),8(Z),10(E),14(Z) -eikosatetraenová kyselina → 12S-hydroperoxy-5 (Z),8(Z),10(E),14(Z) -eikosatetraenová kyselina
12S-HETE podporuje zánětlivé reakce, může se podílet na vnímání puritida (tj. svědění) v kůži a reguluje regionální průtok krve ve zvířecích modelech; také podporuje maligní chování kultivovaných lidských rakovinných buněk a také růst některých druhů rakoviny na zvířecích modelech (viz 12-HETE ). Zatímco arachidonát a 12 (S) -HETE jsou převládajícími substráty a produkty ALOX12, enzym také metabolizuje další PUFA. Metabolizuje omega-3 mastné kyseliny, kyselina dokosahexaenová (DHA tj. 4 (Z),7(Z),10(Z),13(Z),16(Z),19(Z) -docosahexaenoic acid to 14 (R) -hydroperoxy-4 (Z),8(Z),10(Z),12(E),16(Z),19(Z) -dokosahexaenová kyselina) (tj. 17-hydroperoxy-DHA); potom může ALOX12 nebo neidentifikovaný enzym typu epoxidázy metabolizovat tento meziprodukt na epoxid, 13,14-epoxy-4 (Z),7(Z),9(E),11(E),16(Z),19(Z) -dokosahexaenová kyselina (tj. 13,14-e-maresin), která se metabolizovala na 7R,14S-dihydroxy-4Z,8E,10E,12Z,16Z,19Zkyselina dokosahexaenová (tj. Maresin 1), neidentifikovaným epoxid hydroláza enzym typu:
- DHA → 17-hydroperoxy-DHA → 13,14-e-maresin → Maresin-1
Maresin 1 má řadu aktivit, které mohou odporovat těm z 12 (S) -HETE a hepoxiliny; je členem třídy označovaných metabolitů PUFA Specializovaní mediátoři pro rozlišení (SPM), které mají protizánětlivé, bolest zmírňující a další obranné aktivity.[20] ALOX12 také působí na leukotrien A4 (LTA4) ve dvou buněčné reakci nazývané transcelulární metabolismus: lidské neutrofily metabolizují kyselinu arachidonovou na její 5,6-epoxid, LTA4, a uvolňují tento meziprodukt do blízkých neutrofilů, které ji metabolizují na lipoxin A4 (5S,6R,15S-trihydroxy-7E,9E,11Z,13Zkyselina eikosatetraenová) a lipoxin B4 (5S,14R,15S-trihydroxy-6E,8Z,10E,12E-eikosatetraenová kyselina); oba lipoxiny jsou SPM s mnoha aktivitami podobnými SPM (viz lipoxin ).[21] ALOX12 může také metabolizovat menší množství DHA na sekundární produkty včetně 17-hydroperoxy-DHA, 11-hydroperoxy-DHA a 8,14-dihydroxy-DHA[20] ALOX12 může také metabolizovat 5 (S) -HETE do 5S,12S-dihydroxyeikosatetraenová kyselina (12,15-diHETE) a 15S-HETE do 14,15S-diETE.[14] I když tyto sloučeniny nebyly důkladně hodnoceny z hlediska biologické aktivity, 17-hydroperoxy-HDHA a redukovaný produkt, na který se rychle převádí v buňkách, 17-hydroxy-HDHA, bylo prokázáno, že inhibují růst kultivovaných lidských buněk rakoviny prostaty tím, že způsobují je vstoupit apoptóza.[22]
Studie na zvířatech
Studie na hlodavcích, kteří postrádají nebo mají deficit v 12-lipoxygenáze typu leukocytů, Alox12 (který nejvíce souvisí s lidským ALOX15) implikuje tento enzym v: A) prevence rozvoje a komplikací vyvolaných stravou a / nebo geneticky cukrovka, tukové dysfunkce buněk / tkání a obezita; b) vývoj ateroskleróza a Steatohepatitida; b) regulace kontrakce krevních cév, dilatace, tlaku, remodelace a angiogeneze; C) udržování normální funkce ledvin, neurologů a mozku; a d) vývoj Alzheimerova choroba.[8][9][23] V těchto studiích je obvykle nejasné, který z nich, pokud vůbec nějaké metabolity Alox12 byl zapleten.
Předklinické studie
Metabolický syndrom
The metabolický syndrom je shluk alespoň tří z pěti následujících zdravotních stavů: břišní (centrální) obezita, zvýšený krevní tlak, zvýšená plazmatická glukóza nalačno (nebo zjevně cukrovka ), vysoké triglyceridy v séru a nízko lipoprotein s vysokou hustotou Úrovně (HDL). ALOX12 a jeho metabolit, 12 (S) -HETE, jsou zvýšené v Langerhansových ostrůvků pacientů s cukrovka 1. typu nebo cukrovka typu 2 stejně jako v tukových buňkách bílá tuková tkáň morbidně obézních pacientů s diabetem typu 2.[8] The PP buňky (tj. gama buňky) ostrůvků pankreatu se jeví jako hlavní, ne-li pouze místo, kde je u těchto pacientů exprimován ALOX12.[8] Studie naznačují, že na ostrůvcích Langerhans ALOX12 a jeho 12 (S) -HETE produkt způsobuje nadměrnou produkci reaktivní formy kyslíku a záněty, které vedou ke ztrátám v inzulín - vylučování beta buňky a tím cukrovku typu 1 a 2 a že v tukové tkáni je nadbytek AlOX12, 12 (S) -HETE, reaktivní formy kyslíku a zánět vedou k dysfunkci tukových buněk (viz také 12-HETE # Zánět a zánětlivá onemocnění a 12-HETE # Diabetes ). Ve skutečnosti v jedné studii a Jednonukleotidový polymorfismus, rs2073438,[24] nachází se v intron oblast ALOX12 Gen byl významně spojen s celkovou a procentuální tukovou hmotou obézních ve srovnání s neobézními mladými čínskými muži.[8][13][18] ALOX12 a 12 (S) -HETE se rovněž podílejí na esenciální hypertenzi (viz další část). Proto může ALOX12 a jeho metabolit (y) přispívat k rozvoji a / nebo progresi obezity, cukrovky, hypertenze a / nebo metabolického syndromu.
Cévy
Selektivní, ale ne zcela specifický inhibitor ALOX12 snížil růstovou reakci kultivovaného člověka endoteliální buňky na základní růstový faktor fibroblastů a vaskulární endoteliální růstový faktor (VEGF); toto snížení bylo částečně obráceno o 12 (S) -HETE; 12 (S) -HETE také stimuluje buněčné linie lidské prostaty k produkci VEGF.[19] Tyto výsledky naznačují, že růstové reakce na dva růstové faktory zahrnují jejich stimulaci 12 (S) -HETE produkce endotelovými buňkami, a proto, že ALOX12 může být cílem pro snížení neo-vaskularizace, která podporuje artritická a rakovinová onemocnění. 12 (S) -HETE také rozšiřuje lidské koronární mikrocirkulační tepny aktivací hladkého svalstva těchto cév BKca Draselné kanály a proto se navrhuje, aby byl Hyperpolarizační faktor odvozený z endotelu.[9][19] Nakonec a varianta s jedním nukleotidem v ALOX12 gen (R261Q [3957 G> A]) byl spojován s esenciální hypertenze a zvýšení vylučování močí o 12 (S) -HETE u lidí a může být faktorem přispívajícím k esenciální hypertenzi (viz také 12-HETE # Krevní tlak ).[9][25]
Alzheimerova choroba
Pacienti s Alzheimerova choroba nebo jiné formy demence mají významně vyšší hladiny 12 (S) -HETE (a 15 (S) -HETE) v mozkomíšním moku ve srovnání s normálními jedinci stejného věku. Doplňkové studie na modelech hlodavců nesoucích lidské mutované geny pro Amyloidní prekurzorový protein a / nebo tau protein (viz tau protein # Klinický význam ), které produkují syndromy podobné Alzheimerově demenci, implikují 12 (S) -HETE, 15 (S) -HETE a enzym typu 12/15-lipoxygenázy ve vývoji a progresi Alzhiemerových chorob podobných příznaků a nálezů u těchto zvířat.[23] V jedné studii bylo zjištěno, že ALOX12 mRNA je zvýšená v mozkové tkáni Alzheimerova choroba pacientů ve srovnání s kontrolními pacienty.[13] Tyto výsledky naznačují, že ALOX12 (nebo ALOX15) může přispívat k rozvoji Alzheimerovy choroby u lidí.
Rakovina
Studie rakoviny prostaty zjistily, že buněčné linie lidské rakoviny prostaty v kultuře nadměrně exprimují ALOX12, nadprodukují 12 (S) -HETE a odpovězte na 12 (S) -HETE zvýšením jejich rychlosti proliferace, zvýšením jejich exprese na buněčném povrchu integriny, což zvyšuje jejich přežití a oddaluje jejich apoptóza a zvýšení jejich produkce vaskulární endoteliální růstový faktor a MMP9 (tj. matricová metalopeptidáza 9); selektivní (ale ne zcela) specifické inhibitory ALOX12 snižovaly proliferaci a přežití těchto buněk (viz také 12-HETE # rakovina prostaty ). Tato zjištění naznačují, že ALOX12 a jeho 12 (S) -HETE produkt může přispívat k růstu a šíření rakoviny prostaty u lidí.[19] V poslední době hypermethylace ALOX12 Gen v tkáni rakoviny prostaty byl spojen s klinickými prediktory pro vysokou míru recidivujícího onemocnění.[26] Některé studie zjistily, že 12 (S) -HETE také podporuje růst a / nebo související pro-maligní chování různých jiných typů kultivovaných rakovinných buněčných linií (viz 12-HETE # Jiné druhy rakoviny ).[19] ALOX12 bylo prokázáno komunikovat s Keratin 5 a LMNA jak je testováno v kvasinkové dvouhybridní interakční knihovně z lidského epidermoidního karcinomu Buňky A431; tyto proteiny jsou kandidáty na regulaci 12-LOX, zejména v nádorových buňkách.[27]
Funkce krevních destiček
Ačkoli byla nejprve identifikována v lidských krevních destičkách, role ALOX12 a jeho hlavních metabolitů, 12 (S) -HpETE a 12 (S) -HETE ve funkci krevních destiček zůstává kontroverzní a nejasný; je možné, že ALOX12-12 (S) -HETE metabolická cesta má dvojí funkce při podpoře nebo inhibici reakcí krevních destiček v závislosti na stimulačním činidle a studované odpovědi, ale inhibice ALOX12 se nakonec může ukázat jako užitečná při inhibici srážení krve související s krevními destičkami.[19]
Další sdružení
The ALOX12 gen má alely citlivosti (rs6502997,[28] rs312462,[29] rs6502998,[30] a rs434473[31]) pro parazitární onemocnění, lidské vrozené toxoplazmóza.[13][32] Plod nositel těchto alel tak trpí zvýšenou náchylností k této nemoci.
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000108839 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000000320 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Funk CD, Funk LB, FitzGerald GA, Samuelsson B (květen 1992). „Charakterizace lidských genů pro 12-lipoxygenázu“. Sborník Národní akademie věd Spojených států amerických. 89 (9): 3962–6. Bibcode:1992PNAS ... 89.3962F. doi:10.1073 / pnas.89.9.3962. PMC 525611. PMID 1570320.
- ^ „Entrez Gene: ALOX12 arachidonát 12-lipoxygenáza“.
- ^ A b C Yamamoto S, Suzuki H, Ueda N (březen 1997). "Arachidonát 12-lipoxygenázy". Pokrok ve výzkumu lipidů. 36 (1): 23–41. doi:10.1016 / s0163-7827 (97) 00002-7. PMID 9373619.
- ^ A b C d E Tersey SA, Bolanis E, Holman TR, Maloney DJ, Nadler JL, Mirmira RG (červen 2015). „Minireview: 12-lipoxygenáza a dysfunkce ostrůvků β-buněk u diabetu“. Molekulární endokrinologie. 29 (6): 791–800. doi:10.1210 / me.2015-1041. PMC 4447641. PMID 25803446.
- ^ A b C d E Dobrian AD, Lieb DC, Cole BK, Taylor-Fishwick DA, Chakrabarti SK, Nadler JL (leden 2011). „Funkční a patologické role 12- a 15-lipoxygenáz“. Pokrok ve výzkumu lipidů. 50 (1): 115–31. doi:10.1016 / j.plipres.2010.10.005. PMC 3012140. PMID 20970452.
- ^ A b Hamberg M, Samuelsson B (září 1974). "Prostaglandinové endoperoxidy. Nové transformace kyseliny arachidonové v lidských krevních destičkách". Sborník Národní akademie věd Spojených států amerických. 71 (9): 3400–4. Bibcode:1974PNAS ... 71.3400H. doi:10.1073 / pnas.71.9.3400. PMC 433780. PMID 4215079.
- ^ Turner SR, Campbell JA, Lynn WS (červen 1975). „Polymorfonukleární chemotaxe leukocytů vůči oxidovaným lipidovým složkám buněčných membrán“. The Journal of Experimental Medicine. 141 (6): 1437–41. doi:10.1084 / jem.141.6.1437. PMC 2189855. PMID 1127383.
- ^ Yoshimoto T, Yamamoto Y, Arakawa T, Suzuki H, Yamamoto S, Yokoyama C, Tanabe T, Toh H (listopad 1990). "Molekulární klonování a exprese lidského arachidonátu 12-lipoxygenázy". Sdělení o biochemickém a biofyzikálním výzkumu. 172 (3): 1230–5. doi:10.1016 / 0006-291X (90) 91580-L. PMID 2244907.
- ^ A b C d Kuhn H, Banthiya S, van Leyen K (duben 2015). „Savčí lipoxygenázy a jejich biologický význam“. Biochimica et Biophysica Acta (BBA) - molekulární a buněčná biologie lipidů. 1851 (4): 308–30. doi:10.1016 / j.bbalip.2014.10.002. PMC 4370320. PMID 25316652.
- ^ A b Yeung J, Holinstat M (červenec 2011). „12-lipoxygenáza: potenciální cíl pro nová protidestičková terapeutika“. Kardiovaskulární a hematologické látky v léčivé chemii. 9 (3): 154–64. doi:10.2174/187152511797037619. PMC 3171607. PMID 21838667.
- ^ Nugteren DH (únor 1975). "Arachidonát lipoxygenáza v krevních destičkách". Biochimica et Biophysica Acta (BBA) - lipidy a metabolismus lipidů. 380 (2): 299–307. doi:10.1016/0005-2760(75)90016-8. PMID 804329.
- ^ Krieg P, Fürstenberger G (březen 2014). "Úloha lipoxygenáz v pokožce". Biochimica et Biophysica Acta (BBA) - molekulární a buněčná biologie lipidů. 1841 (3): 390–400. doi:10.1016 / j.bbalip.2013.08.005. PMID 23954555.
- ^ Muñoz-Garcia A, Thomas CP, Keeney DS, Zheng Y, Brash AR (březen 2014). „Důležitost dráhy lipoxygenázy a hepoxilinu v epidermální bariéře savců“. Biochimica et Biophysica Acta (BBA) - molekulární a buněčná biologie lipidů. 1841 (3): 401–8. doi:10.1016 / j.bbalip.2013.08.020. PMC 4116325. PMID 24021977.
- ^ A b Pace-Asciak CR (duben 2015). "Patofyziologie hepoxilinů". Biochimica et Biophysica Acta (BBA) - molekulární a buněčná biologie lipidů. 1851 (4): 383–96. doi:10.1016 / j.bbalip.2014.09.007. PMID 25240838.
- ^ A b C d E F Porro B, Songia P, Squellerio I, Tremoli E, Cavalca V (srpen 2014). „Analýza, fyziologický a klinický význam 12-HETE: zanedbaný produkt 12-lipoxygenázy pocházející z krevních destiček“. Journal of Chromatography B. 964: 26–40. doi:10.1016 / j.jchromb.2014.03.015. PMID 24685839.
- ^ A b Weylandt KH (listopad 2015). „Metabolity a mediátory odvozené od kyseliny dokosapentaenové - nový svět medicíny lipidových mediátorů v kostce“. European Journal of Pharmacology. 785: 108–115. doi:10.1016 / j.ejphar.2015.11.002. PMID 26546723.
- ^ Serhan CN (2005). „Lipoxiny a 15-epi-lipoxiny spouštěné aspirinem jsou prvními lipidovými mediátory endogenního protizánětlivého účinku a rozlišení.“ Prostaglandiny, leukotrieny a esenciální mastné kyseliny. 73 (3–4): 141–62. doi:10.1016 / j.plefa.2005.05.002. PMID 16005201.
- ^ O'Flaherty JT, Hu Y, Wooten RE, Horita DA, Samuel MP, Thomas MJ, Sun H, Edwards IJ (2012). „Metabolity 15-lipoxygenázy kyseliny dokosahexaenové inhibují proliferaci a přežití buněk rakoviny prostaty“. PLOS ONE. 7 (9): e45480. Bibcode:2012PLoSO ... 745480O. doi:10.1371 / journal.pone.0045480. PMC 3447860. PMID 23029040.
- ^ A b Joshi YB, Giannopoulos PF, Praticò D (březen 2015). „12/15-lipoxygenáza jako objevující se terapeutický cíl pro Alzheimerovu chorobu“. Trendy ve farmakologických vědách. 36 (3): 181–6. doi:10.1016 / j.tips.2015.01.005. PMC 4355395. PMID 25708815.
- ^ „rs2073438“. NCBI dbSNP.
- ^ Quintana LF, Guzmán B, Collado S, Clària J, Poch E (únor 2006). „Kódující polymorfismus v genu 12-lipoxygenázy je spojen s esenciální hypertenzí a močovým 12 (S) -HETE“. Ledviny International. 69 (3): 526–30. doi:10.1038 / sj.ki.5000147. PMID 16514435.
- ^ Angulo JC, Lopez JI, Dorado JF, Sanchez-Chapado M, Colas B, Ropero S (2016). „Hypermethylační profil DNA nezávisle předpovídá biochemickou recidivu po radikální prostatektomii“. Urologia Internationalis. 97 (1): 16–25. doi:10.1159/000446446. PMID 27220660. S2CID 12659175.
- ^ Tang K, Finley RL, Nie D, Honn KV (březen 2000). "Identifikace interakce 12-lipoxygenázy s buněčnými proteiny kvasinkovým dvouhybridním screeningem". Biochemie. 39 (12): 3185–91. doi:10.1021 / bi992664v. PMID 10727209.
- ^ „rs6502997“. NCBI dbSNP.
- ^ „rs312462“. NCBI dbSNP.
- ^ „rs6502998“. NCBI dbSNP.
- ^ „rs434473“. NCBI dbSNP.
- ^ Witola WH, Liu SR, Montpetit A, Welti R, Hypolite M, Roth M, Zhou Y, Mui E, Cesbron-Delauw MF, Fournie GJ, Cavailles P, Bisanz C, Boyer K, Withers S, Noble AG, Swisher CN, Heydemann PT, Rabiah P, Muench SP, McLeod R (červenec 2014). „ALOX12 v lidské toxoplazmóze“. Infekce a imunita. 82 (7): 2670–9. doi:10.1128 / IAI.01505-13. PMC 4097613. PMID 24686056.
externí odkazy
- Člověk ALOX12 umístění genomu a ALOX12 stránka s podrobnostmi o genu v UCSC Genome Browser.
Další čtení
- Yoshimoto T, Arakawa T, Hada T, Yamamoto S, Takahashi E (prosinec 1992). "Struktura a chromozomální lokalizace genu pro lidskou arachidonátovou 12-lipoxygenázu". The Journal of Biological Chemistry. 267 (34): 24805–9. PMID 1447217.
- Izumi T, Hoshiko S, Rådmark O, Samuelsson B (říjen 1990). "Klonování cDNA pro lidskou 12-lipoxygenázu". Sborník Národní akademie věd Spojených států amerických. 87 (19): 7477–81. Bibcode:1990PNAS ... 87.7477I. doi:10.1073 / pnas.87.19.7477. PMC 54770. PMID 2217179.
- Funk CD, Furci L, FitzGerald GA (srpen 1990). "Molekulární klonování, primární struktura a exprese 12-lipoxygenázy z lidských destiček / erytroleukemických buněk". Sborník Národní akademie věd Spojených států amerických. 87 (15): 5638–42. Bibcode:1990PNAS ... 87.5638F. doi:10.1073 / pnas.87.15.5638. PMC 54382. PMID 2377602.
- Flatman S, Morgan A, McDonald-Gibson RG, Davey A, Jonas GE, Slater TF (květen 1988). "Aktivita 12-lipoxygenázy v děložním čípku člověka". Prostaglandiny, leukotrieny a esenciální mastné kyseliny. 32 (2): 87–94. doi:10.1016/0952-3278(88)90101-9. PMID 3406043.
- Wong PY, Westlund P, Hamberg M, Granström E, Chao PH, Samuelsson B (srpen 1985). "15-Lipoxygenáza v lidských krevních destičkách". The Journal of Biological Chemistry. 260 (16): 9162–5. PMID 3926763.
- Nakamura M, Ueda N, Kishimoto K, Yoshimoto T, Yamamoto S, Ishimura K (březen 1995). „Imunocytochemická lokalizace arachidonát 12-lipoxygenázy destičkového typu v krevních buňkách myší“. The Journal of Histochemistry and Cytochemistry. 43 (3): 237–44. doi:10.1177/43.3.7868854. PMID 7868854.
- Hussain H, Shornick LP, Shannon VR, Wilson JD, Funk CD, Pentland AP, Holtzman MJ (leden 1994). „Epidermis obsahuje destičkovou 12-lipoxygenázu, která je nadměrně exprimována v zárodečných vrstvách keratinocytů u psoriázy“. Americký žurnál fyziologie. 266 (1 Pt 1): C243-53. doi:10.1152 / ajpcell.1994.266.1.C243. PMID 8304420.
- Arora JK, Lysz TW, Zelenka PS (červen 1996). „Role 12 (S) -HETE v reakci lidských epiteliálních buněk čočky na epidermální růstový faktor a inzulín“. Investigativní oftalmologie a vizuální věda. 37 (7): 1411–8. PMID 8641843.
- Hagmann W, Gao X, Timar J, Chen YQ, Strohmaier AR, Fahrenkopf C, Kagawa D, Lee M, Zacharek A, Honn KV (listopad 1996). „12-Lipoxygenáza v buňkách A431: genetická identita, modulace exprese a intracelulární lokalizace“. Experimentální výzkum buněk. 228 (2): 197–205. doi:10.1006 / excr.1996.0317. PMID 8912711.
- Nakamura M, Yamamoto S, Ishimura K (květen 1997). „Subcelulární lokalizace arachidonátu 12-lipoxygenázy a morfologický účinek jeho nadměrné exprese na myší keratinocyty“. Výzkum buněk a tkání. 288 (2): 327–34. doi:10,1007 / s004410050818. PMID 9082968. S2CID 23548308.
- Nigam S, Kumar GS, Sutherland M, Schewe T, Ikawa H, Yamasaki Y, Ueda N, Yamamoto S (září 1999). "Metabolická suprese destičkového typu 12-lipoxygenázy v děložním čípku člověka s invazivním karcinomem". International Journal of Cancer. 82 (6): 827–31. doi:10.1002 / (SICI) 1097-0215 (19990909) 82: 6 <827 :: AID-IJC10> 3.0.CO; 2-Q. PMID 10446449.
- Tornhamre S, Elmqvist A, Lindgren JA (duben 2000). „15-lipoxygenace leukotrienu A (4). Studie účinnosti 12- a 15-lipoxygenázy při katalyzování tvorby lipoxinu“. Biochimica et Biophysica Acta. 1484 (2–3): 298–306. doi:10.1016 / S1388-1981 (00) 00017-2. PMID 10760478.
- Chen BK, Tsai TY, Huang HS, Chen LC, Chang WC, Tsai SB, Chang WC (2003). „Funkční role aktivace kinázy regulované extracelulárním signálem a indukce c-Jun v aktivaci promotoru indukovaného forbolesterem lidského genu pro 12 (S) -lipoxygenázu“. Journal of Biomedical Science. 9 (2): 156–65. doi:10.1159/000048212. PMID 11914583. S2CID 46753449.
- Vítěz I, Normolle DP, Shureiqi I, Sondak VK, Johnson T, Su L, Brenner DE (říjen 2002). „Exprese 12-lipoxygenázy jako biomarkeru pro karcinogenezi melanomu“. Výzkum melanomu. 12 (5): 429–34. doi:10.1097/00008390-200209000-00003. PMID 12394183. S2CID 27336312.
- Gu J, Wen Y, Mison A, Nadler JL (únor 2003). „Cesta 12-lipoxygenázy zvyšuje produkci aldosteronu, 3 ', 5'-cyklickou fosforylaci proteinu vázajícího se na prvek reakce vázajícího se na adenosin monofosfát a aktivaci protein kinázy aktivovanou p38 mitogenem v lidských adrenokortikálních buňkách H295R“. Endokrinologie. 144 (2): 534–43. doi:10.1210 / cs.2002-220580. PMID 12538614.
- Fridman C, EP Ojopi, Gregório SP, Ikenaga EH, Moreno DH, Demetrio FN, Guimarães PE, Vallada HP, Gattaz WF, Dias Neto E (únor 2003). „Sdružení nového polymorfismu v genu ALOX12 s bipolární poruchou“. Evropský archiv psychiatrie a klinické neurovědy. 253 (1): 40–3. doi:10.1007 / s00406-003-0404-r. PMID 12664313. S2CID 21064663.