Jurský - Jurassic - Wikipedia
| jurský | |
|---|---|
| 201.3 ± 0.2 – ~145.0 Ma | |
| Chronologie | |
Klíčové události v Jurassic -205 — – -200 — – -195 — – -190 — – -185 — – -180 — – -175 — – -170 — – -165 — – -160 — – -155 — – -150 — – -145 — – jak aproximovat časový rámec klíčových jurských událostí. Svislá osa: před miliony let. | |
| Etymologie | |
| Formální jméno | Formální |
| Informace o použití | |
| Nebeské tělo | Země |
| Regionální využití | Globální (ICS ) |
| Použitá časová stupnice | Časová stupnice ICS |
| Definice | |
| Chronologická jednotka | Doba |
| Stratigrafická jednotka | Systém |
| Formalita časového rozsahu | Formální |
| Definice dolní hranice | První výskyt Ammonit Psiloceras spelae tirolicum. |
| Dolní hranice GSSP | Sekce Kuhjoch, Pohoří Karwendel, Severní vápenaté Alpy, Rakousko 47 ° 29'02 ″ severní šířky 11 ° 31'50 ″ V / 47,4839 ° N 11,5306 ° E |
| GSSP ratifikováno | 2010 |
| Definice horní hranice | Není formálně definováno |
| Kandidáti na definici horní hranice | Žádný |
| Horní hranice kandidátských sekcí GSSP |
|
| Atmosférická a klimatická data | |
| Střední atmosférický Ó 2 obsah | C. 26 obj.% (130% moderní) |
| Střední atmosférický CO 2 obsah | C. 1950 ppm (7krát předindustriální) |
| Střední povrchová teplota | C. 16,5 ° C (3 ° C nad moderní) |
The jurský (/dʒʊˈr…s.sɪk/ juu-RASS-ik;[1]) je geologické období a Systém která trvala 56 milionů let od konce Trias Období před 201,3 miliony let (Mya ) na začátek Křídový Období 145 Mya.[poznámka 1] Jurassic představuje střední období Druhohor. Jurassic je pojmenován po Pohoří Jura v Evropské Alpy, kde vápenec byly poprvé identifikovány vrstvy z období.
Začátek období byl ve znamení majora Událost zániku triasu – jury. Další dva zánik události, ke kterým došlo během období: Pliensbachian-toarciánský zánik v rané jure a Tithonian událost na konci;[4] ani jedna událost nepatří mezi Hromadné vyhynutí „velké pětky“, nicméně.
Jurské období je rozděleno na tři epochy: Brzy, Střední, a Pozdě. Podobně v stratigrafie, Jurassic je rozdělen na Lower Jurassic, Middle Jurassic a Upper Jurassic série skalních útvarů.
Na začátku Jurassic superkontinent Pangea začal rifting do dvou pozemských mas: Laurasia na sever a Gondwana na jih. Toto vytvořilo více pobřežních linií a posunulo kontinentální klima ze suchého na vlhké a mnoho z nich suchý pouště triasu byly nahrazeny bujnými deštnými pralesy.
Na souši se fauna přeměňovala z triasové fauny, v níž dominovali oba dinosauromorph a crocodylomorph archosaury, kterému dominuje dinosauři sama. První ptactvo také se objevil během jury, když se vyvinul z větve theropod dinosauři. Mezi další významné události patří výskyt prvních ještěrek a vývoj Therian savci. Krokodýli přešli z pozemského na vodní způsob života. Oceány byly obývány mořští plazi jako ichtyosaury a plesiosaurs, zatímco ptakoještěr byli dominantní létající obratlovců.
Etymologie
The chronostratigrafické výraz "Jurassic" je přímo spojen s Pohoří Jura, a pohoří hlavně po průběhu Hranice mezi Francií a Švýcarskem. Při prohlídce regionu v roce 1795[poznámka 2] Alexander von Humboldt poznal hlavně vápenec dominoval pohoří Jura jako samostatná formace, která nebyla zahrnuta do zavedeného stratigrafického systému definovaného Abraham Gottlob Werner a v roce 1799 jej pojmenoval „Jura-Kalkstein“ („jurský vápenec“).[Poznámka 3][7][8][9]
O třicet let později, v roce 1829, francouzský přírodovědec Alexandre Brongniart zveřejnil průzkum na různých terénech, které tvoří kůru Země. V této knize Brongniart označil terény pohoří Jura jako terény Jurassiques, čímž vytvořil a publikoval tento termín poprvé.[10]
Název „Jura“ je odvozen od keltského kořene * jor přes Gaulish * iuris „zalesněná hora“, z níž se jako místní název vypůjčil do latiny Juria a nakonec Jura.[7][8][11]
Geologie
Jurské období je rozděleno na tři epochy: Early, Middle a Late. Podobně v stratigrafie je Jurassic rozdělen na Nižší jura, Middle Jurassic, a Svrchní jura série skalních útvarů, také známý v Evropa tak jako Lias, Dogger a Malm.[12] Rozdělení Jurassic na tři části vzniklo Leopold von Buch.[9] Tyto tři epochy se dělí na kratší volaná časová období věky. Věk jury od nejmladšího po nejstarší je:
| Horní / pozdní jura | Tithonian | (152.1 ± 4 – 145 ± 4 Mya ) |
| Kimmeridgian | (157,3 ± 4 - 152,1 ± 4 Mya) | |
| Oxfordian | (163,5 ± 4 - 157,3 ± 4 Mya) | |
| Middle Jurassic | Callovian | (166,1 ± 4 - 163,5 ± 4 Mya) |
| Bathonian | (168,3 ± 3,5 - 166,1 ± 4 Mya) | |
| Bajocian | (170,3 ± 3 - 168,3 ± 3,5 Mya) | |
| Aalenian | (174,1 ± 2 - 170,3 ± 3 Mya) | |
| Nižší / Early Jurassic | Toarcian | (182,7 ± 1,5 - 174,1 ± 2 Mya) |
| Pliensbachian | (190,8 ± 1,5 - 182,7 ± 1,5 Mya) | |
| Sinemurian | (199,3 ± 1 - 190,8 ± 1,5 Mya) | |
| Hettangian | (201,3 ± 0,6 - 199,3 ± 1 Mya) |
Stratigrafie
Jurská stratigrafie je primárně založena na použití amonity tak jako fosílie indexu, s Datum prvního vzhledu konkrétních taxonů amonitů, které se používají k označení počátků stádií, a také menších časových rozpětí v rámci stadií, označovaných jako „amonitové zóny“, se také někdy dále dělí na subzóny. Globální stratigrafie je založena na standardních evropských amonitových zónách, přičemž ostatní regiony jsou kalibrovány podle evropských nástupnictví.[13]

Nejstarší část jurského období byla historicky označována jako Lias nebo Liassic, zhruba ekvivalentní co do rozsahu k Early Jurassic, ale také zahrnující část předcházejícího Rhaetian. Hettangianské jeviště pojmenoval švýcarský paleontolog Eugène Renevier v roce 1864 po Hettange-Grande v severovýchodní Francii. The Sekce a bod globálního hraničního stratotypu (GSSP) pro základnu Hettangian se nachází na průsmyku Kuhjoch, Pohoří Karwendel, Severní vápenaté Alpy, Rakousko, která byla ratifikována v roce 2010. Začátek Hettangian, a tedy Jurassic jako celek, je poznamenán první dojem z amonit Psiloceras spelae tirolicum.[14] Základna Jurassic byla dříve definována jako první výskyt Psiloceras planorbis Albert Oppel v letech 1856-58, ale toto se změnilo, protože vzhled byl považován za příliš lokalizovanou událost pro mezinárodní hranice.[13]
Sinemurian etapa byla definována a zavedena do vědecké literatury francouzským paleontologem Alcide d'Orbigny v roce 1842. Název si vzal podle francouzského města Semur-en-Auxois, blízko Dijon. Původní definice Sinemurian zahrnovala to, co je nyní Hettangian. GSSP Sinemurian se nachází na útesu severně od osady East Quantoxhead, 6 km východně od Watchet, Somerset, Anglie, v rámci Blue Lias. Začátek Sinemurian je definován prvním výskytem ammonite Vermiceras quantoxense.[13][15]
Pleinsbachian byl pojmenován německým paleontologem Albert Oppel v roce 1858 po vesničce Pliensbach v komunitě Zell unter Aichelberg v Swabian Alb, blízko Stuttgart, Německo. GSSP pro základnu Pleinsbachian se nachází v lokalitě Wine Haven v Robin Hood's Bay, Yorkshire, Anglie, v Redcar Mudstone Formation. Začátek Pleinsbachian je definován prvním výskytem amonitu Bifericeras donovani.[16]
Toarcian je pojmenován po vesnici Thouars (Latinský: Toarcium), jižně od Saumur v Údolí Loiry z Francie, bylo definováno Alcide d'Orbigny v roce 1842 původně z lomu Vrines asi 2 km severozápadně od vesnice. GSSP pro základnu Toarcian se nachází na Peniche, Portugalsko. Hranice je definována prvním výskytem amonitů patřících do podrodu Dactylioceras (Eodactylioceras).[17]
Aalenian je pojmenován po městě Aalen v Německu. Aalenian definoval švýcarský geolog Karl Mayer-Eymar v roce 1864. Dolní hranice byla původně mezi tmavými jíly pohoří Black Jurassic a nadložní jílovitý pískovec a železitý oolit z Brown Jurassic sekvence jihozápadního Německa.[13] GSSP pro základnu Aalenian se nachází na Fuentelsaz v Pyrenejský rozsah u Guadalajara, Španělsko. Základna Aalenian je definována prvním výskytem amonitu Leioceras opalinum.[18]
Bajocian je pojmenován podle města Bayeux (Latinský: Bajoce) v Normandii ve Francii a byl definován Alcide d'Orbigny v roce 1842. GSSP pro základnu Bajocian se nachází v Murtinheira v Portugalsku a byla definována v roce 1997. Základna Bajocian je definována prvním výskytem amonit Hyperlioceras mundum.[19]
Bathonian je pojmenován po městě Koupel, Anglie, představil belgický geolog d'Omalius d'Halloy v roce 1843, po neúplném úseku oolitických vápenců v několika lomech v regionu. GSSP pro základnu Bathonian je Ravin du Bès, oblast Bas-Auran, Alpes de Haute Provence, Francie, která byla definována v roce 2009. Základna Bathonian je definována prvním výskytem amonitu Gonolkites conversgens, na základně Zigzagiceras cikcak amonitová zóna.[20]
Callovian je odvozen z latinizovaného názvu vesnice Kellaways ve Wiltshire v Anglii a byla definována Alcide d'Orbigny v roce 1852, původně se základnou v kontaktu mezi Formace lesního mramoru a Formace kukuřice. Později však bylo zjištěno, že tato hranice leží v horní části Bathonian. Základna Callovian ještě nemá certifikovaný GSSP, od roku 2019.[13]
Oxfordian je pojmenován po městě Oxford v Anglii, a byl jmenován Alcide d'Orbigny v roce 1844 v odkazu na Oxford Clay. Na základně Oxfordian chybí definovaný GSSP. W. J. Arkell ve studiích v letech 1939 a 1946 umístil dolní hranici Oxfordian jako první výskyt amonitů Quenstedtoceras mariae (poté umístěna do rodu Vertumniceras ). Následující návrhy navrhly první výskyt Cardioceras redcliffense jako spodní hranice.[13]
Kimmeridgian je pojmenován po vesnici Kimmeridge na pobřeží Dorset, Anglie. To bylo jmenováno Alcide d'Orbigny v roce 1842, v odkazu na Kimmeridge Clay. I když to nebylo potvrzeno, sekce Flodigarry na adrese Staffin Bay na Ostrov Skye, Skotsko byl předložen jako GSSP pro základnu Kimmeridgian.[21]
Tithonian byl zaveden do vědecké literatury Albertem Oppelem v roce 1865. Název Tithonian je neobvyklý v geologických uměleckých jménech, protože je odvozen z řecká mytologie spíše než placename. Tithonus byl synem Laomedon z Troy a zamiloval se do Eos, řecká bohyně svítání. Jeho jméno si pro to vybral Albert Oppel stratigrafické fázi, protože Tithonian se ocitá ruku v ruce s úsvitem křídy. Na základně Tithonian v současné době chybí GSSP.[13] Horní hranice Jurassic je také aktuálně nedefinovaná. Calpionellidy, záhadná skupina pelagických protistů s kalcitovitým tvarem urny testy krátce hojné během poslední jury až po nejstarší křídu, bylo navrženo, aby představovaly nejslibnější kandidáty na stanovení hranice J / K. [22]
Nerostné a uhlovodíkové usazeniny
Kimmeridge Clay a ekvivalenty jsou hlavní zdrojová skála pro Ropa ze Severního moře.[23] Arabská mísa Intrashelf Basin, uložená od konce střední do horní jury, je prostředím největších světových zásob ropy, včetně Ghawar Field, největší ropné pole na světě.[24] Jurský věk Sargelu[25] a formace Naokelekan[26] jsou v Iráku hlavními zdrojovými kameny ropy. Více než 1 500 gigatonů zásob jurského uhlí se nachází v severozápadní Číně, zejména v Turpan-Hami Basin a Ordos Basin.[27]
Impaktní krátery
Mezi hlavní impaktní krátery patří Kráter Morokweng, kráter o průměru 70 km pohřben pod pouští Kalahari v severní Jihoafrické republice. Náraz je datován k hranici jury a křídy, kolem 145 Ma. Bylo navrženo, aby kráter Morokweng měl roli v obratu při přechodu jury a křídy.[28] Dalším významným impaktní kráterem je Kráter Puchezh-Katunki, 40-80 kilometrů v průměru, pohřben pod Nižnij Novgorod Oblast, Rusko. Dopad byl datován do Sinemurian, kolem 192-196 Mya.[29]
Paleogeografie a tektonika


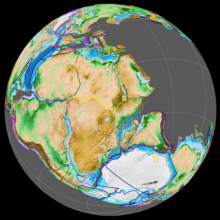
Během raného jury období superkontinent Pangea se rozešli na severní superkontinent Laurasia a jižní superkontinent Gondwana; the Mexický záliv otevřel se v nové trhlině mezi Severní Amerika a co je nyní mexické Poloostrov Yucatán. Jurský sever Atlantický oceán byl relativně úzký, zatímco jižní Atlantik se otevřel až v následujícím křídovém období, kdy se Afrika a Jižní Amerika rozpadly.[30] Kontinenty byly obklopeny Panthalassa, s Oceán Tethys mezi Gondwanou a Asií. Západní a střední Evropa tvořila souostroví ostrovů obklopených mělkými moři. Madagaskar a Antarktida se začaly vytrhávat z Afriky během rané jury a začaly fragmentaci Gondwany.[31][32] Podnebí bylo teplé, bez známek a ledovec když se objevil. Stejně jako v triasu nebyla nad jedním pólem zjevně žádná země a neexistovaly žádné rozsáhlé ledové čepice.
Na základě odhadovaných křivek hladiny moře byla hladina moře blízká současné úrovni během Hettangian a Sinemurian, během pozdního Sinemurian-Pleinsbachian stoupala o několik desítek metrů, než se pozdní Pleinsbachian snížila na téměř současnou úroveň. Zdá se, že během Toarcian došlo k postupnému vzestupu na vrchol ~ 75 m nad současnou hladinou moře. Během poslední části Toacianského moře hladina moře opět klesá o několik desítek metrů. Hladina moře postupně stoupala od Aalenian, kromě poklesů několika desítek metrů v Bajocian a kolem hranice Callovian-Oxfordian, které vyvrcholily v mořské hladině možná až 140 metrů nad současnou hladinou moře v Kimmeridgian-Tithonian hranice. Hladiny moře klesají v pozdním Tithonian, asi na 100 metrů, než se odskočí na zhruba 110 metrů na hranici Tithonian-Berriasian.[33]
Jurský geologický záznam je na západě dobrý Evropa, kde rozsáhlé námořní sekvence naznačují dobu, kdy byla velká část této budoucí pevniny ponořena pod mělkými tropickými moři; slavná národní prostředí zahrnují Jurské pobřeží Světové dědictví UNESCO v jižní Anglii a proslulý pozdní Jurassic lagerstätten z Holzmaden a Solnhofen v Německu.[34] Naproti tomu severoamerický jurský rekord je nejchudší z druhohor, s několika výchozy na povrchu.[35] Ačkoli epikontinentální Sundance moře během pozdní jury zanechaly mořská ložiska v částech severních plání Spojených států a Kanady, nejvíce exponované sedimenty z tohoto období jsou kontinentální, například naplavené vklady Morrison formace.
Jurassic byl čas kalcitové moře geochemie, ve které je nízký obsah hořčíku kalcit byla primární anorganická mořská sraženina uhličitanu vápenatého. Uhličitanová tvrdá zemina byly tedy velmi časté spolu s kalcitem ooids, kalcitové cementy a fauny bezobratlých s dominantní kalcitickou kostrou.[36]
První z několika masivních batoliti byli umístěni na severu Americká cordillera začínající v polovině Jurassic, značení Nevadská vrásnění.[37]
V Africe jsou raně jurské vrstvy distribuovány podobným způsobem jako postele z pozdního triasu, s častějšími výchozy na jihu a méně častými fosilními lůžky, kterým dominují cesty na sever.[38] Jak pokračovala jura, v Africe se rozmnožily větší a ikoničtější skupiny dinosaurů, jako jsou sauropody a ornitopody.[38] Střední jurské vrstvy nejsou v Africe ani dobře zastoupeny, ani dobře studovány.[38] Pozdně jurské vrstvy jsou také špatně zastoupeny, kromě velkolepých Tendaguru fauna v Tanzanii.[38] Pozdnějurský život Tendaguru je velmi podobný že nalezený v západní části Severní Ameriky Morrison formace.[38]

Jurské vápence a slíny ( Matmorská formace ) v jižní části Izrael

Horní jura Morrison formace v Colorado je jedním z nejúrodnějších zdrojů fosilií dinosaurů v Severní Americe

Gigandipus, a dinosaurus stopa ve spodní jure Moenave formace v objevovacím místě dinosaurů svatého Jiří na Johnson Farm, jihozápadní Utah

The Permu prostřednictvím Jurské stratigrafie Colorado Plateau oblast jihovýchodního Utahu




Flóra


Suché kontinentální podmínky charakteristické pro Trias během jury se ustavičně uvolňoval, zejména ve vyšších zeměpisných šířkách; teplé a vlhké podnebí umožňovalo svěží džungli pokrýt většinu krajiny.[39] Gymnospermy byly během Jurského období relativně různorodé.[12] Jehličnany zejména dominoval flóře, jako během triasu; byli nejrozmanitější skupinou a tvořili většinu velkých stromů.
Mezi existující rodiny jehličnanů, které vzkvétaly během jury, patřilo Araucariaceae, Cephalotaxaceae, Pinaceae, Podocarpaceae, Taxaceae a Taxodiaceae.[40] Zaniklá druhohorní rodina jehličnanů Cheirolepidiaceae dominovala vegetace s nízkou zeměpisnou šířkou, stejně jako křoví Bennettitales.[41] Cykasy, podobně jako palmy, byly také běžné ginkgo a stromové kapradiny v lese.[12] Menší kapradiny byly pravděpodobně dominantní podrost. Caytoniaceous semeno kapradiny byly během této doby další skupinou důležitých rostlin a předpokládá se, že byly keřů až malých stromů.[42] Rostliny ginka byly obzvláště běžné ve středních až vysokých severních zeměpisných šířkách.[12] Na jižní polokouli podokarpy byli obzvláště úspěšní Ginkgos a Czekanowskiales byly vzácné.[39][41]
V oceánech moderní korálové řasy se objevil poprvé.[12]
Fauna
Vodní a námořní
Během jury byly hlavními obratlovci žijícími v moři Ryba a námořní plazi. Mezi tyto patří ichtyosaury, které byly na vrcholu své rozmanitosti, plesiosaurs, počítaje v to pliosaury a námořní thalattosuchian crocodyliformes rodin Teleosauridae, Machimosauridae a Metriorhynchidae.[43] Během pozdní jury krmení filtru pachycormiform ryby se začnou diverzifikovat, včetně největších kostnatých ryb, o kterých je známo, že existovaly, Leedsichthys, s odhadovanou maximální délkou přes 15 metrů.[44] Během Middle Jurassic, nejstarší známý makrely žraloky objevil, představovaný rodem Palaeocarcharias.[45]
V bezobratlý světě se objevilo několik nových skupin, včetně rudisté (A útes -formující rozmanitost mlži ) a belemnites. Vápnitý sabellids (Glomerula ) se objevil v rané jure.[46][47] Jurassic měl také různou inkrustaci a nudný (sklerobiont) a došlo k výraznému nárůstu v bioeroze uhličitanových skořápek a tvrdých zemin. Obzvláště běžné je ichnogenus (stopové fosilie ) Gastrochaenolity.[48] Ammoniti byly hojné po celé jury, všechny jurské amonity pocházejí z Phylloceratina, jediná skupina amonitů, která přežila vyhynutí T-J.[49]
Během jury asi čtyři nebo pět z dvanácti clades planktonových organismů, které existují ve fosilních záznamech, buď zažilo masivní evoluční záření, nebo se objevilo poprvé.[12]

Vymřelý plaz z nižších (raných) jurských břidlic na jihu Německo představoval a delfín -jako tvar těla.

Plesiosaurs jako Muraenosaurus bloudili jurskými oceány.


Pozemní

Na souši, různé archosaurian plazi zůstali dominantní. Jurassic byl zlatým věkem pro velké býložravé dinosaury známé jako sauropody —Camarasaurus, Apatosaurus, Diplodocus, Brachiosaurus a mnoho dalších - kteří se toulali po zemi pozdě v období; jejich potravní pozemky byly buď prérie z kapradiny jako dlaň cykasy a bennettitales nebo vyšší růst jehličnanů podle jejich adaptací. Menší Ornithischian býložravé dinosaury, jako stegosaury a malé ornitopody byly méně převládající, ale hrály důležité role. Byli loveni velkými teropody, jako Ceratosaurus, Megalosaurus, Torvosaurus a Allosaurus, všechny tyto patří k ‚ještěrka hipped 'nebo saurischian pobočka dinosauři.[50]
Během pozdní jury první avialans, jako Archeopteryx, vyvinul od malého coelurosaurian dinosauři. Ve vzduchu, ptakoještěr byly běžné; ovládali oblohu a plnili mnoho ekologických rolí, které nyní zaujímají ptactvo,[51] a možná už vyprodukovali jedny z největších létajících zvířat všech dob.[52][53] V podrostu byly různé typy časných savci, stejně jako tritylodonty jako ještěrka sphenodonts a brzy lissamphibians. Zbytek Lissamphibia se v tomto období vyvinuly zavedením prvního mloci a caecilians.[54]

Diplodocus, dosahující délky přes 30 m, byl běžný sauropod během pozdní jury.

Allosaurus byl jedním z největších suchozemských predátorů během jury.

Stegosaurus je jedním z nejznámějších rodů dinosaurů a žil během střední až pozdní jury.

Aurornis xui, který žil v pozdní jurě, může být nejvíce primitivní avialan dinosaurus, který je dosud znám, a je jedním z prvních dosud nalezených avialanů.




Plazi
Želvy
Kmenová skupina želvy (Testudinata ) by se během jury diverzifikoval. Jurské kmenové želvy patří ke dvěma progresivně pokročilejším subtypům, k Mezochelydie a Perichelydie.[55] Předpokládá se, že předkové pro Mesochelydians jsou vodní, na rozdíl od pozemských pro Testudinata.[56] Dvě moderní skupiny želv (Testudiny ), Pleurodires a Kryptoměny, by se odchýlil od středního jury.[57][58] Rané linie kryptoměn jako Xinjiangchelyidae jsou známí ze střední jury,[59] zatímco raná kmenová pleurodire linie, Platychelyidae je známo z pozdní jury.[60]
Lepidosaury

Lepidosaury, který zahrnuje squamaty (ještěrky a hadi) a rhynchocephalians (který dnes zahrnuje pouze Tuatara ) by se během jury diverzifikoval. Rynchocephalians by během Jurassic obsadí širokou škálu morfologií a životních stylů, jako je specializovaná vodní Pleurosauridae a také jako býložravci Opisthodontia.[61] Squamates by také diverzifikoval během Jurassic, s odhadovaným původem živých ještěrek během Early Jurassic (~ 190 Mya) a divergence většiny hlavních squamatan skupin během Early-Middle Jurassic.[62] Mnoho jurských squamatů má nejasné vztahy k živým skupinám.[63] První členové hadí linie Ophidia se objeví během Middle Jurassic.[64]
Obojživelníci

Drtivá většina z temnospondyly vyhynuli na konci triasu, pouze s brachyopoidy přežívající do jury i mimo ni. Členové rodiny Brachyopidae jsou známy z jurských ložisek v Asii,[65] zatímco chigutisaurid Siderops je znám z rané jury Austrálie.[66] Moderní lissamphibians by se také začaly diverzifikovat během jury. Early Jurassic Prosalirus myslel, že představuje první žabího příbuzného s morfologií schopnou skákat jako živé žáby.[67] Morfologicky rozeznatelné stonkové žáby Notobatrachus jsou známy ze střední jury.[68] Zatímco první obojživelníci mločí linie jsou známí z triasu.[69] Crown skupina mloci se poprvé objevují během Middle-Late Jurassic v Eurasii, vedle příbuzných kmenových skupin. Mnoho salamandrů ze skupiny jurských kmenů Marmorerpeton a Kokartus jsou považovány za byly neotenický.[70] Mezi rané zástupce mloků skupiny korun patří Chunerpeton, Pangerpeton a Linglongtriton od Middle-Late Jurassic Yanliao Biota z Číny, které patří k Kryptobranchoidea, který obsahuje bydlení asijský a obří mloci.[71] Zatímco Beiyanerpeton, také ze stejné bioty je považován za časného člena Salamandroidea, skupina, která obsahuje všechny ostatní živé mloky.[72] Salamandři by se do Severní Ameriky rozptýlili do konce Jurassic, o čemž svědčí Iridotriton nalezený v pozdně jurském morrisonském souvrství.[73] Nejstarší nesporný kmencaecilian je Early Jurassic Eocaecilia z Arizony.[74] Čtvrtá skupina lissamphibians, zaniklý albanerpetontids, se nejprve objeví ve střední jure, zastoupené Anoualerpeton priscus z Bathonian v Británii, stejně jako neurčité pozůstatky od ekvivalentně věku Anoual formace Maroka.[75]
Savci
Savci se během jury značně diverzifikovali, patří k významným skupinám jurských savců Morganucodonta, Docodonta, Eutriconodonta, Dryolestida, Haramiyida a Multituberculata. Zatímco většina jurských savců je známa pouze z izolovaných zubů a úlomků čelistí, výjimečně zachované pozůstatky odhalily různé životní styly. Docodontan Castorocauda byl upraven pro vodní život, podobně jako ptakopysk a vydry.[76] Někteří členové Haramiyidy[77] a kmen eutriconodontan Volaticotherini[78] vlastnil a patagium podobný těm z létající veverky, což jim umožňuje klouzat vzduchem. Fruitafossor byl pravděpodobně specialistou na koloniální hmyz, podobně jako živý mravenečníci.[79] Non-savec cynodonty z rodiny Tritylodontidae přežil vyhynutí T-J a nadále existoval prostřednictvím rané křídy. Theriiform savci, dnes zastoupeni živými placenty a vačnatci, se také objeví během rané pozdní jury, zastoupené Juramaia, A eutherian savec blíže k původu placentálů než vačnatci.[80] Juramaia je mnohem pokročilejší, než se na svůj věk očekávalo, protože ostatní teriiformní savci se objevují až ve starší křídě.[81]
Hmyz a pavoukovci
Mnoho významných fosilních lokalit hmyzu je známo z jury Eurasie, z nichž nejdůležitější je Karabastau formace Kazachstánu a různá ložiska Yanliao Biota ve Vnitřním Mongolsku v Číně, například Daohugou postel, datování do Callovian -Oxfordian. Rozmanitost hmyzu stagnovala v celé rané a střední jurě, ale během druhé třetiny jurského původu se podstatně zvýšila, zatímco míra vymírání zůstala stejná.[82] Middle-Late Jurassic was a time of major diverisification for brouci.[83] Weevils se poprvé objevují ve fosilních záznamech během střední a vrchní jury, ale existuje podezření, že vznikly během pozdního triasu a rané jury.[84] Nejstarší známý lepidopterani (skupina obsahující motýly a můry) jsou známy z trias-jury hranice, s křídlovými šupinami patřícími do podřádu Glossata a Micropterigidae -gradovat můry z vkladů tohoto věku v Německu.[85] Ačkoli moderní zástupci nejsou známi až do kenozoika, ektoparazitický hmyz, o kterém se předpokládá, že představuje a kmenová skupina na blechy se poprvé objeví během jury, jako např Pseudopulex jurassicus. Tento hmyz se podstatně liší od moderních blech, postrádá specializovanou morfologii moderních blech a je větší.[86][87] Nejstarší skupina Phasmatodea (lepkavý hmyz), okřídlený Susumanioidea, outgroup to living Phasmatodeans, first appear during the Middle Jurassic.[88] Nejstarší člen Mantophasmatidae (gladiátoři) se během této doby také objevili.[89]
Z jury je známa jen hrstka záznamů o roztočích Jureremus, an Oribatid roztoč patřící do rodiny Cymbaeremaeidae známý z horní jury Británie a Ruska.[90] Pavouci by se diverzifikovali prostřednictvím jury.[91] Early Jurassic Seppo koponeni pravděpodobně představuje kmenovou skupinu Palpimanoidea.[92] Eoplectreurys ze střední jury Číny se považuje za kmenovou linii Synspermiata. Nejstarší člen rodiny Archaeidae, Patarchaea, je znám ze střední jury.[91] Mongolarachne ze střední jury v Číně patří mezi největší známé fosilní pavouky s nohama o délce přes 5 centimetrů.[93] Jediným štírem známým z jury je Liassoscorpionides z německé dolní jury, nejistého umístění.[94] Eupnoi Opiliones jsou známí ze střední jury, včetně členů rodiny Sclerosomatidae.[95][96]
V populární kultuře
Od počátku 90. let, termín jurský byl popularizován Jurský park franšíza, která začala v roce 1990 s Michael Crichton je román stejného titulu a jeho filmová adaptace, poprvé vydáno v roce 1993.
Viz také
Reference
Poznámky
- ^ V roce 2014 byl na základě stratigrafické studie navržen věk 140 Ma pro jurskou křídu namísto obvykle přijímaných 145 Ma. Formace Vaca Muerta v Neuquénská pánev, Argentina.[2] Víctor Ramos, jeden z autorů studie, která navrhuje hraniční věk 140 Ma, považuje studii za „první krok“ k formální změně věku v Mezinárodní unie geologických věd.[3]
- ^ "Ich hatte mich auf einer geognostischen Reise, die ich 1795 durch das südliche Franken, die westliche Schweiz und Ober-Italien machte, davon überzeugt, daß der Jura-Kalkstein, welchen Werner zu seinem Muschelkalk rechnete, eine eigne formation bildete. Ve hře Schrift über die unterirdischen Gasarten, welche mein Bruder Wilhelm von Humboldt 1799 während meines Aufenthalts in Südamerika herausgab, wird der Formation, die ich vorläufig mit dem Namen Jura-Kalkstein bezeichnete, zuerst gedacht.„(, Na geologické prohlídce, kterou jsem podnikl v roce 1795 jižní Francií, západním Švýcarskem a horní Itálií, jsem se přesvědčil, že jurský vápenec, který Werner zahrnul do svého lasturového vápence, představuje samostatný útvar. plyny, které můj bratr Wilhelm von Humboldt zveřejnil v roce 1799 během mého pobytu v Jižní Americe, je nejprve koncipována formace, kterou jsem prozatímně označil jménem „Jura vápenec“.)[5]
- ^ "[…] Die ausgebreitete Formation, welche zwischen dem alten Gips und neueren Sandstein liegt, und welchen ich vorläufig mit dem Nahmen Jura-Kalkstein bezeichne.„… Rozšířená formace, která leží mezi starým sádrovcem a novějším pískovcem a kterou prozatímně označuji jménem„ vápenec Jura “.“[6]
Citace
- ^ "Jurský". Dictionary.com Nezkrácené. Random House.
- ^ Vennari a kol. 2014, str. 374-385.
- ^ Jaramillo 2014.
- ^ Hallam 1986, str. 765-768.
- ^ von Humboldt 1858, str. 632.
- ^ von Humboldt 1799, str. 39.
- ^ A b Hölder 1964.
- ^ A b Arkell 1956.
- ^ A b Pieńkowski a kol. 2008, str. 823–922.
- ^ Brongniart 1829.
- ^ Válec 1903.
- ^ A b C d E F Kazlev 2002.
- ^ A b C d E F G Ogg, J.G .; Hinnov, L. A.; Huang, C. (2012), "Jurský", Geologická časová stupnice, Elsevier, str. 731–791, doi:10.1016 / b978-0-444-59425-9.00026-3, ISBN 978-0-444-59425-9, vyvoláno 2020-12-05
- ^ Hillebrandt, A.v .; Krystyn, L .; Kürschner, W.M .; Bonis, N.R .; Ruhl, M .; Richoz, S .; Schobben, M. A. N .; Urlichs, M .; Bown, P.R .; Kment, K .; McRoberts, C.A. (2013-09-01). „Global Stratotype Sections and Point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains, Northern Calcareous Alps, Tyrol, Austria)“. Epizody. 36 (3): 162–198. doi:10.18814 / epiiugs / 2013 / v36i3 / 001. ISSN 0705-3797.
- ^ Bloos, Gert; Page, Kevin N. (2002-03-01). „Global Stratotype Section and Point for base of the Sinemurian Stage (Lower Jurassic)“. Epizody. 25 (1): 22–28. doi:10.18814 / epiiugs / 2002 / v25i1 / 003. ISSN 0705-3797.
- ^ Meister, Christian; Aberhan, Martin; Blau, Joachim; Dommergues, Jean-Louis; Feist-Burkhardt, Susanne; Hailwood, Ernie A .; Hart, Malcom; Hesselbo, Stephen P .; Hounslow, Mark W .; Hylton, Mark; Morton, Nicol (2006-06-01). „The Global Boundary Stratotype Section and Point (GSSP) for the base of Pliensbachian Stage (Lower Jurassic), Wine Haven, Yorkshire, UK“. Epizody. 29 (2): 93–106. doi:10.18814 / epiiugs / 2006 / v29i2 / 003. ISSN 0705-3797.
- ^ Fantasia, Alicia; Adatte, Thierry; Spangenberg, Jorge E .; Písmo, Eric; Duarte, Luís V .; Föllmi, Karl B. (listopad 2019). „Globální versus lokální procesy během přechodu Pliensbachian – Toarcian na Peniche GSSP, Portugalsko: záznam s více proxy servery“. Recenze vědy o Zemi. 198: 102932. Bibcode:2019ESRv..19802932F. doi:10.1016 / j.earscirev.2019.102932.
- ^ Barrón, Eduardo; Ureta, Soledad; Goy, Antonio; Lassaletta, Luis (srpen 2010). „Palynology of Toarcian – Aalenian Global Boundary Stratotype Section and Point (GSSP) at Fuentelsaz (Lower-Middle Jurassic, Iberian Range, Spain)“. Recenze paleobotaniky a palynologie. 162 (1): 11–28. doi:10.1016 / j.revpalbo.2010.04.003.
- ^ Pavia, G .; Enay, R. (01.03.1997). „Definice hranice aalénsko-bajociánské etapy“. Epizody. 20 (1): 16–22. doi:10.18814 / epiiugs / 1997 / v20i1 / 004. ISSN 0705-3797.
- ^ López, Fernández; Rafael, Sixto; Pavia, Giulio; Erba, Elisabetta; Guiomar, Myette; Paiva Henriques, María Helena; Lanza, Roberto; Mangold, Charles; Morton, Nicol; Olivero, Davide; Tiraboschi, Daniele (2009). „The Global Boundary Stratotype Section and Point (GSSP) for base of the Bathonian Stage (Middle Jurassic), Ravin du Bès Section, SE France“ (PDF). Epizody. 32 (4): 222–248. doi:10.18814 / epiiugs / 2009 / v32i4 / 001. Archivovány od originál (PDF) dne 4. března 2016. Citováno 5. června 2015.
- ^ BARSKI, Marcin (06.09.2018). „Dinoflagelátová cysta se shromažďuje přes hranici Oxfordian / Kimmeridgian (horní jura) v Flodigarry, Staffin Bay, ostrov Skye, Skotsko - navrhovaný GSSP pro základnu Kimmeridgian“. Volumina Jurassica. XV (1): 51–62. doi:10.5604/01.3001.0012.4594. ISSN 1731-3708.
- ^ WIMBLEDON, William A.P. (2017-12-27). „Vývoj se stanovením hranice Tithonian / Berriasian (J / K)“. Volumina Jurassica (1): 0. doi:10.5604/01.3001.0010.7467. ISSN 1731-3708.
- ^ Gautier D.L. (2005). „Celkový ropný systém kimmeridiánských břidlic provincie Graben v Severním moři“ (PDF). Geologický průzkum Spojených států. Citováno 2. listopadu 2018.
- ^ Wilson, A. O. (2020). „Kapitola 1 Úvod do jurské arabské vnitrozemské pánve“. Geologická společnost, Londýn, Monografie. 53 (1): 1–19. doi:10.1144 / M53.1. ISSN 0435-4052. S2CID 226967035.
- ^ Abdula, Rzger A. (srpen 2015). „Uhlovodíkový potenciál formace Sargelu a korelace zdrojů ropy, irácký Kurdistán“. Arabian Journal of Geosciences. 8 (8): 5845–5868. doi:10.1007 / s12517-014-1651-0. ISSN 1866-7511. S2CID 129120960.
- ^ Soranská univerzita; Abdula, Rzger A. (2016-10-16). „Zdrojové hodnocení formace naokelekanu v iráckém Kurdistánu“. Journal of Zankoy Sulaimani - část A. 19 (1): 103–124. doi:10.17656 / jzs.10589.
- ^ Ao, Weihua; Huang, Wenhui; Weng, Chengmin; Xiao, Xiuling; Liu, Dameng; Tang, Xiuyi; Chen, Ping; Zhao, Zhigen; Wan, Huan; Finkelman, Robert B. (leden 2012). „Petrologie uhlí a vznik jurského uhlí v pánvi Ordos v Číně“. Geoscience Frontiers. 3 (1): 85–95. doi:10.1016 / j.gsf.2011.09.004.
- ^ Tennant, Jonathan P .; Mannion, Philip D .; Upchurch, Paul (02.09.2016). „Dynamika tetrapodské diverzity regulovaná hladinou moře prostřednictvím jury / křídy“. Příroda komunikace. 7 (1): 12737. Bibcode:2016NatCo ... 712737T. doi:10.1038 / ncomms12737. ISSN 2041-1723. PMC 5025807. PMID 27587285.
- ^ Holm-Alwmark, S .; Alwmark, C .; Ferrière, L .; Lindström, S .; Meier, M. M. M .; Scherstén, A .; Herrmann, M .; Masaitis, V. L .; Mashchak, M. S .; Naumov, M. V .; Jourdan, F. (srpen 2019). „Časný Jurský věk pro nárazovou strukturu Puchezh-Katunki (Rusko) na základě dat a palynologie 40 Ar / 39 Ar“. Meteoritika a planetární věda. 54 (8): 1764–1780. Bibcode:2019M & PS ... 54.1764H. doi:10.1111 / maps.13309. ISSN 1086-9379.
- ^ Scotese 2003.
- ^ Geiger, Markus; Clark, David Norman; Mette, Wolfgang (březen 2004). „Přehodnocení načasování rozpadu Gondwany na základě sedimentologických a seismických důkazů z povodí Morondavy na Madagaskaru“. Journal of African Earth Sciences. 38 (4): 363–381. Bibcode:2004JAfES..38..363G. doi:10.1016 / j.jafrearsci.2004.02.003.
- ^ Nguyen, Luan C .; Hall, Stuart A .; Bird, Dale E .; Ball, Philip J. (červen 2016). „Rekonstrukce kontinentálních okrajů východní Afriky a Antarktidy: REKONSTRUKCE AFRIKA-ANTARCTIKA“. Journal of Geophysical Research: Solid Earth. 121 (6): 4156–4179. doi:10.1002 / 2015JB012776.
- ^ Haq, Bilal U. (01.01.2018). „Varianty na úrovni jurského moře: přehodnocení“. GSA dnes: 4–10. doi:10.1130 / GSATG359A.1.
- ^ "Země a moře během Jurassic". urweltmuseum.de. Archivovány od originál dne 2007-07-14.
- ^ „Severoamerická tapiserie času a terénu: Jurské skály - před 208 až 146 miliony let“. nationalatlas.gov. Archivovány od originál dne 2007-07-15.
- ^ Stanley & Hardie 1998.
- ^ Monroe & Wicander 1997, str. 607.
- ^ A b C d E Jacobs 1997, s. 2-4.
- ^ A b Haines 2000, str. 65.
- ^ Behrensmeyer et al. 1992, str. 349.
- ^ A b Behrensmeyer et al. 1992, str. 352.
- ^ Behrensmeyer et al. 1992, str. 353.
- ^ Motani 2000.
- ^ Liston, J., Newbrey, M., Challands, T. a Adams, C., 2013, „Growth, age and size of the Jurassic pachycormid Leedsichthys problematicus (Osteichthyes: Actinopterygii) in: Arratia, G., Schultze, H. and Wilson, M. (eds.) Mesozoic Fishes 5 - Global Diversity and Evolution. Verlag Dr. Friedrich Pfeil, Mnichov, Německo, str. 145–175
- ^ Jambura, Patrick L .; Kindlimann, René; López-Romero, Faviel; Marramà, Giuseppe; Pfaff, Cathrin; Stumpf, Sebastian; Türtscher, Julia; Underwood, Charlie J .; Ward, David J .; Kriwet, Jürgen (prosinec 2019). „Mikropočítačové tomografie zobrazují vývoj jedinečného vzoru mineralizace zubů u žraloků makrely (Chondrichthyes; Lamniformes) v hlubokém čase“. Vědecké zprávy. 9 (1): 9652. Bibcode:2019NatSR ... 9.9652J. doi:10.1038 / s41598-019-46081-3. ISSN 2045-2322. PMC 6609643. PMID 31273249.
- ^ Vinn & Mutvei 2009, str. 286-296.
- ^ Vinn, ten Hove & Mutvei 2008, str. 295–301.
- ^ Taylor & Wilson 2003, str. 1-103.
- ^ Page, Kevin N. (leden 2008). „Vývoj a geografie jurských amonoidů“. Sborník sdružení geologů. 119 (1): 35–57. doi:10.1016 / S0016-7878 (08) 80257-X.
- ^ Haines 2000.
- ^ Feduccia 1996.
- ^ Witton, Martill & Loveridge 2010, str. 79-81.
- ^ Witton 2016.
- ^ Carroll 1988.
- ^ Joyce, Walter G. (duben 2017). „Přehled fosilních záznamů bazálních mezozoických želv“. Bulletin of Peabody Museum of Natural History. 58 (1): 65–113. doi:10.3374/014.058.0105. ISSN 0079-032X. S2CID 54982901.
- ^ Sterli, Juliana; de la Fuente, Marcelo S .; Rougier, Guillermo W. (07.07.2018). „Nové pozůstatky Condorchelys antiqua (Testudinata) z raně střední jury Patagonie: anatomie, fylogeneze a paedomorphosis v raném vývoji želv“. Časopis paleontologie obratlovců. 38 (4): (1)–(17). doi:10.1080/02724634.2018.1480112. ISSN 0272-4634. S2CID 109556104.
- ^ Wings et al. 2012, str. 925-935.
- ^ Gannon 2012.
- ^ Rabi, Márton; Joyce, Walter G .; Wings, Oliver (září 2010). „Přehled mezozoických želv v Junggarské pánvi (Sin-ťiang, severozápadní Čína) a paleobiogeografie jurských až raněkřídových asijských testudinátů“. Palaeobiodiversity a Palaeoenvironmentments. 90 (3): 259–273. doi:10.1007 / s12549-010-0031-3. ISSN 1867-1594. S2CID 128805609.
- ^ Sullivan, Patrick M .; Joyce, Walter G. (srpen 2017). "Anatomie skořápky a pánve pozdně jurské želvy Platychelys oberndorferi na základě materiálu ze Solothurnu ve Švýcarsku". Švýcarský věstník paleontologie. 136 (2): 323–343. doi:10.1007 / s13358-017-0136-7. ISSN 1664-2376. S2CID 90587841.
- ^ Herrera-Flores, Jorge A .; Stubbs, Thomas L .; Benton, Michael J. (2017). „Makroevoluční vzorce v Rhynchocephalia: je tuatara (Sphenodon punctatus) živou fosilií?“. Paleontologie. 60 (3): 319–328. doi:10.1111 / pala.12284. ISSN 1475-4983.
- ^ Burbrink, Frank T; Grazziotin, Felipe G; Pyron, R Alexander; Cundall, David; Donnellan, Steve; Ir, Frances; Keogh, J. Scott; Kraus, Fred; Murphy, Robert W; Noonan, Brice; Raxworthy, Christopher J (2020-05-01). Thomson, Robert (ed.). „Dotaz na data v měřítku genomu pro squamata (ještěrky, hady a amfisbaeny) neukazuje žádnou podporu pro klíčové tradiční morfologické vztahy“. Systematická biologie. 69 (3): 502–520. doi:10.1093 / sysbio / syz062. ISSN 1063-5157. PMID 31550008.
- ^ Dong, Liping; Wang, Yuan; Mou, Lijie; Zhang, Guoze; Evans, Susan E. (2019-09-13). „Nový Jurský ještěr z Číny“. Geodiversitas. 41 (16): 623. doi:10,5252 / geodiversitas2019v41a16. ISSN 1280-9659. S2CID 204256127.
- ^ Caldwell, M. W .; Nydam, R.L .; Palci, A .; Apesteguía, S. N. (2015). „Nejstarší známí hadi ze střední jury a spodní křídy poskytují poznatky o vývoji hadů“. Příroda komunikace. 6: 5996. Bibcode:2015NatCo ... 6,5996C. doi:10.1038 / ncomms6996. PMID 25625704.
- ^ Averianov, Alexander O .; Martin, Thomas; Skutschas, Pavel P .; Rezvyi, Anton S .; Bakirov, Aizek A. (březen 2008). "Obojživelníci ze středního jura Balabansai Svita v období deprese Fergana v Kyrgyzstánu (Střední Asie)". Paleontologie. 51 (2): 471–485. doi:10.1111 / j.1475-4983.2007.00748.x.
- ^ Warren, A. A .; Hutchinson, M. N .; Hill, Dorothy (1983-09-13). „Poslední Labyrinthodont? Nový brachyopoid (Amphibia, Temnospondyli) z rané formace Jurassic Evergreen v australském Queenslandu“. Filozofické transakce Královské společnosti v Londýně. B, Biologické vědy. 303 (1113): 1–62. Bibcode:1983RSPTB.303 ... 1W. doi:10.1098 / rstb.1983.0080.
- ^ Reilly, Stephen M .; Jorgensen, Michael E. (únor 2011). „Vývoj skákání u žab: morfologické důkazy o pohybovém stavu bazálního anuranu a radiaci pohybového systému u anuranů korunní skupiny“. Journal of Morphology. 272 (2): 149–168. doi:10.1002 / jmor.10902. PMID 21210487. S2CID 14217777.
- ^ Báez, Ana Maria; Nicoli, Laura (březen 2008). „Nový druh Notobatrachus (Amphibia, Salientia) ze střední jury severozápadní Patagonie“. Journal of Paleontology. 82 (2): 372–376. doi:10.1666/06-117.1. ISSN 0022-3360. S2CID 130032431.
- ^ Schoch, Rainer R .; Werneburg, Ralf; Voigt, Sebastian (2020-05-26). „Triasový mlok skvrnitý z Kyrgyzstánu a původ mloků“. Sborník Národní akademie věd. 117 (21): 11584–11588. doi:10.1073 / pnas.2001424117. ISSN 0027-8424. PMC 7261083. PMID 32393623.
- ^ Skutschas, Pavel; Stein, Koen (duben 2015). „Histologie dlouhých kostí kmene salamandra Kokartus honorarius (Amphibia: Caudata) ze střední jury v Kyrgyzstánu“. Anatomy Journal. 226 (4): 334–347. doi:10.1111 / joa.12281. PMC 4386933. PMID 25682890.
- ^ Jia, Jia; Gao, Ke-Qin (04.03.2019). "A new stem hynobiid salamander (Urodela, Cryptobranchoidea) from the Upper Jurassic (Oxfordian) of Liaoning Province, China". Časopis paleontologie obratlovců. 39 (2): e1588285. doi:10.1080/02724634.2019.1588285. ISSN 0272-4634. S2CID 164310171.
- ^ Gao, K.-Q .; Shubin, N. H. (2012-04-10). „Late Jurassic salamandroid from western Liaoning, China“. Sborník Národní akademie věd. 109 (15): 5767–5772. doi:10.1073 / pnas.1009828109. ISSN 0027-8424. PMC 3326464. PMID 22411790.
- ^ Evans, S. E.; Lally, C .; Chure, D. C .; Elder, A .; Maisano, J. A. (2005). „Pozdní jurský mlok (Amphibia: Caudata) z morrisonského souvrství Severní Ameriky“. Zoologický žurnál Linneanské společnosti. 143 (4): 599–616. doi:10.1111 / j.1096-3642.2005.00159.x.
- ^ Santos, Rodolfo Otávio; Laurin, Michel; Zaher, Hussam (2020-11-03). "A review of the fossil record of caecilians (Lissamphibia: Gymnophionomorpha) with comments on its use to calibrate molecular timetrees". Biologický žurnál společnosti Linnean. 131 (4): 737–755. doi:10.1093/biolinnean/blaa148. ISSN 0024-4066.
- ^ Haddoumi, Hamid; Allain, Ronan; Meslouh, Said; Metais, Grégoire; Monbaron, Michel; Pons, Denise; Rage, Jean-Claude; Vullo, Romain; Zouhri, Samir (leden 2016). „Guelb el Ahmar (Bathonian, Anoual Syncline, východní Maroko): první kontinentální flóra a fauna včetně savců ze střední jury Afriky“ (PDF). Výzkum v Gondwaně. 29 (1): 290–319. Bibcode:2016GondR..29..290H. doi:10.1016 / j.gr.2014.12.004. ISSN 1342-937X.
- ^ Ji, Q .; Luo, Z.-X .; Yuan, C.-X .; Tabrum, A. R. (2006). „Plavecký savec ze střední jury a ekomorfologická diverzifikace raných savců“. Věda. 311 (5, 764): 1, 123–1, 127. Bibcode:2006Sci ... 311.1123J. doi:10.1126 / science.1123026. PMID 16497926. S2CID 46067702.
- ^ Meng, Qing-Jin; Grossnickle, David M.; Liu, Di; Zhang, Yu-Guang; Neander, April I.; Ji, Qiang; Luo, Zhe-Xi (August 2017). "New gliding mammaliaforms from the Jurassic". Příroda. 548 (7667): 291–296. Bibcode:2017Natur.548..291M. doi:10.1038/nature23476. ISSN 1476-4687. PMID 28792929. S2CID 205259206.
- ^ Meng, J .; Hu, Y.-M.; Wang, Y.-Q.; Wang, X.-L .; Li, C.-K. (2007). "Corrigendum: A Mesozoic gliding mammal from northeastern China". Nature 446 (7131): 102. Bibcode:2007Natur.446Q.102M. doi:10.1038 / nature05639.
- ^ Luo, Z.-X .; Wible, J.R. (2005). "A Late Jurassic Digging Mammal and Early Mammalian Diversification". Věda. 308 (5718): 103–107. Bibcode:2005Sci...308..103L. doi:10.1126/science.1108875. ISSN 0036-8075. PMID 15802602. S2CID 7031381.
- ^ Zhe-Xi Luo; Chong-Xi Yuan; Qing-Jin Meng; Qiang Ji (25 August 2011). "A Jurassic eutherian mammal and divergence of marsupials and placentals" (PDF). Příroda. 476 (7361): 442–445. Bibcode:2011Natur.476..442L. doi:10.1038 / příroda10291. PMID 21866158. S2CID 205225806. Archivovány od originál (PDF) dne 10. listopadu 2013. Electronic supplementary material
- ^ King, Benedict; Beck, Robin M. D. (2020-06-10). "Tip dating supports novel resolutions of controversial relationships among early mammals". Sborník Královské společnosti B: Biologické vědy. 287 (1928): 20200943. doi:10.1098/rspb.2020.0943. PMC 7341916. PMID 32517606.
- ^ Labandeira, Conrad C. (2018-05-23), "The Fossil History of Insect Diversity", Biodiverzita hmyzu, Chichester, UK: John Wiley & Sons, Ltd, pp. 723–788, doi:10.1002/9781118945582.ch24, ISBN 978-1-118-94558-2, vyvoláno 2020-12-06
- ^ McKenna, Duane D .; Shin, Seunggwan; Ahrens, Dirk; Balke, Michael; Beza-Beza, Cristian; Clarke, Dave J.; Donath, Alexander; Escalona, Hermes E.; Friedrich, Frank; Letsch, Harald; Liu, Shanlin (2019-12-03). "The evolution and genomic basis of beetle diversity". Sborník Národní akademie věd. 116 (49): 24729–24737. doi:10.1073/pnas.1909655116. ISSN 0027-8424. PMC 6900523. PMID 31740605.
- ^ Shin, Seunggwan; Clarke, Dave J; Lemmon, Alan R; Moriarty Lemmon, Emily; Aitken, Alexander L; Haddad, Stephanie; Farrell, Brian D; Marvaldi, Adriana E; Oberprieler, Rolf G; McKenna, Duane D (2018-04-01). "Phylogenomic Data Yield New and Robust Insights into the Phylogeny and Evolution of Weevils". Molekulární biologie a evoluce. 35 (4): 823–836. doi:10.1093/molbev/msx324. ISSN 0737-4038. PMID 29294021. S2CID 4366092.
- ^ van Eldijk, Timo J. B.; Wappler, Torsten; Strother, Paul K .; van der Weijst, Carolien M. H.; Rajaei, Hossein; Visscher, Henk; van de Schootbrugge, Bas (January 2018). "A Triassic-Jurassic window into the evolution of Lepidoptera". Vědecké zálohy. 4 (1): e1701568. Bibcode:2018SciA....4.1568V. doi:10.1126/sciadv.1701568. ISSN 2375-2548. PMC 5770165. PMID 29349295.
- ^ Huang, DiYing; Engel, Michael S .; Cai, ChenYang; Nel, André (May 2013). „Mezozoické obří blechy ze severovýchodní Číny (Siphonaptera): taxonomie a důsledky pro paleodiverzitu“. Bulletin čínské vědy. 58 (14): 1682–1690. Bibcode:2013ChSBu..58.1682H. doi:10.1007 / s11434-013-5769-3. ISSN 1001-6538. S2CID 53578959.
- ^ Gao, Taiping; Shih, Chungkun; Rasnitsyn, Alexandr P .; Xu, Xing; Wang, Shuo; Ren, Dong (July 2013). „Nové přechodné blechy z Číny, které zdůrazňují rozmanitost ektoparazitického hmyzu z období křídy“. Aktuální biologie. 23 (13): 1261–1266. doi:10.1016 / j.cub.2013.05.040. PMID 23810530. S2CID 9646168.
- ^ Yang, Hongru; Shi, Chaofan; Engel, Michael S; Zhao, Zhipeng; Ren, Dong; Gao, Taiping (2020-04-02). "Early specializations for mimicry and defense in a Jurassic stick insect". National Science Review: nwaa056. doi:10.1093 / nsr / nwaa056. ISSN 2095-5138.
- ^ Huang, Di-ying; Nel, André; Zompro, Oliver; Waller, Alain (2008-06-11). "Mantophasmatodea now in the Jurassic". Naturwissenschaften. 95 (10): 947–952. Bibcode:2008NW.....95..947H. doi:10.1007/s00114-008-0412-x. ISSN 0028-1042. PMID 18545982. S2CID 35408984.
- ^ Selden, Paul A .; Baker, Anne S .; Phipps, Kenneth J. (2008). "An Oribatid Mite (arachnida: Acari) from the Oxford Clay (jurassic: Upper Callovian) of South Cave Station Quarry, Yorkshire, Uk". Paleontologie. 51 (3): 623–633. doi:10.1111/j.1475-4983.2008.00769.x. hdl:1808/8353. ISSN 1475-4983.
- ^ A b Magalhaes, Ivan L. F .; Azevedo, Guilherme H. F .; Michalik, Peter; Ramírez, Martín J. (February 2020). „Znovu se objevují fosilní záznamy pavouků: důsledky pro kalibraci stromů a důkazy o velkém faunálním obratu od druhohor“. Biologické recenze. 95 (1): 184–217. doi:10.1111 / brv.12559. ISSN 1464-7931. PMID 31713947. S2CID 207937170.
- ^ Selden, Paul A .; Dunlop, Jason A. (2014). "První fosilní pavouk (Araneae: Palpimanoidea) ze spodní jury (Grimmen, Německo)". Zootaxa. 3894 (1): 161–168. doi:10.11646 / zootaxa.3894.1.13. PMID 25544628.
- ^ Selden, P. A .; Shih, C. K.; Ren, D. (2013). "A giant spider from the Jurassic of China reveals greater diversity of the orbicularian stem group". Naturwissenschaften. 100 (12): 1171–1181. Bibcode:2013NW....100.1171S. doi:10.1007/s00114-013-1121-7. PMC 3889289. PMID 24317464.
- ^ Dunlop, Jason A .; Kamenz, Carsten; Scholtz, Gerhard (June 2007). "Reinterpreting the morphology of the Jurassic scorpion Liassoscorpionides". Struktura a vývoj členovců. 36 (2): 245–252. doi:10.1016/j.asd.2006.09.003. PMID 18089103.
- ^ Huang, Diying; Selden, Paul A .; Dunlop, Jason A. (August 2009). "Harvestmen (Arachnida: Opiliones) from the Middle Jurassic of China". Naturwissenschaften. 96 (8): 955–962. Bibcode:2009NW.....96..955H. doi:10.1007/s00114-009-0556-3. ISSN 0028-1042. PMID 19495718. S2CID 9570512.
- ^ Giribet, Gonzalo; Tourinho, Ana Lúcia; Shih, ChungKun; Ren, Dong (March 2012). "An exquisitely preserved harvestman (Arthropoda, Arachnida, Opiliones) from the Middle Jurassic of China". Organismy Rozmanitost a evoluce. 12 (1): 51–56. doi:10.1007/s13127-011-0067-x. ISSN 1439-6092. S2CID 15658216.
Zdroje
- Arkell, W.J. (1956). Jurassic Geology of the World. Edinburgh: Oliver & Boyd.
- Behrensmeyer, A.K .; Damuth, J.D.; DiMichele, W.A .; Potts, R.; Sues, H.D.; Wing, S.L., eds. (1992). Terrestrial Ecosystems through Time: the Evolutionary Paleoecology of Terrestrial Plants and Animals. Chicago: University of Chicago Press. ISBN 0-226-04154-9.
- Brongniart, Alexandre (1829). Tableau des terrains qui composent l'écorce du globe ou essai sur la structure de la partie connue de la terre [Description of the Terrains that Constitute the Crust of the Earth or Essay on the Structure of the Known Lands of the Earth] (francouzsky). Strasbourg – via Gallica.
- Carroll, R. L. (1988). Paleontologie a evoluce obratlovců. New York: WH Freeman. ISBN 978-0-7167-1822-2.
- Feduccia, A. (1996). Původ a vývoj ptáků. New Haven: Yale University Press. ISBN 978-0-300-06460-5.
- Gannon, Megan (31 October 2012). "V Číně byl nalezen hřbitov Jurských želv". cbsnews.com. Citováno 10. října 2019.
- Haines, Tim (2000). Walking with Dinosaurs: A Natural History. New York: Dorling Kindersley. ISBN 978-0-7894-5187-3.
- Hallam, A. (1986). "The Pliensbachian and Tithonian extinction events". Příroda. 319 (6056): 765–768. Bibcode:1986Natur.319..765H. doi:10.1038/319765a0. S2CID 4310433.
- Hölder, H. (1964). Jura – Handbuch der stratigraphischen Geologie (v němčině). IV. Stuttgart: Enke-Verlag.
- Jacobs, Louis, L. (1997). "African Dinosaurs". In Currie, Phillip J .; Padian, Kevin (eds.). Encyklopedie dinosaurů. Akademický tisk.
- Jaramillo, Jessica (March–April 2014). „Entrevista al Dr. Víctor Alberto Ramos, Premio México Ciencia y Tecnología 2013“ (ve španělštině). Sv. 17 č. 66.
Si logramos publicar esos nuevos resultados, sería el primer paso para cambiar formalmente la edad del Jurásico-Cretácico. Interní mezinárodní spolupráce v Ciencias Geológicas y la Comisión Internacional International Estratigrafía certificatería o no, depende de los resultados, ese cambio.
Cite magazine vyžaduje| časopis =(Pomoc) - Kazlev, M. Alan (2002). "Palaeos Mesozoic: Jurassic: The Jurassic Period". Palaeos. Archivovány od originál dne 5. ledna 2006. Citováno 8. ledna 2006.
- Pieńkowski, G.; Schudack, M.E.; Bosák, P.; Enay, R.; Feldman-Olszewska, A.; Golonka, J .; Gutowski, J.; et al. (2008). "Jurassic". In McCann, T. (ed.). The Geology of Central Europe. Mesozoic and Cenozoic. London: Geologická společnost.
- Rollier, L. (1903). "Das Schweizerische Juragebirge". Sonderabdruck aus dem Geographischen Lexikon der Schweiz (v němčině). Attinger, Neuenburg: von Gebr.
- Monroe, James S .; Wicander, Reed (1997). Měnící se Země: Zkoumání geologie a evoluce (2. vyd.). Belmont: West Publishing Company. ISBN 0-314-09577-2.
- Motani, R. (2000). "Rulers of the Jurassic Seas". Scientific American. Sv. 283 č. 6. doi:10.1038/scientificamerican1200-52.
- Scotese, Christopher R. (2003). "Pangea Begins to Rift Apart". scotese.com. Citováno 2019-10-10.
- Stanley, S.M .; Hardie, L.A. (1998). "Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry". Paleogeografie, paleoklimatologie, paleoekologie. 144 (1–2): 3–19. Bibcode:1998PPP ... 144 .... 3S. doi:10.1016/s0031-0182(98)00109-6.
- Taylor, P.D .; Wilson, M.A. (2003). "Palaeoecology and evolution of marine hard substrate communities" (PDF). Recenze vědy o Zemi. 62 (1): 1–103. Bibcode:2003ESRv ... 62 .... 1T. doi:10.1016/s0012-8252(02)00131-9. Archivovány od originál (PDF) dne 25.03.2009.
- Vennari, Verónica V .; Lescano, Marina; Naipauer, Maximiliano; Aguirre-Urreta, Beatriz; Concheyro, Andrea; Schaltegger, Urs; Armstrong, Richard; Pimentel, Marcio; Ramos, Victor A. (2014). „Nová omezení na hranici jury a křídy ve Vysokých Andách s použitím vysoce přesných dat U – Pb“. Výzkum v Gondwaně. 26 (1): 374–385. Bibcode:2014GondR..26..374V. doi:10.1016 / j.gr.2013.07.005.
- Vinn, O .; ten Hove, HA; Mutvei, H. (2008). „O ultrastruktuře trubice a původu kalcifikace u sabellidů (Annelida, Polychaeta)“. Paleontologie. 51 (2): 295–301. doi:10.1111 / j.1475-4983.2008.00763.x.
- Vinn, O .; Mutvei, H. (2009). "Calcareous tubeworms of the Phanerozoic" (PDF). Estonian Journal of Earth Sciences. 58 (4): 286–296. doi:10.3176/earth.2009.4.07. Vichřice A216178929.
- von Humboldt, Alexander (1799). Ueber die unterirdischen Gasarten und die Mittel, ihren Nachteil zu vermindern, ein Beitrag zur Physik der praktischen Bergbaukunde [On the types of subterranean gases and means of minimizing their harm, a contribution to the physics of practical mining] (v němčině). Braunschweig: Vieweg.
- von Humboldt, Alexander (1858). Kosmos (v němčině). 4. Stuttgart: Cotta. p. 632 – via HathiTrust.
- Wings, Oliver; Rabi, Márton; Schneider, Jörg W .; Schwermann, Leonie; Sun, Ge; Zhou, Chang-Fu; Joyce, Walter G. (2012). "An enormous Jurassic turtle bone bed from the Turpan Basin of Xinjiang, China". Naturwissenschaften. 114 (11): 925–935. Bibcode:2012NW ..... 99..925W. doi:10.1007 / s00114-012-0974-5. PMID 23086389. S2CID 17423081.
- Witton, Mark P.; Martill, David M .; Loveridge, Robert F. (2010). "Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity". Acta Geoscientica Sinica. 31 (Supp 1): 79–81.
- Witton, Mark P. (23 June 2016). "Why the giant azhdarchid Arambourgiania philadelphiae needs a fanclub". markwitton-com.blogspot.com.
Další čtení
- Mader, Sylvia (2004). Biologie (osmé vydání).
- Ogg, Jim (červen 2004). Overview of Global Boundary Stratotype Sections and Points (GSSP's). Mezinárodní komise pro stratigrafii. p. 17.
- Stanley, S.M .; Hardie, L.A. (1999). "Hypercalcification; paleontology links plate tectonics and geochemistry to sedimentology". GSA dnes. 9: 1–7.
externí odkazy
- Examples of Jurassic Fossils
- Jurassic (chronostratigraphy scale)
- Jurassic fossils in Harbury, Warwickshire
- Jurassic Microfossils: 65+ images of Foraminifera
- . Encyklopedie Britannica. 15 (11. vydání). 1911. With map and table.