Ostrovní nanismus - Insular dwarfism - Wikipedia

Ostrovní nanismus, forma fyletický nanismus,[1] je proces a stav velkých zvířat, která se vyvíjejí nebo mají zmenšenou velikost těla[A] když je rozsah jejich populace omezen na malé prostředí, zejména ostrovy. Tento přirozený proces se liší od záměrného vytváření trpasličích plemen, tzv trpaslík. K tomuto procesu došlo mnohokrát v průběhu evoluční historie, včetně příkladů dinosauři, jako Europasaurus a moderní zvířata jako sloni a jejich příbuzní. Tento proces a další “ostrovní genetika „artefakty se mohou vyskytovat nejen na ostrovech, ale také v jiných situacích, kdy je ekosystém izolován od vnějších zdrojů a chovu. To může zahrnovat jeskyně, poušť oázy, izolovaná údolí a izolované hory ("nebe ostrovy Ostrovní nanismus je jedním z aspektů obecnějších „ostrovní efekt“ nebo „Fosterovo pravidlo“, což předpokládá, že když zvířata na pevnině kolonizují ostrovy, malé druhy mají tendenci vyvíjet se větší těla (ostrovní gigantismus ) a velké druhy mají tendenci vyvíjet menší těla.
Možné příčiny
Existuje několik navrhovaných vysvětlení mechanismu, který produkuje takový zakrslost.[3][4]
Jedním z nich je selektivní proces, při kterém přežijí pouze menší zvířata uvězněná na ostrově, protože jídlo pravidelně klesá na hraniční úroveň. Menší zvířata potřebují méně zdrojů a menší teritoria, a je tedy větší pravděpodobnost, že se dostanou přes hranici bodu, kdy pokles populace umožňuje dostatečné doplnění zdrojů potravy, aby přeživší mohli vzkvétat. Menší velikost je také výhodná z hlediska reprodukce, protože znamená kratší gestační období a generační časy.[3]
V tropech by měla být malá velikost termoregulace jednodušší.[3]
U býložravců poskytuje velká velikost výhody při zvládání jak konkurence, tak dravců, takže zmenšení nebo absence obou by usnadnila zakrslost; konkurence se jeví jako důležitější faktor.[4]
Mezi masožravci se považuje za hlavní faktor velikost a dostupnost zdrojů kořisti a konkurence se považuje za méně důležitou.[4] v tygří hadi, ostrovní nanismus se vyskytuje na ostrovech, kde je dostupná kořist omezena na menší velikosti, než jaké běžně berou hadi z pevniny. Vzhledem k tomu, že preference velikosti kořisti u hadů je obecně úměrná velikosti těla, mohou se malé hady lépe přizpůsobit tomu, aby si vzaly malou kořist.[5]
Dwarfismus vs. gigantismus
Inverzní proces, při kterém se malá zvířata chovaná na izolovaných ostrovech bez predátorů velkých suchozemských mas mohou stát mnohem většími než obvykle, se nazývá ostrovní gigantismus. Vynikajícím příkladem je dodo, jejichž předkové měli normální velikost holubi. Existuje také několik druhů obří krysy, jeden stále existuje, který koexistoval s oběma Homo floresiensis a trpaslík stegodonty na Flores.
Proces ostrovního převyšování může podle evolučních standardů probíhat relativně rychle. To je v kontrastu s nárůstem maximální velikosti těla, který je mnohem pozvolnější. Při normalizaci na generační délku bylo zjištěno, že maximální rychlost úbytku tělesné hmotnosti během ostrovního převyšování je více než 30krát větší než maximální přírůstek tělesné hmotnosti při desetinásobné změně u savců.[6] Tato nerovnost odráží skutečnost, že pedomorfismus nabízí relativně snadnou cestu k vývoji menší velikosti těla dospělého; na druhé straně je pravděpodobné, že vývoj větší maximální velikosti těla bude přerušen vznikem řady omezení, která musí být překonána evolučními inovacemi, než může proces pokračovat.[6]
Faktory ovlivňující rozsah zakrslosti
U býložravců i masožravců se zdá, že velikost ostrova, stupeň izolace ostrova a velikost kontinentálních druhů předků nemají pro stupeň zakrslosti zásadní přímý význam.[4] Když však vezmeme v úvahu pouze tělesné hmotnosti nedávných špičkových býložravců a masožravců, a včetně údajů z kontinentálních i ostrovních suchozemských mas, bylo zjištěno, že tělesné hmotnosti největších druhů v pevnině se zvětšují na velikost zemské masy s svahy asi 0,5 log (tělesná hmotnost / kg) na log (plocha půdy / km2).[7] Byly samostatné regresní čáry pro endotermický nejlepší dravci, ectothermic špičkoví predátoři, endotermní top býložravci a (na základě omezených údajů) ektotermní top býložravci, takže příjem potravy byl u špičkových býložravců 7 až 24krát vyšší než u špičkových predátorů a přibližně stejný pro endotermy a ektotermy stejných tropická úroveň (to vede k tomu, že ektotermy jsou 5 až 16krát těžší než odpovídající endotermy).[7]
Příklady
Non-avian dinosaurs
Uznání, že ostrovní nanismus by se mohl vztahovat na dinosaury, vzniklo díky práci Ferenc Nopcsa, maďarský aristokrat, dobrodruh, vědec a paleontolog. Nopcsa intenzivně studovala transylvánské dinosaury a všimla si, že jsou menší než jejich bratranci jinde na světě. Například objevil šest metrů dlouhý sauropody, skupina dinosaurů, která jinde běžně rostla na 30 metrů nebo více. Nopcsa odvodil, že oblast, kde byly nalezeny pozůstatky, byl ostrov, Ostrov Hațeg (nyní povodí Haţeg nebo Hatzeg v Rumunsko ) Během Druhohor éra.[8][9] Nopcsův návrh dinosaurského nanismu na ostrově Hațeg je dnes široce přijímán poté, co další výzkum potvrdil, že nalezené pozůstatky nepocházejí od mladistvých.[10]
Sauropody
| Příklad | Druh | Rozsah | Časové okno | Kontinentální příbuzný |
|---|---|---|---|---|
Ampelosaurus | A. atacis | Ibero -Armorican ostrov | Pozdní křída / Maastrichtian |  Nemegtosaurids |
 Europasaurus | E. holgeri | Dolní Sasko | Pozdní Jurassic / Střední Kimmeridgian |  Brachiosaurs |
 Magyarosaurus | M. dacus | Ostrov Hateg | Pozdní křída / Maastrichtian |  Rapetosaurus |
 Lirainosaurus[11] | L. astibiae | Ibero-Armorican Island | Pozdní křída | |
 Paludititan | P. nalatzensis | Ostrov Hateg | Pozdní křída / Maastrichtian |  Epachthosaurus |
jiný
| Příklad | Druh | Rozsah | Časové okno | Kontinentální příbuzný |
|---|---|---|---|---|
Langenbergův lom torvosaur (modrý) | Bezejmený | Dolní Sasko | Pozdní Jurassic / Střední Kimmeridgian | Torvosaurus |
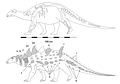 Struthiosaurus[12] | S. austriacus S. transylvanicus S. languedocensis | Ostrovy Ibero-Armorican, Australoalpine a Hateg | Pozdní křída |  Edmontonia |
 Telmatosaurus | T. transsylvanicus | Ostrov Hateg | Pozdní křída | Hadrosauridy |
 Tethyshadros | T. insularis | Terstská provincie | Pozdní křída | |
 Thecodontosaurus[9] | T. antiquus | Jižní Anglie | Pozdní trias / Rhaetian |  Plateosaurové |
 Zalmoxes[9] (nachový) | Z. robustus Z. shqiperorum | Ostrov Hateg | Pozdní křída |  Tenontosaurus |
Kromě toho rod Balaur byl původně popsán jako a Velociraptor - velikost dromaeosaurid (a v důsledku toho pochybný příklad ostrovního nanismu), ale od té doby byl překlasifikován na druhotně nelétavého kmenového ptáka, blíže moderním ptákům než Jeholornis (tedy vlastně příklad ostrovní gigantismus ).
Ptactvo
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
 havajský nelétavý ibises | Apteribis glenos | Molokai | Vyhynulý (Pozdě Kvartérní ) |  Americké ibises | |
| Apteribis brevis | Maui | ||||
| Cozumel curassow[13] | Crax rubra griscomi | Cozumel | Neznámý |  Velký curassow | |
 Klokaní ostrov emu[14] | Dromaius novaehollandiae baudinianus | Klokaní ostrov, Jižní Austrálie | Vyhynulý (kolem r. 1827) |  Emu | |
 King Island emu[15] (Černá) | Dromaius novaehollandiae minor | King Island, Tasmánie | Vyhynulý (AD 1822) | LR ≈ 0,48 [b] | |
 Cozumel Thrasher[13] | Toxostoma gluttatum | Cozumel | Kriticky ohrožený |  Ostatní mlátičky |
Squamates
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
 Madagaskarský trpasličí chameleon | Brookesia minima | Nosy Be ostrov, Madagaskar | Ohrožený |  Madagaskarské listové chameleony | |
 Nosy Hara chameleon[16] | Brookesia micra | Nosy Hara ostrov, Madagaskar | Zranitelný | ||
| Tygr hadího ostrova Roxby[5] | Notechis scutatus | Roxby Island, jižní Austrálie | Neznámý |  Tygří had | |
| Trpasličí barmský krajta | Python bivittatus progschai | Jáva, Bali, Sumbawa a Sulawesi, Indonésie | Neznámý |  Barmský krajta | LR ≈ 0,44 [C] |
| Tanahjampea síťovaný krajta[19] | Python reticulatus jampeanus | Tanahjampea, mezi Sulawesi a Flores | Neznámý |  Síťovaný python | LR ≈ 0,41, muži LR ≈ 0,49, ženy [d] |
Savci
Piloti
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
 Lenochod tříprstý | Bradypus pygmaeus | Isla Escudo de Veraguas, Panama | Kriticky ohrožený |  Lenost s hrdlem |
 Acratocnus | A. antillensis A. odontrigonus A. vy | Kuba, Hispaniola a Portoriko | Vyhynulý (c. 3000 BC) |  Kontinentální lenochodi |
| Imagocnus | I. zazae | Kuba | Vyhynulý (Raný miocén ) | |
 Megalocnus | M. rodens M. zile | Kuba a Hispaniola | Vyhynulý (kolem 2700 př. n. l.) | |
 Neoknus | Neoknus spp. | Kuba a Hispaniola | Vyhynulý (c. 3000 BC) |
Proboscideans
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
| Sulawesi trpasličí slon | Elephas celebensis | Sulawesi | Vyhynulý (Raný pleistocén) |  Asijský slon |
 Krétský trpasličí mamut | Mammuthus creticus | Kréta | Vyhynulý | Mammuthus |
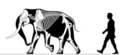 Normanské ostrovy mamut | Mammuthus exilis | Santa Rosae ostrov | Vyhynulý (Pozdní pleistocén ) | Columbian mamut |
| Sardinský mamut | Mammuthus lamarmorai | Sardinie | Vyhynulý (Pozdní pleistocén) |  Stepní mamut |
| Vlněný mamut ostrova Saint Paul[22][23] | Mammuthus primigenius | Ostrov svatého Pavla Aljaška | Vyhynulý (asi 3750 př. n. l.) | Vlněný mamut |
 Siculo-maltští sloni | Palaeoloxodon antiquus leonardi P. mnaidriensis P. melitensis P. falconeri | Sicílie a Malta | Vyhynulý |  Rovný kel (vlevo, odjet) |
| Krétské slony | Palaeoloxodon chaniensis P. creutzburgi | Kréta | Vyhynulý | |
 Kypr trpasličí slon | Palaeoloxodon cypriotes | Kypr | Vyhynulý (kolem 9000 př. n.l.) | |
| Naxos trpasličí slon | Palaeoloxodon sp. | Naxos | Vyhynulý | |
| Rhodos a Tilos trpasličí slon | Palaeoloxodon tiliensis | Rhodos a Tilos | Vyhynulý | |
| Bumiayu trpaslík sinomastodont[24] | Sinomastodon bumiajuensis | Ostrov Bumiayu (nyní součástí Jáva ) | Vyhynulý (Raný pleistocén) |  Sinomastodon |
 Japonský stegodont[25] | Stegodon aurorae | Japonsko a Tchaj-wan[26] | Vyhynulý (Raný pleistocén ) |  čínština Stegodon |
| Trpasličí stegodont větší Flores[3] | Stegodon florensis | Flores | Vyhynulý (Pozdní pleistocén) |  Sundaland Stegodon |
| Jávské trpasličí stegodonty | Stegodon hypsilophus[24] S. semedoensis[27] S. sp.[24] | Jáva | Vyhynulý (Kvartérní ) | |
| Mindanao trpasličí stegodont[28] | Stegodon mindanensis | Mindanao a Sulawesi | Vyhynulý (Střední pleistocén) | |
| Sulawesi trpasličí stegodont[24] | Stegodon sompoensis | Sulawesi | Vyhynulý | |
| Trpasličí stegodont z Malé Flores[3] | Stegodon sondaari | Flores | Vyhynulý (Střední pleistocén) | |
| Sumba trpaslík stegodont[29] | Stegodon sumbaensis | Sumba, Indonésie | Vyhynulý (Střední pleistocén) | |
| Trpasličí trpaslík stegodont[24] | Stegodon timorensis | Timor | Vyhynulý | |
| Trpasličí stegolophodont[30] | Stegolophodon pseudolatidens | Japonsko | Vyhynulý (Miocén ) |  Stegolophodon |
Primáti
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
| Nosy Hara trpasličí lemur[31] | Cheirogaleus sp. | Nosy Hara ostrov u Madagaskaru | Neznámý |  Trpasličí lemury |
 Flores Man[32] | Homo floresiensis | Flores | Vyhynulý (Pozdní pleistocén) |  Homo erectus |
 Callao Man | Homo luzonensis[33][34] | Luzon, Filipíny | Vyhynulý (Pozdní pleistocén) | |
| Moderní Pygmejové z Floresu[35] | Homo sapiens | Flores | Existující |  Homo sapiens |
| Early Palau moderní lidé (sporné)[36] | Homo sapiens | Palau | Zaniklý (?) | |
 Andamanský[37][38] | Homo sapiens | Andamanské ostrovy | Existující | |
 Sardinský makak[39] | Macaca majori | Sardinie | Vyhynulý (Pleistocén) |  Barbary makak |
 Zanzibarský červený colobus | Piliocolobus kirkii | Unguja | Ohrožený |  Udzungwa červený colobus |
Carnivorans
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
 Japonský vlk | Canis lupus hodophilax | Japonsko (kromě Hokkaido ) | Vyhynulý (1905 nl) |  šedý vlk | |
 Sardinský dhole (vpřed) | Cynotherium sardous | Korsika a Sardinie | Vyhynulý (asi 8300 př. n.l.) |  Xenocyon | |
| Trinil pes | Mececyon trinilensis | Jáva | Vyhynulý (Pleistocén) | ||
| Nosál Cozumel Island[13] | Nasua narica nelsoni | Cozumel | Kriticky ohrožený |  Yucatan nosál bílý | |
 Zanzibarský leopard | Panthera pardus pardus | Unguja | Kriticky ohrožený nebo vyhynulý |  Africký leopard | |
 Bali tygr | Panthera tigris sondaica | Bali | Vyhynulý (kolem r. 1940) |  Tygr sumaterský | |
 Javanský tygr | Jáva | Vyhynulý (c. AD 1975) | |||
 Mýval Cozumel | Procyon pygmaeus | Cozumel | Kriticky ohrožený |  Společný mýval | |
 Ostrov liška | Urocyon littoralis | Šest z Normanské ostrovy v Kalifornii | Blízko ohrožení |  Šedá liška | LR ≈ 0,84 [E] LR ≈ 0,75 [F] |
| Liška Cozumel | Urocyon sp. | Cozumel | Kriticky ohrožený nebo vyhynulý |
Nepřežvýkavec kopytníci
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
 Madagaskarské trpasličí hrochy | Hippopotamus laloumena H. lemerlei H. madagascariensis | Madagaskar | Vyhynulý (asi 1000 nl) |  Společný hroch |
| Bumiayu trpasličí hroch[24] | Hexaprotodon simplex | Ostrov Bumiayu (nyní Jáva ) | Vyhynulý (Raný pleistocén ) |  Asijské hrochy |
 Krétský trpasličí hroch | Hippopotamus creutzburgi | Kréta | Vyhynulý (Střední pleistocén ) |  Evropský hroch |
 Maltézský trpasličí hroch | Hippopotamus melitensis | Malta | Vyhynulý (Pleistocén) | |
 Kypr trpasličí hroch | Hippopotamus minor | Kypr | Vyhynulý (kolem 8000 př. n.l.) | |
 Sicilský trpasličí hroch | Hippopotamus pentlandi | Sicílie | Vyhynulý (Pleistocén) | |
| Cozumel s límečkem[13] | Pecari tajacu nanus | Cozumel | Neznámý |  Límečkem pekari |
| Filipínský nosorožec[42] | Rhinoceros philippinensis | Luzon | Vyhynulý (Střední pleistocén) |  Nosorožec jávský |
Bovids
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
| Sicilský bizon[25] | Bison priscus siciliae | Sicílie | Vyhynulý (Pozdní pleistocén) |  Stepní bizon |
| Sicilské zubry[43] | Bos primigenius siciliae[25] | Sicílie | Vyhynulý (Pozdní pleistocén) |  Euroasijský zubr |
| Cebu tamaraw | Bubalus cebuensis | Cebu, Filipíny | Vyhynulý |  Divoký vodní buvol |
 Dolní anoa | Bubalus depressicornis | Sulawesi a Buton, Indonésie | Ohrožený | |
 Tamaraw | Bubalus mindorensis | Mindoro, Filipíny | Kriticky ohrožený | |
 Hora anoa | Bubalus quarlesi | Sulawesi a Buton, Indonésie | Ohrožený | |
 Jeskynní koza na Baleárských ostrovech | Myotragus balearicus | Mallorca a Menorca | Vyhynulý (po roce 3000 př. n.l.) | Gallogoral |
| Nesogoral[44] | Nesogoral spp. | Sardinie | Vyhynulý | |
| Dahlak Kebir gazela[45] | Nanger soemmerringi ssp. | Dahlak Kebir ostrov, Eritrea | Zranitelný |  Soemmerringova gazela |
Cervids a příbuzní
| Příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
 Krétské trpasličí megaceriny[G] | Candiacervus spp. | Kréta | Vyhynulý (Pleistocén) |  Praemegaceros verticornis[9] |
 Sardinský megacerin[9] (druhý zleva) | Praemegaceros cazioti | Sardinie | Vyhynulý (asi 5500 př. n. l.) | |
 Ryukyu trpasličí jelen[48] | Cervus astylodon | Rjúkjú | Vyhynulý |  Sika jelen (?) Cervus praenipponicus (?) |
| Populace jelenů Jersey[49] | Cervus elaphus jerseyensis | Trikot | Vyhynulý (Pleistocén) |  Jelen |
 Korsický jelen | Cervus elaphus corsicanus | Korsika a Sardinie | Blízko ohrožení | |
| Pleistocénní sicilský jelen[25] | Cervus siciliae | Sicílie | Vyhynulý (Pozdní pleistocén) | |
 Hoplitomeryx[h] | Hoplitomeryx spp. | Ostrov Gargano | Vyhynulý (Časný pliocén ) |  Pecorans |
| Sicilský megacerin[25] | Megaloceros carburangelensis | Sicílie | Vyhynulý (Pozdní pleistocén) |  Irský los |
 Florida Key jelen | Odocoileus virginianus clavium | Florida Keys | Ohrožený |  Virginský jelen |
 Svalbardský sob | Rangifer tarandus platyrhynchus | Špicberky | Neznámý |  Sob |
 Filipínský jelen | Rusa marianna | Filipíny | Zranitelný |  Sambar jelen |
Rostliny
| Možný příklad | Binomické jméno | Nativní rozsah | Postavení | Kontinentální příbuzný |
|---|---|---|---|---|
 Ostrovní sloní kaktusy[50][51] | Pachycereus pringlei | Vzdálené ostrovy v Cortezovo moře (např. Santa Cruz, San Pedro Mártir ) | Neohodnocený |  Sloní kaktusy na pevnině |
Viz také
Poznámky
- ^ Příkladem neizolovaného fyletického nanismu je vývoj trpaslíků kosmani a tamaríny mezi opicemi Nového světa, které vyvrcholily objevením nejmenšího příkladu, Cebuella pygmaea.[2]
- ^ Na základě výšek na obr. 1 Heupink et al., 2011[15]
- ^ Založeno na maximální délce 2,5 m pro trpasličí formu[17] a 5,74 m pro pevninskou formu[18]
- ^ Na základě maximální celkové délky (TL) Tanahjampea python 2,10 m pro muže a 3,35 m pro ženy[19] a maximální čenich jižní Sumatry python do délky větrání (SVL) 4,5 m u mužů a 6,1 m u žen[20] s SVL korigovanými na TL vynásobením faktorem 1,127, odvozeným z průměrné relativní délky ocasu (0,113) afrických a indických skalních pythonů[21]
- ^ Pro nedaleké pevniny šedé lišky[40]
- ^ Obecně pro lišky šedé[41]
- ^ Jako Hoplitomeryx, Candiacervus se jeví jako neobvyklý případ, kdy se členové tohoto rodu vyvinuli v ostrovní druhy širokého rozsahu velikostí, nejen trpasličí formy, ale také některé, které lze považovat za obry.[46][47]
- ^ Hoplitomeryx je evidentně docela neobvyklý případ, protože členové tohoto rodu se zjevně vyvinuli do trpasličích i obrovských ostrovních forem na stejném ostrově (ostrovech).[46]
Reference
- ^ Prothero, D. R .; Sereno, P. C. (zima 1982). „Allometrie a paleoekologie středních miocénních trpasličích nosorožců z pobřežní nížiny Texasského zálivu“. Paleobiologie. 8 (1): 16–30. doi:10.1017 / S0094837300004322. JSTOR 2400564.
- ^ Perelman, P .; et al. (2011). „Molekulární fylogeneze živých primátů“. Genetika PLOS. 7 (3): 1–17. doi:10.1371 / journal.pgen.1001342. PMC 3060065. PMID 21436896.
- ^ A b C d E Van Den Bergh, G. D .; Rokhus Due Awe; Morwood, M. J .; Sutikna, T .; Jatmiko; Wahyu Saptomo, E. (květen 2008). "Nejmladší Stegodon zůstává v jihovýchodní Asii z pozdního pleistocénního archeologického naleziště Liang Bua, Flores, Indonésie ". Kvartérní mezinárodní. 182 (1): 16–48. Bibcode:2008QuInt.182 ... 16V. doi:10.1016 / j.quaint.2007.02.001.
- ^ A b C d Raia, P .; Meiri, S. (srpen 2006). „Vládne ostrov u velkých savců: paleontologie se setkává s ekologií“. Vývoj. 60 (8): 1731–1742. doi:10.1111 / j.0014-3820.2006.tb00516.x. PMID 17017072. S2CID 26853128.
- ^ A b Keogh, J. S .; Scott, I. A. W .; Hayes, C. (leden 2005). „Rychlý a opakovaný původ ostrovního gigantismu a nanismu u australských tygřích hadů“. Vývoj. 59 (1): 226–233. doi:10.1111 / j.0014-3820.2005.tb00909.x. PMID 15792242. S2CID 58524.
- ^ A b Evans, A. R .; et al. (2012-01-30). „Maximální rychlost vývoje savců“. PNAS. 109 (11): 4187–4190. Bibcode:2012PNAS..109,4187E. doi:10.1073 / pnas.1120774109. PMC 3306709. PMID 22308461. Citováno 2011-02-11.
- ^ A b Burness, G. P .; Diamond, J.; Flannery, T. (2001-12-04). „Dinosauři, draci a trpaslíci: vývoj maximální velikosti těla“. Sborník Národní akademie věd. 98 (25): 14518–14523. Bibcode:2001PNAS ... 9814518B. doi:10.1073 / pnas.251548698. ISSN 0027-8424. JSTOR 3057309. PMC 64714. PMID 11724953.
- ^ „Trpasličí ostrov dinosaurů skutečně existoval, tvrdí vědci“. Telegraph Media Group. 2010-02-22. Citováno 2010-02-26.
- ^ A b C d E Benton, M. J .; Csiki, Z .; Grigorescu, D .; Redelstorff, R .; Sander, P. M .; Stein, K .; Weishampel, D. B. (2010-01-28). „Dinosauři a ostrov vládnou: Trpasličí dinosauři z ostrova Haţeg“ (PDF). Paleogeografie, paleoklimatologie, paleoekologie. 293 (3–4): 438–454. Bibcode:2010PPP ... 293..438B. doi:10.1016 / j.palaeo.2010.01.026. Archivovány od originál (PDF) dne 10.7.2011. Citováno 2017-07-30.
- ^ Dyke, G. (2011-09-20). „Dinosauří baron Transylvánie“. Scientific American. 305 (4): 80–83. Bibcode:2011SciAm.305c..80D. doi:10.1038 / scientificamerican1011-80. PMID 22106812.
- ^ Company, J. (2010). „Histologie kostí titanosaura Lirainosaurus astibiae (Dinosauria: Sauropoda) z poslední křídy Španělska “. Naturwissenschaften. 98 (1): 67–78. doi:10.1007 / s00114-010-0742-3. hdl:10251/148874. PMID 21120450. S2CID 31752413.
- ^ Carpenter, K. (2001) Obrněné dinosaury. Indiana University Press, 526 stran.
- ^ A b C d Cuarón, A. D .; Martínez-Morales, M. A .; McFadden, K. W .; Valenzuela, D .; Gompper, M. E. (2004). "Stav trpasličích masožravců na ostrově Cozumel v Mexiku". Biodiverzita a ochrana. 13 (2): 317–331. CiteSeerX 10.1.1.511.2040. doi:10.1023 / b: bioc.0000006501.80472.cc. S2CID 25730672.
- ^ Parker S (1984) Zaniklý ostrov Klokan Emu, dosud neuznaný druh. Bulletin klubu britských ornitologů 104: 19–22.
- ^ A b Heupink, T. H .; Huynen, L .; Lambert, D. M. (2011). „Starověká DNA naznačuje, že trpaslík a„ obří “emu jsou shodné“. PLOS ONE. 6 (4): e18728. Bibcode:2011PLoSO ... 618728H. doi:10.1371 / journal.pone.0018728. PMC 3073985. PMID 21494561.
- ^ Glaw, F .; Köhler, J .; Townsend, T. M .; Vences, M. (2012-02-14). „Soupeřit s nejmenšími plazy na světě: Objev miniaturizovaných a mikroendemických nových druhů chameleonů listových (Brookesia) ze severního Madagaskaru ". PLOS ONE. 7 (2): e31314. Bibcode:2012PLoSO ... 731314G. doi:10.1371 / journal.pone.0031314. PMC 3279364. PMID 22348069.
- ^ de Lang R, Vogel G (2005). The Snakes of Sulawesi: A Field Guide to the Land Snakes of Sulawesi with Identification Keys. Frankfurtské příspěvky do skupiny Natural History Band 25, vydání Chimaira 2005. ISBN 3-930612-85-2. 23-27, 198-201.
- ^ Barker, D.G .; Barten, S.L .; Ehrsam, J.P .; Daddono, L. (2012). „Opravené délky dvou známých obřích Pythonů a stanovení nového záznamu maximální délky pro barmské Pythony, Python bivittatus" (PDF). Bulletin of the Chicago Herpetological Society. 47 (1): 1–6. Citováno 2020-03-02.
- ^ A b Auliya, M .; Mausfeld, P .; Schmitz, A .; Böhme, W. (04.04.2002). "Recenze síťovaného krajta (Python reticulatus Schneider, 1801) s popisem nového poddruhu z Indonésie “. Naturwissenschaften. 89 (5): 201–213. Bibcode:2002NW ..... 89..201A. doi:10.1007 / s00114-002-0320-4. PMID 12135085. S2CID 4368895.
- ^ Shine, R .; Harlow, P.S .; Keogh, J.S .; Boeadi, N.I. (1998). „Vliv pohlaví a velikosti těla na stravovací návyky obřího tropického hada, Python reticulatus ". Funkční ekologie. 12 (2): 248–258. doi:10.1046 / j.1365-2435.1998.00179.x.
- ^ Sheehy, C.M .; Albert, J.S .; Lillywhite, H.B .; Van Damme, R. (2016). „Vývoj délky ocasu u hadů spojený s různými gravitačními prostředími“. Funkční ekologie. 30 (2): 244–254. doi:10.1111/1365-2435.12472.; viz tabulka S1
- ^ Schirber, Michael. Přežít vyhynutí: Kde vydrželi vlní mamuti. Živá věda. Společnost Imaginova Corporation. Citováno 2007-07-20.
- ^ Mamuti z Ostrov Wrangel, severně od Sibiř, již nejsou považováni za trpaslíky. Viz: Tichonov, Alexej; Larry Agenbroad; Sergey Vartanyan (2003). Srovnávací analýza populací mamutů na Wrangel Island a Normanských ostrovech. DEINSEA 9: 415–420. ISSN 0923-9308
- ^ A b C d E F Aziz, F .; van den Bergh, G. D. (25. září 1995). „Trpaslík Stegodon ze Sambungmacanu (Střední Jáva, Indonésie) ". Proc. Kon. Ned. Akad. V. Wetensch. 98 (3): 229–241. Citováno 2017-07-31.
- ^ A b C d E Sondaar, P. Y .; A.A.E. van der Geer (2005). „Evoluce a zánik kopytníků na pleistocénním ostrově“. International Journal of the French Quaternary Association. 2: 241–256. Citováno 2017-07-31.
- ^ http://www.rhinoresourcecenter.com/pdf_files/129/1291330178.pdf
- ^ Siswanto, S., & Noerwidi, S. (2014). PROBOSCIDEA FOSSIL Z WEBU SEMEDO: Jeho korelace s biostratigrafií a příchodem člověka v Javě. Berkala Arkeologi, 34 (2).
- ^ Zaim, Y. (20. srpna 2010). „Geologické důkazy pro nejranější výskyt homininů v Indonésii“. In Fleagle, J. G; Shea, J. J .; Grine, F. E .; Baden, A. L .; Leakey, R. E. (eds.). Out of Africa I: The First Hominin Colonization of Eurasia. Springer Science & Business Media. p. 106. ISBN 978-90-481-9036-2. OCLC 668096676.
- ^ http://ro.uow.edu.au/cgi/viewcontent.cgi?article=3055&context=smhpapers
- ^ Saegusa, H. (2008). "Trpaslík Stegolophodon z japonského miocénu: cestující na potápějících se člunech “. Kvartérní mezinárodní. 182 (1): 49–62. Bibcode:2008QuInt.182 ... 49S. doi:10.1016 / j.quaint.2007.08.001.
- ^ „Nová skupina trpasličích lemurů může být nejvzácnějším primátem na světě“.
- ^ Vědec studující morbitování Hobita, abc.net.au
- ^ Wade, L. (10.04.2019). "Nový druh starověkého člověka objevený na Filipínách". Věda. 364. doi:10.1126 / science.aax6501.
- ^ Détroit, F .; Mijares, A. S .; Corny, J .; Daver, G .; Zanolli, C .; Dizon, E .; Robles, E .; Grün, R .; Piper, P. J. (2019). "Nový druh Homo z pozdního pleistocénu na Filipínách “. Příroda. 568 (7751): 181–186. Bibcode:2019Natur.568..181D. doi:10.1038 / s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Tucci, S .; et al. (03.08.2018). „Evoluční historie a adaptace lidské trpasličí populace ostrova Flores v Indonésii“. Věda. 361 (6401): 511–516. Bibcode:2018Sci ... 361..511T. doi:10.1126 / science.aar8486. PMC 6709593. PMID 30072539.
- ^ "Staří malí lidé na Palau, nikoli trpaslíci, uvádí studie ". Zprávy z National Geographic. 27. srpna 2008.
- ^ Gibbons, A. (2018). „Ostrovní život může zmenšit člověka“. Věda. doi:10.1126 / science.aau9750.
- ^ Mondal, M .; Casals, F .; Xu, T .; Dall'Olio, G. M .; Pybus, M .; Netea, M. G .; Comas, D .; Laayouni, H .; Li, Q .; Majumder, P. P .; Bertranpetit, J. (2016). „Genomická analýza andamanštiny poskytuje pohled na migraci starých lidí do Asie a adaptaci“ (PDF). Genetika přírody. 48 (9): 1066–1070. doi:10,1038 / ng. 3621. hdl:10230/34401. PMID 27455350. S2CID 205352099.
- ^ Rook, L. (2008-12-31). „První workshop o evropských záznamech o fosilních primátech (Siena a Grosseto, 11. – 13. Září 2008) s aktualizací italských studií paleoprimatologie“ (PDF). Atti Muss. Stor. Nat. Maremma (22): 129–143.
- ^ Parfit, M .; Groo, M. (22. dubna 2020). „Povznášející příběh těchto malých ostrovních lišek, téměř zničených katastrofou“. NationalGeographic.com. národní geografie. Citováno 2020-04-23.
- ^ Moore, C.M .; Collins, P.W. (1995). „Savčí druhy - Urocyon littoralis" (PDF). 489: 1–7. Archivovány od originál (PDF) dne 22. ledna 2012. Citováno 16. září 2011. Citovat deník vyžaduje
| deník =(Pomoc) - ^ Renema, Willem (2007). Biogeografie, čas a místo: distribuce, bariéry a ostrovy. Springer Science & Business Media. p. 334. ISBN 978-1-4020-6374-9. OCLC 228153573.
- ^ van Vuure, Cis (2005). Stopování Aurochs: Historie, morfologie a ekologie vyhynulého divokého vola. Coronet Books Incorporated. ISBN 978-954-642-235-4. OCLC 472741798.
- ^ van der Geer, A .; Lyras, G; de Vos, J .; Dermitzakis, M. (14. února 2011). „Sardinie a Korsika“. Evoluce ostrovních savců: Adaptace a vyhynutí placentárních savců na ostrovech. John Wiley & Sons. ISBN 978-1-4443-9128-2. OCLC 894698082.
- ^ Chiozzi, G .; Bardelli, G .; Ricci, M .; De Marchi, G .; Cardini, A. (2014). „Jenom další ostrovní trpaslík? Fenotypová rozlišovací schopnost ve špatně známé Soemmerringově Gazelle, Nanger soemmerringii (Cetartiodactyla: Bovidae), z ostrova Dahlak Kebir ". Biologický žurnál společnosti Linnean. 111 (3): 603–620. doi:10.1111 / bij.12239.
- ^ A b Mazza, P.P.A .; Rossi, M. A.; Agostini, S. (2015). „Hoplitomerycidae (pozdní miocén, Itálie), příklad gigantismu u ostrovních přežvýkavců“. Journal of Mammalian Evolution. 22 (2): 271–277. doi:10.1007 / s10914-014-9277-2. S2CID 16437411.
- ^ van der Geer, A.A.E. (2018). „Jednotnost v odrůdě: morfologie a vývoj parohu v prostředí bez predátorů“. Palaeontologia Electronica (21.1.9A): 1–31. doi:10.26879/834.
- ^ Kaifu, Y .; Fujita, M .; Yoneda, M .; Yamasaki, S. (15. února 2015). "Pleistocene Seafaring and Colonization of the Ryukyu Islands, Southwestern Japan". V Kaifu, Y .; Izuho, M .; Goebel, T .; Sato, H .; Ono, A. (eds.). Vznik a rozmanitost moderního lidského chování v paleolitické Asii. Texas A&M University Press. ISBN 978-1-62349-277-9. OCLC 985023261.
- ^ Lister, A. M. (1989-11-30). „Rychlý trpasličí jelen na Jersey v posledním meziglaciálu“. Příroda. 342 (6249): 539–542. Bibcode:1989 Natur.342..539L. doi:10.1038 / 342539a0. PMID 2685610. S2CID 4343091.
- ^ Wilder, B.T .; Felger, R.S. (30. září 2010). „Dwarf Giants, Guano, and Isolation: Vegetation and Floristic Diversity of San Pedro Mártir Island, Gulf of California, Mexico“ (PDF). Sborník San Diego Society of Natural History. 42: 1–24, viz str. 9–13. Citováno 2020-01-05.
(str. 12) Trpaslík rostlin San Pedro Mártir se zdá být způsoben výběrem kratších jedinců, kteří přežijí divoké tropické bouře, možnou konkurencí kořenů v tak hustém lese a nedefinovaným účinkem vysokých hladin dusíku a fosforu hojné guano, které by mohlo zastavit růst. Genetické studie nebyly provedeny ...
- ^ Burns, K.C. (Květen 2019). Evoluce v izolaci: Hledání ostrovního syndromu u rostlin. Cambridge University Press. 174–177. doi:10.1017/9781108379953. ISBN 978-1108379953. OCLC 1108160200.
(str. 174–175) ... rozsah, v jakém je jeho zakrslá postava geneticky podmíněna, a vysvětlení, proč by ostrovní zakrslost mohla být selektivně výhodná, čeká na další studii.
externí odkazy
| Pravidla |
|  |
|---|---|---|
| Příbuzný |
| |