Neurospora crassa - Neurospora crassa
| Neurospora crassa | |
|---|---|
 | |
| Vědecká klasifikace | |
| Království: | |
| Kmen: | |
| Podkmen: | |
| Třída: | |
| Objednat: | |
| Rodina: | |
| Rod: | |
| Druh: | N. crassa |
| Binomické jméno | |
| Neurospora crassa Stříhat & B.O. Vyhnout se | |
Neurospora crassa je druh formy červeného chleba kmene Ascomycota. Jméno rodu, což v řečtině znamená „nervová spora“, odkazuje na charakteristické proužky na výtrusy. První zveřejněná zpráva o této houbě pocházela z napadení francouzských pekáren v roce 1843.[1]
N. crassa se používá jako modelový organismus protože se snadno pěstuje a má haploidní životní cyklus, který dělá genetický analýza jednoduchá, protože u potomků se projeví recesivní rysy. Analýzu genetické rekombinace usnadňuje uspořádané uspořádání produktů meiózy v Neurospora askospory. Celý genom sedm chromozomů bylo sekvenováno.[2]
Neurospora byl používán uživatelem Edward Tatum a George Wells Beadle ve svých experimentech, za které vyhráli Nobelova cena za fyziologii nebo medicínu v roce 1958. Beadle a Tatum vystaveni N. crassa na rentgenové záření, způsobující mutace. Poté pozorovali selhání v metabolické cesty způsobené konkrétními chybami enzymy. To je vedlo k tomu, aby navrhli hypotéza „jeden gen, jeden enzym“ to konkrétní geny kód pro konkrétní bílkoviny. Jejich hypotéza byla později vyvinuta na enzymové dráhy Norman Horowitz, také pracuje na Neurospora. Jak Norman Horowitz vzpomínal v roce 2004,[3] „Tyto experimenty založily vědu o tom, čemu Beadle a Tatum říkali„ biochemická genetika “. Ve skutečnosti se ukázaly jako úvodní zbraň v tom, co se stalo molekulární genetikou a veškerým vývojem, který z toho vyplynul.“
Ve vydání ze dne 24. Dubna 2003 Příroda, genom z N. crassa byl označen jako úplně seřazeno.[4] Genom je dlouhý asi 43 megabází a zahrnuje přibližně 10 000 genů. Probíhá projekt produkce kmenů obsahujících knokaut mutanti každého N. crassa gen.[5]
Ve svém přirozeném prostředí N. crassa žije hlavně v tropických a subtropických oblastech.[6] Lze jej nalézt rostoucí na mrtvé rostlinné hmotě po požárech.
Neurospora se aktivně používá ve výzkumu po celém světě. Je to důležité při objasňování molekulárních událostí cirkadiánní rytmy, epigenetika a umlčení genů, polarita buněk, buněčná fúze, vývoj, stejně jako mnoho aspektů buněčné biologie a biochemie.
Sexuální cyklus
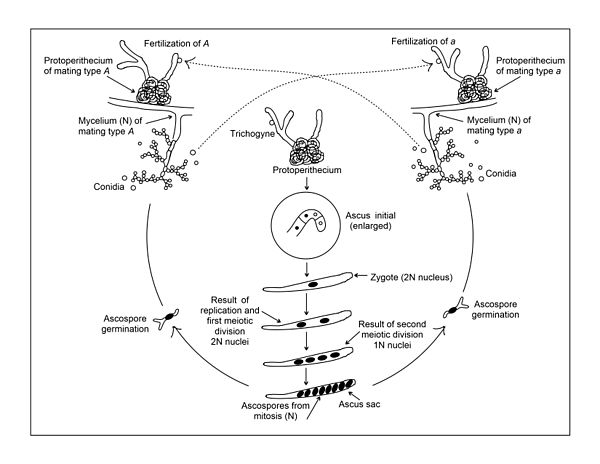
Sexuální plodnice (perithecia) lze vytvořit pouze tehdy, když se spojí dvě mycelia různého typu páření (viz obrázek). Stejně jako ostatní Ascomycetes, N. crassa má dva typy páření, které jsou v tomto případě symbolizovány A a A. Mezi. Není patrný morfologický rozdíl A a A kmeny typu páření. Oba mohou tvořit hojnou protoperithecii, ženskou reprodukční strukturu (viz obrázek). Protoperithecie se nejsnadněji tvoří v laboratoři, když dochází k růstu na pevném (agarovém) syntetickém médiu s relativně nízkým zdrojem dusíku.[7] Hladování dusíkem se jeví jako nezbytné pro expresi genů podílejících se na sexuálním vývoji.[8] Protoperithecium se skládá z ascogonia, stočeného mnohobuněčného hyfy, která je uzavřena v uzlovité agregaci hyf. Rozvětvený systém štíhlých hyf, nazývaných trichogyny, se táhne od špičky ascogonia vyčnívající za oplášťující hyfy do vzduchu. Sexuální cyklus je zahájen (tj. Dochází k oplodnění), když se buňka (obvykle konidium) opačného páření dotkne části trichogynu (viz obrázek). Po takovém kontaktu může následovat buněčná fúze vedoucí k tomu, že jedno nebo více jader z oplodňující buňky migruje dolů trichogynem do ascogonia. Protože oba A a A kmeny mají stejné sexuální struktury, žádný kmen nelze považovat za výlučně mužský nebo ženský. Jako příjemce však protoperithecium obou A a A kmeny lze považovat za ženskou strukturu a plodící konidium lze považovat za mužského účastníka.
Následující kroky následující po fúzi A a A haploidní buňky, popsal Fincham a Day[9] a Wagner a Mitchell.[10] Po fúzi buněk je další fúze jejich jader zpožděna. Místo toho se jádro z oplodňující buňky a jádro z ascogonia spojí a začnou se synchronně dělit. Produkty těchto jaderných divizí (stále v párech na rozdíl od páření, tj. A / a) migrují do mnoha askogenních hyf, které pak začnou růst z ascogonia. Každý z těchto askogenních hyf se ohýbá, aby vytvořil háček (nebo crozier) na špičce a A a A dvojice haploidních jader uvnitř crozierova rozdělení se synchronizuje. Dále se tvoří septa, která rozdělují crozier na tři buňky. Centrální buňka v křivce háku obsahuje jednu A a jeden A jádro (viz obrázek). Tato dvoujaderná buňka iniciuje tvorbu asku a nazývá se buňka „ascus-initial“. Další dva uninucleate buňky na obou stranách první buňky vytvářející ascus se navzájem fúzují a tvoří a dvoujaderná buňka které mohou růst a vytvořit další krokodýl, který pak může vytvořit vlastní ascus-počáteční buňku. Tento proces lze poté opakovat několikrát.
Po vytvoření ascus-počáteční buňky se A a A jádro se spojí a vytvoří diploidní jádro (viz obrázek). Toto jádro je jediným diploidním jádrem v celém životním cyklu N. crassa. Diploidní jádro má 14 chromozomů vytvořených ze dvou kondenzovaných haploidních jader, které měly každý po 7 chromozomech. Za vznikem diploidního jádra okamžitě následuje redukční dělení buněk. Dvě postupná rozdělení meiózy vedou ke čtyřem haploidním jádrům, dvěma z A páření a dva z A typ páření. Jedno další mitotické dělení vede ke čtyřem A a čtyři A jádro v každém asku. Meióza je nezbytnou součástí životního cyklu všech pohlavně se množících organismů a ve svých hlavních rysech meióza v N. crassa se zdá být typický pro meiózu obecně.
Vzhledem k tomu, že dochází k výše uvedeným událostem, myceliální obal, který obalil ascogonium, se vyvíjí jako stěna perithecia, impregnuje melaninem a zčernává. Zralý perithecium má baňkovitou strukturu.
Zralý perithecium může obsahovat až 300 asci, každý odvozený od identických fúzních diploidních jader. Obvykle v přírodě, když perithecie dozrávají, jsou askospory vymrštěny poměrně prudce do vzduchu. Tyto askospory jsou odolné vůči teplu a v laboratoři vyžadují zahřívání na 60 ° C po dobu 30 minut, aby došlo ke klíčení. U normálních kmenů trvá celý sexuální cyklus 10 až 15 dní. Ve zralém asku obsahujícím osm askospor jsou páry sousedních spór identické v genetické konstituci, protože poslední dělení je mitotické a protože askospory jsou obsaženy v ascus vaku, který je drží v určitém pořadí určeném směrem nukleárních segregací během redukční dělení buněk. Vzhledem k tomu, že čtyři primární produkty jsou také uspořádány v pořadí, lze rozlišit první segregační vzor genetických markerů od druhého segregačního vzoru.
Genetická analýza jemné struktury
Kvůli výše uvedeným funkcím N. crassa bylo shledáno jako velmi užitečné pro studium genetických událostí vyskytujících se v jednotlivých meiózách. Zralé asci z perithecia mohou být odděleny na mikroskopickém sklíčku a spory experimentálně manipulovány. Tyto studie obvykle zahrnovaly samostatnou kulturu jednotlivých askospor vyplývající z jedné meiotické události a určující genotyp každé spory. Studie tohoto typu prováděné v několika různých laboratořích prokázaly fenomén „genové přeměny“ (viz např. Odkazy)[11][12][13]).
Jako příklad fenoménu genové přeměny zvažte genetické křížení dvou N. crassa mutantní kmeny defektní v gen pánev-2. Tento gen je nezbytný pro syntézu kyseliny pantothenové (vitamin B5) a mutanty defektní v tomto genu lze experimentálně identifikovat podle jejich požadavku na kyselinu pantothenovou v jejich růstovém médiu. Dva pánev-2 mutace B5 a B3 jsou umístěny na různých místech v pánev-2 gen, takže křížením B5 'B3 se získají rekombinanty divokého typu s nízkou frekvencí.[12] Analýza 939 asci, ve které lze určit genotypy všech meiotických produktů (askospor), našla 11 asci s výjimečným segregačním vzorem. Tito zahrnovali šest asci, ve kterých byl jeden meiotický produkt divokého typu, ale neočekávaný reciproční produkt s dvojitou mutací (B5B3). Kromě toho ve třech asci byl poměr meiotických produktů 1B5: 3B3, spíše než v očekávaném poměru 2: 2. Tato studie, stejně jako řada dalších studií v N. crassa a další houby (přezkoumáno Whitehouse[14]), vedlo k rozsáhlé charakterizaci genové konverze. Z této práce vyšlo najevo, že k událostem genové přeměny dochází, když k události molekulární rekombinace dojde v blízkosti studovaných genetických markerů (např. pánev-2 mutace ve výše uvedeném příkladu). Studie genové konverze tedy umožnily nahlédnout do podrobností molekulárního mechanismu rekombinace. V průběhu desetiletí od původních pozorování Mary Mitchellové v roce 1955[11] byla navržena sekvence molekulárních modelů rekombinace na základě nově vznikajících genetických údajů ze studií genové přeměny a studií reakčních schopností DNA. Aktuální pochopení molekulárního mechanismu rekombinace je popsáno v článcích Wikipedie Genová konverze a Genetická rekombinace. Porozumění rekombinaci je relevantní pro několik základních biologických problémů, jako je role rekombinace a rekombinační opravy u rakoviny (viz BRCA1 ) a adaptivní funkce meiózy (viz Redukční dělení buněk ).
Adaptivní funkce typu páření
To páření N. crassa může nastat pouze mezi kmeny různého typu páření naznačuje, že určitý stupeň křížení je upřednostňován přirozeným výběrem. U haploidních mnohobuněčných hub, jako je např N. crassa„Meióza vyskytující se v krátkém diploidním stadiu je jedním z jejich nejsložitějších procesů. Haploidní mnohobuněčný vegetativní stupeň, i když je fyzicky mnohem větší než diploidní stupeň, má charakteristicky jednoduchou modulární konstrukci s malou diferenciací. v N. crassa, u přirozených populací jsou recesivní mutace ovlivňující diploidní fázi životního cyklu poměrně časté.[15] Tyto mutace, jsou-li v diploidním stadiu homozygotní, často způsobují, že výtrusy mají vady zrání nebo produkují neplodná plodnice s několika askosporami (sexuálními výtrusy). Většina těchto homozygotních mutací způsobuje abnormální meiózu (např. Narušení párování chromozomů nebo narušení pachytenu nebo diplotenu).[16] Počet genů ovlivňujících diploidní fázi se odhadoval na nejméně 435[15] (přibližně 4% z celkového počtu 9 730 genů). Křížení, podporované nutností spojení opačných typů páření, tedy pravděpodobně poskytuje výhodu maskování recesivních mutací, které by jinak byly škodlivé pro tvorbu sexuálních spor (viz Doplnění (genetika) ).
Aktuální výzkum
Neurospora crassa je nejen modelovým organismem pro studium fenotypových typů ve vyřazovacích variantách, ale zvláště užitečným organismem široce používaným v výpočetní biologie a cirkadiánní hodiny. Má přirozený reprodukční cyklus 22 hodin a je ovlivňován vnějšími faktory, jako je světlo a teplota. Vyřazte varianty divokého typu N. crassa jsou široce studovány k určení vlivu konkrétních genů (vidět Frekvence (gen) ).
Viz také
- Charles Yanofsky - americký genetik
- David Perkins - americký genetik
- Edward Tatum - americký genetik
- Načechraný transkripční faktor - Gen pro Neurospora crassa potřebný pro bezpohlavní sporulaci
- George Beadle - americký genetik
- Norman Horowitz - americký genetik
- Hypotéza jednoho genu - jednoho enzymu - Myšlenka, že geny působí produkcí enzymů, přičemž každý gen je zodpovědný za produkci jednoho enzymu
- Robert Metzenberg - americký genetik
Poznámky a odkazy
- ^ Davis, Perkins (2002). „Neurospora: model modelových mikrobů“. Genetika hodnocení přírody. 3 (5): 397–403. doi:10.1038 / nrg797. PMID 11988765.
- ^ Iniciativa Trans-NIH Neurospora
- ^ Horowitz NH, Berg P, Singer M a kol. (Leden 2004). „Sté výročí: George W. Beadle, 1903-1989“. Genetika. 166 (1): 1–10. doi:10.1534 / genetika.166.1.1. PMC 1470705. PMID 15020400.
- ^ Galagan J .; Calvo S .; Borkovich K .; Selker E .; Přečtěte si N. D .; et al. (2003). „Sekvence genomu vláknité houby Neurospora crassa". Příroda. 422 (6934): 859–868. Bibcode:2003 Natur.422..859G. doi:10.1038 / nature01554. PMID 12712197.
- ^ Colot H.V .; Park G .; Turner G.E .; Ringleberg C .; Crew C.M .; Litvínova L .; Weiss R.L .; Borkovitch K.A .; Dunlap J.C .; et al. (2006). „Postup vyřazení genu s vysokou propustností pro Neurospora odhaluje funkce pro více transkripčních faktorů ". Sborník Národní akademie věd, USA. 103 (27): 10352–10357. Bibcode:2006PNAS..10310352C. doi:10.1073 / pnas.0601456103. PMC 1482798. PMID 16801547.
- ^ Perkins D. D .; Turner B. C. (1988). "Neurospora z přirozených populací: Směrem k populační biologii haploidního eukaryota “. Experimentální mykologie. 12 (2): 91–131. doi:10.1016/0147-5975(88)90001-1.
- ^ Westergaard M, Mitchell HK (1947). „Neurospora. V.„ Syntetické médium podporující sexuální reprodukci “. Jsem J Bot. 34 (10): 573–577. doi:10.2307/2437339. JSTOR 2437339.
- ^ Nelson MA, Metzenberg RL (září 1992). "Geny sexuálního vývoje Neurospora crassa". Genetika. 132 (1): 149–62. PMC 1205113. PMID 1356883.
- ^ Fincham J RS, Day PR (1963). Plísňová genetika. Blackwell Scientific Publications, Oxford, Velká Británie. ASIN: B000W851KO
- ^ Wagner RP, Mitchell HK. (1964). Genetika a metabolismus. John Wiley and Sons, Inc., New York ASIN: B00BXTC5BO
- ^ A b Mitchell MB (duben 1955). „ABERRANTNÍ RECOMBINACE PYRIDOXINOVÝCH MUTANTŮ Neurospora“. Proc. Natl. Acad. Sci. USA. 41 (4): 215–20. Bibcode:1955PNAS ... 41..215M. doi:10.1073 / pnas.41.4.215. PMC 528059. PMID 16589648.
- ^ A b Případ ME, Giles NH (květen 1958). „DŮKAZY Z ANALÝZY TETRADU PRO OBĚ NORMÁLNÍ A ABERRANTNÍ REKOMBINACI MEZI ALELICKÝMI MUTANTY V Neurospora Crassa“. Proc. Natl. Acad. Sci. USA. 44 (5): 378–90. Bibcode:1958PNAS ... 44..378C. doi:10.1073 / pnas.44.5.378. PMC 335434. PMID 16590210.
- ^ Stadler DR (červenec 1959). „Génová konverze cysteinových mutantů v Neurospora“. Genetika. 44 (4): 647–56. PMC 1209971. PMID 17247847.
- ^ Whitehouse, HLK. (1982). Genetická rekombinace. Wiley, New York ISBN 978-0471102052
- ^ A b Leslie JF, Raju NB (prosinec 1985). „Recesivní mutace z přirozených populací Neurospora crassa, které jsou exprimovány v sexuální diplofáze“. Genetika. 111 (4): 759–77. PMC 1202670. PMID 2933298.
- ^ Raju NB, Leslie JF (říjen 1992). „Cytologie recesivních mutantů v sexuální fázi z divokých kmenů Neurospora crassa“. Genom. 35 (5): 815–26. doi:10,1139 / g92-124. PMID 1427061.
Reference
- Perkins, D; Davis, R (prosinec 2000), „Důkazy o bezpečnosti druhů Neurospora pro akademická a komerční použití“ (PDF), Aplikovaná a environmentální mikrobiologie, 66 (12), s. 5107–5109, doi:10.1128 / aem.66.12.5107-5109.2000, PMC 92429, PMID 11097875.
- Osherov, N; Květen, GS (30. května 2001), „Molekulární mechanismy konidiálního klíčení“, FEMS Microbiol Lett, 199 (2), s. 153–60, doi:10.1111 / j.1574-6968.2001.tb10667.x, PMID 11377860
- Froehlich, AC; No, B; Vierstra, RD, Loros J & Dunlap JC (prosinec 2005), „Genetická a molekulární analýza fytochromů z vláknité houby Neurospora crassa“, Eukaryotova buňka, 4 (12), s. 2140–52, doi:10.1128 / ec.4.12.2140-2152.2005, PMC 1317490, PMID 16339731
- Horowitz, NH (duben 1991), „Před padesáti lety: revoluce Neurospora“, Genetika, 127 (4), s. 631–5, PMC 1204391, PMID 1827628
- Horowitz, NH; Berg, P; Singer, M, Lederberg J, Susman M, Doebley J & Crow JF. (Leden 2004), „Sté výročí: George W. Beadle, 1903-1989“, Genetika, 166 (1), s. 1–10, doi:10.1534 / genetika.166.1.1, PMC 1470705, PMID 15020400
- Kaldi, K; Gonzalez, BH; Brunner, M (23. prosince 2005), „Transkripční regulace genu cirkadiánních hodin wc-1 Neurospora ovlivňuje fázi cirkadiánního výdeje“, EMBO Rep, 7 (2), s. 199–204, doi:10.1038 / sj.embor.7400595, PMC 1369249, PMID 16374510
- Pittalwala, Iqbal (29. dubna 2003), „Vědci z UC Riverside přispívají ke studiu, které odhaluje sekvenci genomu formy na chléb“, Newsroom (University of California, Riverside).
- Ruoff, P; Loros, JJ; Dunlap, JC (6. prosince 2005), „Vztah mezi stabilitou proteinu FRQ a teplotní kompenzací v cirkadiánních hodinách Neurospora“, Proc Natl Acad Sci USA, 102 (49), s. 17681–6, Bibcode:2005PNAS..10217681R, doi:10.1073 / pnas.0505137102, PMC 1308891, PMID 16314576
externí odkazy
- Neurospora crassa genom [1]
- „Domovská stránka Neurospora“. Fungal Genetics Stock Center (FGSC). Citováno 27. prosince 2005.
- „The Neurospora Compendium“. Fungal Genetics Stock Center (FGSC). Citováno 27. prosince 2005.
- „Iniciativa Neurospora-Fungal Genome“. Projekt genomu Neurospora. Archivovány od originál 4. března 2016. Citováno 12. června 2015.
- „Iniciativa Trans-NIH Neurospora“. National Institutes of Health (NIH - Spojené státy). Citováno 27. prosince 2005.
- [2] Montenegro-Montero A. (2010) "The Všemohoucí houby: Revoluční Neurospora crassa". Historický pohled na mnoho příspěvků tohoto organismu k molekulární biologii.