Gibberellin - Gibberellin
tento článek může obsahovat nadměrné množství složitých detailů, které mohou zajímat pouze konkrétní publikum. (Září 2019) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
Gibberelliny (GA) jsou rostlinné hormony které regulují různé vývojové procesy, počítaje v to zastavit prodloužení, klíčení, klidu, kvetoucí, květ vývoj a list a ovoce stárnutí.[1] GA jsou jednou z nejdéle známých tříd rostlinných hormonů. Předpokládá se, že šlechtění (i když v bezvědomí) kmenů plodin, které postrádaly syntézu GA, byl jedním z klíčových faktorů „zelená revoluce „v 60. letech[2] revoluce, o níž se připisuje, že zachránila více než miliardu životů po celém světě.[3]
Dějiny
První narážkou na porozumění GA byl vývoj z rostlinná patologie oboru, se studiemi na Bakanae nebo nemoc "pošetilých sazenic" v rýže. Pošetilá choroba sazenic způsobuje silné prodloužení stonků a listů rýže a nakonec způsobí jejich převrácení.[4] V roce 1926 japonský vědec Eiichi Kurosawa zjistil, že pošetilá choroba sazenic byla způsobena houba Gibberella fujikuroi.[4] Pozdější práce na Tokijské univerzitě ukázaly, že látka produkovaná touto houbou vyvolala příznaky pošetilé nemoci sazenic a tuto látku pojmenovali „giberelin“.[1][4]
Následovala zvýšená komunikace mezi Japonskem a Západem druhá světová válka zvýšil zájem o gibberellin v EU Spojené království (UK) a Spojené státy (NÁS).[1] Pracovníci v Imperial Chemical Industries ve Velké Británii[5] a ministerstvo zemědělství v USA samostatně izolované kyselina giberelová[4] (s tím, že Američané tuto chemikálii původně označovali jako „gibberellin-X“, než přijali britský název - tato chemikálie je v Japonsku známá jako giberelin A3 nebo GA3)[1]
Znalosti o giberelinách se rozšířily po celém světě, protože se stal zřetelnějším potenciál pro jejich použití v různých komerčně významných rostlinách. Například výzkum, který začal na University of California, Davis v polovině 60. let 20. století vedlo ke komerčnímu využití Stolové hrozny Thompson bez pecek po celé Kalifornii do roku 1962.[6][je zapotřebí objasnění ] Známým inhibitorem biosyntézy giberelinu je paclobutrazol (PBZ), což zase inhibuje růst a vyvolává časnou sadu plodů i sadu semen.
Během prudkého stoupání světové populace v šedesátých letech se obával chronického nedostatku potravin. To bylo odvráceno vývojem vysoce výnosné odrůdy rýže. Tato odrůda polotrpasličí rýže se nazývá IR8, a má malou výšku kvůli mutaci genu sd1.[7] Sd1 kóduje GA20ox, takže se očekává, že mutantní sd1 bude vykazovat malou výšku, která odpovídá deficitu GA.[2]
Chemie
Všechny známé gibereliny jsou diterpenoid kyseliny, které jsou syntetizovány terpenoidní cestou v plastidy a poté upraven v endoplazmatické retikulum a cytosol dokud nedosáhnou své biologicky aktivní formy.[8] Všechny gibereliny jsou odvozeny prostřednictvím ent-gibberellane kostra, ale jsou syntetizovány prostřednictvím ent-kaurene. Gibereliny jsou pojmenovány GA1 až GAn v pořadí podle objevu. Kyselina giberelová, který byl prvním strukturálně charakterizovaným giberelinem, je GA3.
V roce 2003 bylo identifikováno 126 GA z rostlin, hub a bakterií.[1]
Gibberelliny jsou tetracyklické diterpenové kyseliny. Existují dvě třídy založené na přítomnosti buď 19 nebo 20 uhlíků. 19-uhlíkaté gibereliny, jako je kyselina giberelinová, ztratily uhlík 20 a na místě mají pětičlenný lakton most, který spojuje uhlíky 4 a 10. 19-uhlíkové formy jsou obecně biologicky aktivní formy giberelinů. Hydroxylace má také velký účinek na biologickou aktivitu giberelinu. Obecně jsou biologicky nejaktivnějšími sloučeninami dihydroxylované gibereliny, které mají hydroxylové skupiny jak na uhlíku 3, tak na uhlíku 13. Kyselina giberelová je dihydroxylovaný giberelin.[9]
Bioaktivní GA
Bioaktivní GA jsou GA1, GA3, GA4 a GA7.[10] Mezi těmito GA jsou tři společné strukturní rysy: hydroxylová skupina na C-3β, karboxylová skupina na C-6 a lakton mezi C-4 a C-10.[10] 3β-hydroxylovou skupinu lze vyměnit za jiné funkční skupiny v polohách C-2 a / nebo C-3.[10] GA5 a GA6 jsou příklady bioaktivních GA, které nemají hydroxylovou skupinu na C-3β.[10] Přítomnost GA1 u různých druhů rostlin naznačuje, že se jedná o běžný bioaktivní GA.[11]

Gibberellin A1 (GA1) 
Kyselina giberelová (GA3) 
ent-Gibberellane 
ent-Kaurene




Biologická funkce

Gibberelliny jsou zapojeny do přirozeného procesu lámání klidu a další aspekty klíčení. Před tím, než se fotosyntetický aparát dostatečně rozvíjí v počátečních stádiích klíčení, jsou uložené zásoby energie škrob vyživujte sazenici. Obvykle v klíčení, rozkladu škrobu na glukóza v endosperm začíná krátce poté, co je semeno vystaveno vodě.[12] Předpokládá se, že gibberelliny v semenném embryu signalizují škrob hydrolýza indukcí syntézy enzymu α-amyláza v aleuronových buňkách. V modelu pro produkci α-amylázy vyvolanou gibberellinem se ukazuje, že gibereliny (označované GA) produkované v scutellum difundují do aleuronových buněk, kde stimulují sekreci α-amylázy.[8] α-amyláza poté hydrolyzuje škrob, který je hojný v mnoha semenech, na glukózu, kterou lze použít v buněčném dýchání k výrobě energie pro embryo semen. Studie tohoto procesu ukázaly, že gibereliny způsobují vyšší hladiny transkripce genu kódujícího enzym a-amylázu, stimulovat syntézu a-amylázy.[9]
Gibberelliny se produkují ve větší hmotnosti, když je rostlina vystavena nízkým teplotám. Stimulují prodloužení buněk, lámání a pučení, plody bez semen a klíčení semen. Gibberelliny způsobují klíčení semen porušením vegetačního klidu semen a působením chemického posla. Jeho hormon se váže na receptor a vápník aktivuje protein klimodulin a komplex se váže na DNA a produkuje enzym stimulující růst embrya.
Metabolismus
Biosyntéza
GA jsou obvykle syntetizovány z methylerythritol fosfát (MEP) cesta ve vyšších rostlinách.[13] V této cestě je bioaktivní GA produkován z trans-geranylgeranyldifosfátu (GGDP).[13] V dráze MEP se k získání GA z GGDP používají tři třídy enzymů: terpenové syntézy (TPS), cytochrom P450 monooxygenázy (P450s) a 2-oxoglutarát-dependentní dioxygenázy (2ODDs).[10] Cesta MEP zahrnuje osm kroků:[10]
- GGDP se převádí na ent-kopalyldifosfát (ent-CPD) ent-kopalyldifosfát syntázou
- etn-CDP se převádí na ent-kauren ent-kaurensyntázou
- ent-kauren se převádí na ent-kaurenol ent-kauren oxidázou (KO)
- ent-kaurenol se převádí na ent-kaurenal pomocí KO
- ent-kaurenal se převádí na kyselinu ent-kaurenovou pomocí KO
- kyselina ent-kaurenová se převádí na ent-7a-hydroxykaurenovou kyselinu pomocí oxidázy ent-kaurenové kyseliny (KAO)
- Kyselina ent-7a-hydroxykaurenová se převádí na GA12-aldehyd pomocí KAO
- GA12-aldehyd se převádí na GA12 pomocí KAO. GA12 je zpracován na bioaktivní GA4 oxidací na C-20 a C-3, čehož je dosaženo 2 rozpustnými ODD: GA 20-oxidáza a GA 3-oxidáza.
Jeden nebo dva geny kódují enzymy odpovědné za první kroky biosyntézy GA v Arabidopsis a rýže.[10] Nulové alely genů kódujících CPS, KS a KO vedou k deficitu GA Arabidopsis trpaslíci.[14] Multigenové rodiny kódují 2ODD, které katalyzují tvorbu GA12 na bioaktivní GA4.[10]
AtGA3ox1 a AtGA3ox2, dva ze čtyř genů, které kódují GA3ox Arabidopsis, ovlivňují vegetativní vývoj.[15] Podněty prostředí regulují aktivitu AtGA3ox1 a AtGA3ox2 během klíčení semen.[16][17] v Arabidopsis„Nadměrná exprese GA20ox vede ke zvýšení koncentrace GA.[18][19]
Místa biosyntézy
Většina bioaktivních GA se nachází v aktivně rostoucích orgánech na rostlinách.[13] Oba geny GA20ox a GA3ox (geny kódující GA 20-oxidázu a GA 3-oxidázu) a gen SLENDER1 (GA signální transdukce Gen) se nacházejí v rostoucích orgánech na rýži, což naznačuje, že v jejich místě působení v rostoucích orgánech v rostlinách dochází k bioaktivní syntéze GA.[20] Během vývoje květu je tapetum prašníků považováno za primární místo biosyntézy GA.[20][21]
Rozdíly mezi biosyntézou hub a nižších rostlin
Arabidopsis, rostlina, a Gibberella fujikuroi Houba má různé dráhy GA a enzymy.[10] P450 v houbách plní funkce analogické funkcím KAO v rostlinách.[22] Funkci CPS a KS v rostlinách vykonává jediný enzym, CPS / KS, v houbách.[23][24][25] U hub se geny pro biosyntézu GA nacházejí na jednom chromozomu, ale v rostlinách se vyskytují náhodně na více chromozomech.[26][27] Rostliny produkují malé množství GA3, proto je GA3 produkován pro průmyslové účely mikroorganismy. Kyselinu giberelovou lze průmyslově vyrábět ponořenou fermentací, ale tento proces představuje nízký výtěžek s vysokými výrobními náklady, a tudíž vyšší prodejní hodnotou, nicméně jiný alternativní postup ke snížení nákladů na výrobu GA3 je fermentace v pevném stavu (SSF), který umožňuje použití zemědělsko-průmyslových zbytků.[28]
Katabolismus
Bylo identifikováno několik mechanismů pro deaktivaci GA. 2β-hydroxylace deaktivuje GA a je katalyzována GA2-oxidázami (GA2oxs).[13] Některé GA2oxy používají jako substráty C19-GA a jiné GA2oxy používají C20-GA.[29][30] Monooxygenáza cytochromu P450, kódovaná prodlouženou nejvyšší internodou (eui), převádí GA na 16α, 17-epoxidy.[31] Mutanti rýžového eui hromadí bioaktivní GA na vysokých úrovních, což naznačuje, že cytochrom P450 mono-oxygenáza je hlavním enzymem odpovědným za deaktivaci GA v rýži.[31] Geny Gamt1 a gamt2 kódují enzymy, které methylují C-6 karboxylovou skupinu GA.[32] U mutantů gamt1 a gamt2 se zvyšuje koncentrace GA vyvíjejících se semen.[32]
Homeostáza
Zpětná vazba a dopředná regulace udržuje hladinu bioaktivních GA v rostlinách.[33][34] Úrovně exprese AtGA20ox1 a AtGA3ox1 se zvyšují v prostředí s nedostatkem GA a snižují se po přidání bioaktivních GA,[16][35][36][37][38] Naopak exprese AtGA2ox1 a AtGA2ox2, GA deaktivačních genů, se zvyšuje přidáním GA.[29]
Nařízení
Regulace jinými hormony
Kyselina auxinindol-3-octová (IAA) reguluje koncentraci GA1 v prodlužujících se internodiích v hrášku.[39] Odstranění IAA odstraněním apikálního pupenu, zdroje auxinu, snižuje koncentraci GA1 a opětovné zavedení IAA tyto účinky zvrací a zvyšuje koncentraci GA1.[39] Tento jev byl také pozorován u rostlin tabáku.[40] Auxin zvyšuje GA 3-oxidaci a snižuje GA 2-oxidaci u ječmene.[41] Auxin také reguluje biosyntézu GA během vývoje ovoce v hrášku.[42] Tyto objevy u různých druhů rostlin naznačují, že regulace auxinu v metabolismu GA může být univerzálním mechanismem.
Ethylen snižuje koncentraci bioaktivních GA.[43]
Regulace faktory prostředí
Nedávné důkazy naznačují, že kolísání koncentrace GA ovlivňuje světlo regulované klíčení semen, fotomorfogeneze v době deetiolace, a fotoperioda regulace prodloužení a kvetení stonku.[10] Analýza Microarray ukázala, že přibližně čtvrtina genů reagujících na chlad je příbuzná s geny regulovanými GA, což naznačuje, že GA ovlivňuje reakci na chladné teploty.[17] Rostliny snižují rychlost růstu, jsou-li vystaveny stresu. U ječmene byl navržen vztah mezi hladinami GA a množstvím prožívaného stresu.[44]
Role ve vývoji semen
Bioaktivní GA a kyselina abscisová úrovně mají inverzní vztah a regulují vývoj semen a klíčení.[45][46] Úrovně FUS3, an Arabidopsis transkripční faktor, jsou upregulovány ABA a downregulovány GA, což naznačuje, že existuje regulační smyčka, která stanoví rovnováhu GA a ABA.[47]
Signalizační mechanismus
Receptor
Na počátku 90. let existovalo několik linií důkazů, které naznačovaly existenci GA receptoru v oves semena, která byla umístěna na plazmatická membrána. Navzdory intenzivnímu výzkumu však dosud nebyl izolován žádný receptor GA vázaný na membránu. To spolu s objevem rozpustného receptoru, GA necitlivého trpaslíka 1 (GID1) vedlo mnoho lidí k pochybnostem, že existuje membránově vázaný receptor.[1]
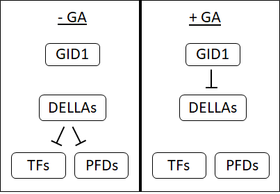
GID1 byl poprvé identifikován v rýže[48] a v Arabidopsis existují tři ortology GID1, AtGID1a, b a c.[1] GID1 mají vysokou afinitu k bioaktivní Plyn.[48] GA se váže na specifickou vázací kapsu na GID1; C3-hydroxyl na GA navazuje kontakt s tyrosinem-31 ve vazebné kapse GID1.[49][50] Vazba GA na GID1 způsobí změny ve struktuře GID1, což způsobí, že „víčko“ na GID1 pokryje kapsu vazby GA. Pohyb tohoto víčka má za následek vystavení povrchu, který umožňuje vazbu GID1 na proteiny DELLA.[49][50]
Proteiny DELLA: Represe represoru
Proteiny DELLA, jako je SLR1 v rýži nebo GAI a RGA v Arabidopsis jsou represory vývoje rostlin. DELLA inhibují klíčení semen, růst semen, kvetení a GA tyto účinky zvrací.[51] Proteiny DELLA se vyznačují přítomností motivu DELLA (aspartát -glutamát -leucin -leucin-alanin nebo D-E-L-L-A jediným písmenem kód aminokyseliny ).[52]
Když se GA váže na receptor GID1, zvyšuje interakci mezi proteiny GID1 a DELLA a vytváří komplex GA-GID1-DELLA. V komplexu GA-GID1-DELLA se předpokládá, že proteiny DELLA procházejí změnami struktury, které umožňují jejich vazbu na F-box proteiny (SLY1 v Arabidopsis nebo GID2 v rýži).[53][52][54] F-box proteiny katalyzovat přidání ubikvitin k jejich cílům.[53] Přidání ubikvitinu k proteinům DELLA podporuje jejich degradaci prostřednictvím 26S-proteosom.[52] Degradace proteinů DELLA uvolňuje buňky z jejich represivních účinků.
Cíle proteinů DELLA
Transkripční faktory
Prvními identifikovanými cíli proteinů DELLA byly FYTOCHROMATICKÉ INTERAKTIVNÍ FAKTORY (PIF). PIF jsou transkripční faktory které negativně regulují světelnou signalizaci a jsou silnými promotory růstu prodloužení. V přítomnosti GA jsou DELLA degradovány a to pak umožňuje PIF podporovat prodloužení.[55] Později bylo zjištěno, že DELLA potlačují velké množství dalších transkripčních faktorů, mezi nimiž jsou pozitivní regulátory auxin, brassinosteriod a ethylen signalizace.[56][57] DELLA mohou potlačovat transkripční faktory buď zastavením jejich vazby na DNA, nebo podporou jejich degradace.[55]
Sestava prefoldinů a mikrotubulů
Kromě potlačování transkripčních faktorů se DELLA váží také na prefoldiny (PFD). PFD jsou molekulární chaperony, což znamená, že pomáhají při skládání dalších proteinů. Funkce PFD v cytosol ale když se DELLA váží na PFD, omezuje je na jádro. Důležitou funkcí PFD je pomoc při skládání β-tubulin. V nepřítomnosti GA (pokud je vysoká hladina proteinů DELLA) je tedy funkce PDF snížena a existuje nižší buněčná zásoba β-tubulinu. Když je přítomen GA, DELLA jsou degradovány, soubory PDF se mohou přesunout na cytosol a pomáhat při skládání β-tubulinu. β-tubulin je životně důležitou složkou cytoskelet (ve formě mikrotubuly ). Jako takový umožňuje GA reorganizaci cytoskeletu a prodloužení buněk.[58]
Mikrotubuly jsou také nutné pro obchodování s membránové vezikuly. K správnému umístění několika je zapotřebí obchodování s membránovými váčky transportéry hormonů. Jedním z nejlépe charakterizovaných transportérů hormonů jsou PIN proteiny, které jsou odpovědné za pohyb hormonu auxinu mezi buňkami. V nepřítomnosti GA proteiny DELLA snižují hladiny mikrotubulů a tím inhibují transport membránových vezikul. To snižuje hladinu PIN proteinů na buněčná membrána a úroveň auxinu v buňce. GA tento proces obrací a umožňuje přenos PIN proteinu do buněčné membrány za účelem zvýšení hladiny auxinu v buňce.[59]
Reference
- ^ A b C d E F G Hedden P, Sponsel V (2015). „Století výzkumu gibberellinu“. Časopis regulace růstu rostlin. 34 (4): 740–60. doi:10.1007 / s00344-015-9546-1. PMC 4622167. PMID 26523085.
- ^ A b Spielmeyer W, Ellis MH, Chandler PM (červen 2002). „Semidwarf (sd-1),„ zelená revoluce “rýže, obsahuje vadný gen gibberellin 20-oxidázy“. Sborník Národní akademie věd Spojených států amerických. 99 (13): 9043–8. Bibcode:2002PNAS ... 99.9043S. doi:10.1073 / pnas.132266399. PMC 124420. PMID 12077303.
- ^ „Norman Borlaug: Miliarda životů zachráněna“. www.agbioworld.org. Citováno 2018-05-11.
- ^ A b C d B B Stowe; Yamaki a T. (1957). „Historie a fyziologické působení giberelinů“. Roční přehled fyziologie rostlin. 8 (1): 181–216. doi:10.1146 / annurev.pp.08.060157.001145.
- ^ Mees, G.C .; Elson, G.W. (1978). „Kapitola 7: Gibereliny“. V Peacock, F.C. (vyd.). Jealott's Hill: Padesát let zemědělského výzkumu v letech 1928-1978. Imperial Chemical Industries Ltd. pp.55 –60. ISBN 0901747017.
- ^ Gibberellin a bezsemenné hrozny Archivováno 2006-12-06 na Wayback Machine od a University of California, Davis webová stránka
- ^ Sasaki A, Ashikari M, Ueguchi-Tanaka M, Itoh H, Nishimura A, Swapan D, Ishiyama K, Saito T, Kobayashi M, Khush GS, Kitano H, Matsuoka M (duben 2002). „Zelená revoluce: mutantní gen pro syntézu giberelinu v rýži“. Příroda. 416 (6882): 701–2. Bibcode:2002 Natur.416..701S. doi:10.1038 / 416701a. PMID 11961544. S2CID 4414560.
- ^ A b Campbell N, Reec JB (2002). Biologie (6. vydání). San Francisco: Benjamin Cummings.
- ^ A b „Gibberellins“. AccessScience. doi:10.1036/1097-8542.289000.
- ^ A b C d E F G h i j Yamaguchi S (2008). "Metabolismus gibberellinu a jeho regulace". Roční přehled biologie rostlin. 59: 225–51. doi:10.1146 / annurev.arplant. 59.032607.092804. PMID 18173378.
- ^ MacMillan J (prosinec 2001). "Výskyt giberelinů u cévnatých rostlin, hub a bakterií". Časopis regulace růstu rostlin. 20 (4): 387–442. doi:10,1007 / s003440010038. PMID 11986764. S2CID 44504525.
- ^ Davies PJ. "Růst rostlin". AccessScience. doi:10.1036/1097-8542.523000.
- ^ A b C d Hedden P, Thomas SG (květen 2012). „Biosyntéza gibberellinu a její regulace“. The Biochemical Journal. 444 (1): 11–25. doi:10.1042 / BJ20120245. PMID 22533671. S2CID 25627726.
- ^ Koornneef M, van der Veen JH (listopad 1980). "Indukce a analýza mutantů citlivých na gibberellin v Arabidopsis thaliana (L.) heynh". ŠTÍTEK. Teoretická a aplikovaná genetika. Theoretische und Angewandte Genetik. 58 (6): 257–63. doi:10.1007 / BF00265176. PMID 24301503. S2CID 22824299.
- ^ Mitchum MG, Yamaguchi S, Hanada A, Kuwahara A, Yoshioka Y, Kato T, Tabata S, Kamiya Y, Sun TP (březen 2006). „Výrazné a překrývající se role dvou gibberellin 3-oxidáz ve vývoji Arabidopsis“. The Plant Journal. 45 (5): 804–18. doi:10.1111 / j.1365-313X.2005.02642.x. PMID 16460513.
- ^ A b Yamaguchi S, Smith MW, Brown RG, Kamiya Y, Sun T (prosinec 1998). „Regulace fytochromu a diferenciální exprese genů gibberellinu 3-beta-hydroxylázy v klíčících semenech Arabidopsis“. Rostlinná buňka. 10 (12): 2115–26. doi:10.1105 / tpc.10.12.2115. PMC 143973. PMID 9836749.
- ^ A b Yamauchi Y, Ogawa M, Kuwahara A, Hanada A, Kamiya Y, Yamaguchi S (únor 2004). „Aktivace biosyntézy giberelinu a dráhy odezvy při nízké teplotě během vstřebávání semen Arabidopsis thaliana“. Rostlinná buňka. 16 (2): 367–78. doi:10.1105 / tpc.018143. PMC 341910. PMID 14729916.
- ^ Coles JP, Phillips AL, Croker SJ, García-Lepe R, Lewis MJ, Hedden P (březen 1999). „Modifikace produkce gibberellinu a vývoj rostlin u Arabidopsis pomocí smyslové a antisense exprese genů gibberellin 20-oxidázy“. The Plant Journal. 17 (5): 547–56. doi:10.1046 / j.1365-313X.1999.00410.x. PMID 10205907.
- ^ Huang S, Raman AS, Ream JE, Fujiwara H, Cerny RE, Brown SM (listopad 1998). „Nadměrná exprese 20-oxidázy poskytuje u Arabidopsis fenotyp nadprodukce gibberellinu“. Fyziologie rostlin. 118 (3): 773–81. doi:10,1104 / pp.118.3.773. PMC 34787. PMID 9808721.
- ^ A b Kaneko M, Itoh H, Inukai Y, Sakamoto T, Ueguchi-Tanaka M, Ashikari M, Matsuoka M (červenec 2003). „Kde se vyskytuje biosyntéza giberelinu a signalizace giberelinu v rostlinách rýže?“. The Plant Journal. 35 (1): 104–15. doi:10.1046 / j.1365-313X.2003.01780.x. PMID 12834406.
- ^ Itoh H, Tanaka-Ueguchi M, Kawaide H, Chen X, Kamiya Y, Matsuoka M (říjen 1999). „Gen kódující tabákový gibberellin 3beta-hydroxylázu je exprimován v místě působení GA během prodloužení stonku a vývoje květních orgánů“. The Plant Journal. 20 (1): 15–24. doi:10.1046 / j.1365-313X.1999.00568.x. PMID 10571861.
- ^ Rojas MC, Hedden P, Gaskin P, Tudzynski B (květen 2001). „Gen P450-1 gibberella fujikuroi kóduje multifunkční enzym v biosyntéze giberelinu“. Sborník Národní akademie věd Spojených států amerických. 98 (10): 5838–43. Bibcode:2001PNAS ... 98.5838R. doi:10.1073 / pnas.091096298. PMC 33300. PMID 11320210.
- ^ Kawaide H, Imai R, Sassa T, Kamiya Y (srpen 1997). „Ent-kaurensyntáza z houby Phaeosphaeria sp. L487. Izolace, charakterizace a bakteriální exprese bifunkční diterpencyklázy v biosyntéze houbového giberelinu“ cDNA. The Journal of Biological Chemistry. 272 (35): 21706–12. doi:10.1074 / jbc.272.35.21706. PMID 9268298.
- ^ Toyomasu T, Kawaide H, Ishizaki A, Shinoda S, Otsuka M, Mitsuhashi W, Sassa T (březen 2000). „Klonování plné délky cDNA kódující ent-kaurensyntázu z Gibberella fujikuroi: funkční analýza bifunkční diterpenové cyklázy“. Bioscience, biotechnologie a biochemie. 64 (3): 660–4. doi:10,1271 / bbb.64,660. PMID 10803977.
- ^ Tudzynski B, Kawaide H, Kamiya Y (září 1998). „Biosyntéza gibberellinu v gibberella fujikuroi: klonování a charakterizace genu kopalyldifosfát syntázy“. Současná genetika. 34 (3): 234–40. doi:10,1007 / s002940050392. PMID 9745028. S2CID 3021994.
- ^ Hedden P, Phillips AL, Rojas MC, Carrera E, Tudzynski B (prosinec 2001). „Biosyntéza gibberellinu u rostlin a hub: případ konvergentní evoluce?“. Časopis regulace růstu rostlin. 20 (4): 319–331. doi:10,1007 / s003440010037. PMID 11986758. S2CID 25623658.
- ^ Kawaide H (březen 2006). "Biochemické a molekulární analýzy biosyntézy giberelinu v houbách". Bioscience, biotechnologie a biochemie. 70 (3): 583–90. doi:10,1271 / bbb.70,583. PMID 16556972. S2CID 20952424.
- ^ Lopes AL, Silva DN, Rodrigues C, Costa JL, Machado MP, Penha RO, Biasi LA, Ricardo C (2013). „Extrakt fermentovaný kyselinou giberelovou získaný fermentací v pevném stavu s použitím buničiny citronové od Fusarium moniliforme: Vliv na Lavandula angustifolia Mill. Kultivovaný in vitro.“ Pak J Bot. 45: 2057–2064.
- ^ A b Thomas SG, Phillips AL, Hedden P (duben 1999). „Molekulární klonování a funkční exprese gibberellin-2-oxidázy, multifunkční enzymy podílející se na deaktivaci gibberellinu“. Sborník Národní akademie věd Spojených států amerických. 96 (8): 4698–703. Bibcode:1999PNAS ... 96.4698T. doi:10.1073 / pnas.96.8.4698. PMC 16395. PMID 10200325.
- ^ Schomburg FM, Bizzell CM, Lee DJ, Zeevaart JA, Amasino RM (leden 2003). „Nadměrná exprese nové třídy 2-oxidáz gibberellinu snižuje hladinu giberelinu a vytváří zakrslé rostliny“. Rostlinná buňka. 15 (1): 151–63. doi:10,1105 / tpc.005975. PMC 143488. PMID 12509528.
- ^ A b Zhu Y, Nomura T, Xu Y, Zhang Y, Peng Y, Mao B, Hanada A, Zhou H, Wang R, Li P, Zhu X, Mander LN, Kamiya Y, Yamaguchi S, He Z (únor 2006). „DLOUHODOBÝ NEJVYŠŠÍ INTERNODE kóduje monooxygenázu cytochromu P450, která epoxiduje gibereliny v nové deaktivační reakci v rýži“. Rostlinná buňka. 18 (2): 442–56. doi:10.1105 / tpc.105.038455. PMC 1356550. PMID 16399803.
- ^ A b Varbanova M, Yamaguchi S, Yang Y, McKelvey K, Hanada A, Borochov R, Yu F, Jikumaru Y, Ross J, Cortes D, Ma CJ, Noel JP, Mander L, Shulaev V, Kamiya Y, Rodermel S, Weiss D , Pichersky E (leden 2007). „Methylace giberelinů Arabidopsis GAMT1 a GAMT2“. Rostlinná buňka. 19 (1): 32–45. doi:10.1105 / tpc.106.044602. PMC 1820973. PMID 17220201.
- ^ Hedden P, Phillips AL (prosinec 2000). „Metabolismus gibberellinu: nové poznatky odhalené geny“. Trendy ve vědě o rostlinách. 5 (12): 523–30. doi:10.1016 / S1360-1385 (00) 01790-8. PMID 11120474.
- ^ Olszewski N, Sun TP, Gubler F (2002). „Signalizace gibberellinu: biosyntéza, katabolismus a cesty odezvy“. Rostlinná buňka. 14 Suppl (Suppl): S61–80. doi:10.1105 / tpc.010476. PMC 151248. PMID 12045270.
- ^ Chiang HH, Hwang I, Goodman HM (únor 1995). "Izolace lokusu Arabidopsis GA4". Rostlinná buňka. 7 (2): 195–201. doi:10,1105 / tpc.7.2.195. PMC 160775. PMID 7756830.
- ^ Matsushita A, Furumoto T, Ishida S, Takahashi Y (březen 2007). „AGF1, protein AT-hook, je nezbytný pro negativní zpětnou vazbu AtGA3ox1 kódujícího GA 3-oxidázu“. Fyziologie rostlin. 143 (3): 1152–62. doi:10.1104 / str. 106.093542. PMC 1820926. PMID 17277098.
- ^ Phillips AL, Ward DA, Uknes S, Appleford NE, Lange T, Huttly AK, Gaskin P, Graebe JE, Hedden P (červenec 1995). „Izolace a exprese tří cDNA klonů gibberellin-20-oxidázy z Arabidopsis“. Fyziologie rostlin. 108 (3): 1049–57. doi:10.1104 / str. 108.3.1049. PMC 157456. PMID 7630935.
- ^ Xu YL, Li L, Gage DA, Zeevaart JA (květen 1999). „Regulace zpětné vazby exprese GA5 a metabolické inženýrství hladin giberelinu u Arabidopsis“. Rostlinná buňka. 11 (5): 927–36. doi:10,1105 / tpc.11.5.927. PMC 144230. PMID 10330476.
- ^ A b Ross JJ, O'Neill DP, Smith JJ, Kerckhoffs LH, Elliott RC (březen 2000). "Důkaz, že auxin podporuje biosyntézu gibberellinu A1 v hrachu". The Plant Journal. 21 (6): 547–52. doi:10.1046 / j.1365-313x.2000.00702.x. PMID 10758505.
- ^ Wolbang CM, Ross JJ (listopad 2001). „Auxin podporuje biosyntézu giberelinu v rostlinách tabáku s dekapitací“. Planta. 214 (1): 153–7. doi:10,1007 / s004250100663. PMID 11762165. S2CID 31185063.
- ^ Wolbang CM, Chandler PM, Smith JJ, Ross JJ (únor 2004). „Auxin z vyvíjejícího se květenství je nutný pro biosyntézu aktivních giberelinů ve stoncích ječmene“. Fyziologie rostlin. 134 (2): 769–76. doi:10.1104 / pp.103.030460. PMC 344552. PMID 14730077.
- ^ Ngo P, Ozga JA, Reinecke DM (červenec 2002). "Specifičnost auxinové regulace exprese genu gibberellin 20-oxidázy v perikarpu hrachu". Molekulární biologie rostlin. 49 (5): 439–48. doi:10.1023 / A: 1015522404586. PMID 12090620. S2CID 22530544.
- ^ Achard P, Baghour M, Chapple A, Hedden P, Van Der Straeten D, Genschik P, Moritz T, Harberd NP (duben 2007). „Rostlinný stresový hormon ethylen řídí květinový přechod regulací květinových meristemových identit závislých na DELLA“. Sborník Národní akademie věd Spojených států amerických. 104 (15): 6484–9. Bibcode:2007PNAS..104,6484A. doi:10.1073 / pnas.0610717104. PMC 1851083. PMID 17389366.
- ^ Vettakkorumakankav NN, Falk D, Saxena P, Fletcher RA (1999). „Zásadní role giberelinů při ochraně rostlin před stresem“. Fyziologie rostlin a buněk. 40 (5): 542–548. doi:10.1093 / oxfordjournals.pcp.a029575.
- ^ Batge SL, Ross JJ, Reid JB (1999). „Hladiny kyseliny abscisové v semenech mutantu lh-2 s nedostatkem gibberellinu hrachu (Pisum sativum)“. Physiologia Plantarum. 195 (3): 485–490. doi:10.1034 / j.1399-3054.1999.105313.x.
- ^ White CN, Proebsting WM, Hedden P, Rivin CJ (duben 2000). "Gibberelliny a vývoj semen v kukuřici. I. Důkazy, že rovnováha giberelinu a kyseliny abscisové řídí klíčivost versus způsoby zrání". Fyziologie rostlin. 122 (4): 1081–8. doi:10.1104 / pp.122.4.1081. PMC 58942. PMID 10759503.
- ^ Gazzarrini S, Tsuchiya Y, Lumba S, Okamoto M, McCourt P (září 2004). „Transkripční faktor FUSCA3 řídí načasování vývoje u Arabidopsis prostřednictvím hormonů gibberellin a kyselina abscisová“. Vývojová buňka. 7 (3): 373–85. doi:10.1016 / j.devcel.2004.06.017. PMID 15363412.
- ^ A b Ueguchi-Tanaka M, Nakajima M, Katoh E, Ohmiya H, Asano K, Saji S, Hongyu X, Ashikari M, Kitano H, Yamaguchi I, Matsuoka M (červenec 2007). „Molekulární interakce rozpustného gibberellinového receptoru, GID1, s rýžovým proteinem DELLA, SLR1 a giberelinem“. Rostlinná buňka. 19 (7): 2140–55. doi:10.1105 / tpc.106.043729. PMC 1955699. PMID 17644730.
- ^ A b Murase K, Hirano Y, Sun TP, Hakoshima T (listopad 2008). „Rozpoznávání DELLA vyvolané gibberellinem receptorem gibberellinu GID1“. Příroda. 456 (7221): 459–63. Bibcode:2008Natur.456..459M. doi:10.1038 / nature07519. PMID 19037309. S2CID 16280595.
- ^ A b Shimada A, Ueguchi-Tanaka M, Nakatsu T, Nakajima M, Naoe Y, Ohmiya H, Kato H, Matsuoka M (listopad 2008). "Strukturální základna pro rozpoznávání giberelinu podle jeho receptoru GID1". Příroda. 456 (7221): 520–3. Bibcode:2008Natur.456..520S. doi:10.1038 / nature07546. PMID 19037316. S2CID 205215510.
- ^ Achard P, Genschik P (2009). „Uvolnění brzd růstu rostlin: jak GAs vypínají proteiny DELLA“. Journal of Experimental Botany. 60 (4): 1085–92. doi:10.1093 / jxb / ern301. PMID 19043067.
- ^ A b C Davière JM, Achard P (březen 2013). „Gibberellinová signalizace v rostlinách“. Rozvoj. 140 (6): 1147–51. doi:10,1242 / dev.087650. PMID 23444347.
- ^ A b Lechner E, Achard P, Vansiri A, Potuschak T, Genschik P (prosinec 2006). "F-box proteiny všude". Aktuální názor na biologii rostlin. 9 (6): 631–8. doi:10.1016 / j.pbi.2006.09.003. PMID 17005440.
- ^ McGinnis KM, Thomas SG, Soule JD, Strader LC, Zale JM, Sun TP, Steber CM (květen 2003). „Arabidopsis SLEEPY1 gen kóduje domnělou F-box podjednotku SCF E3 ubikvitin ligázy“. Rostlinná buňka. 15 (5): 1120–30. doi:10.1105 / tpc.010827. PMC 153720. PMID 12724538.
- ^ A b Zheng Y, Gao Z, Zhu Z (říjen 2016). „Moduly DELLA-PIF: Old Dogs Learn New Tricks“. Trendy ve vědě o rostlinách. 21 (10): 813–815. doi:10.1016 / j.tplantts.2016.08.006. PMID 27569991.
- ^ Oh E, Zhu JY, Bai MY, Arenhart RA, Sun Y, Wang ZY (květen 2014). „Prodloužení buněk je regulováno centrálním okruhem interagujících transkripčních faktorů v hypokotylu Arabidopsis“. eLife. 3. doi:10,7554 / eLife.03031. PMC 4075450. PMID 24867218.
- ^ Marín-de la Rosa N, Sotillo B, Miskolczi P, Gibbs DJ, Vicente J, Carbonero P, Oñate-Sánchez L, Holdsworth MJ, Bhalerao R, Alabadí D, Blázquez MA (říjen 2014). „Rozsáhlá identifikace transkripčních faktorů souvisejících s gibberellinem definuje FAKTORY ETHYLENOVÉ ODPOVĚDI skupiny VII jako funkční partnery DELLA“. Fyziologie rostlin. 166 (2): 1022–32. doi:10.1104 / pp.114.244723. PMC 4213073. PMID 25118255.
- ^ Locascio A, Blázquez MA, Alabadí D (květen 2013). „Dynamická regulace organizace kortikálních mikrotubulů prostřednictvím prefoldin-DELLA interakce“. Aktuální biologie. 23 (9): 804–9. doi:10.1016 / j.cub.2013.03.053. PMID 23583555.
- ^ Salanenka Y, Verstraeten I, Löfke C, Tabata K, Naramoto S, Glanc M, Friml J (duben 2018). „Signalizace gibberellinu DELLA se zaměřuje na komplex retromeru k přesměrování přenosu proteinů na plazmatickou membránu“. Sborník Národní akademie věd Spojených států amerických. 115 (14): 3716–3721. doi:10.1073 / pnas.1721760115. PMC 5889667. PMID 29463731.
externí odkazy
- Gibberellin v DataBase vlastností pesticidů (PPDB)