BAR doména - BAR domain
| BAR doména | |||||||||
|---|---|---|---|---|---|---|---|---|---|
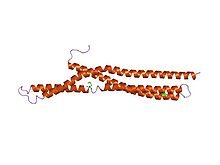 Struktura BAR amfifyzinu.[1] | |||||||||
| Identifikátory | |||||||||
| Symbol | BAR | ||||||||
| Pfam | PF03114 | ||||||||
| InterPro | IPR004148 | ||||||||
| CHYTRÝ | SM00721 | ||||||||
| STRÁNKA | PDOC51021 | ||||||||
| SCOP2 | 1uru / Rozsah / SUPFAM | ||||||||
| CDD | cd07307 | ||||||||
| |||||||||
V molekulární biologii BAR domény jsou vysoce konzervované domény dimerizace proteinů, které se vyskytují v mnoha proteinech zapojených do membránové dynamiky v buňce. Doména BAR je banánového tvaru a váže se na membránu prostřednictvím své konkávní tváře. Je schopen snímat zakřivení membrány tak, že se přednostně váže na zakřivené membrány. BAR domény jsou pojmenovány po třech proteinech, ve kterých se nacházejí: Bin, Amfifyzin a Rvs.
Domény BAR se vyskytují v kombinaci s jinými doménami
Mnoho proteinů rodiny BAR obsahuje alternativní domény lipidové specificity, které pomáhají cílit tyto proteiny na konkrétní membránové oddíly. Někteří také mají SH3 domény které se vážou k dynamin a tak proteiny jako amfifyzin a endofilin jsou zapojeny do orchestrace štěpení vezikul.
N-BAR doména
Některé proteiny obsahující doménu BAR mají N-koncovou amfipatickou šroubovici před doménou BAR. Tato šroubovice se vloží (jako v epsinové doméně ENTH) do membrány a indukuje zakřivení, které je stabilizováno BAR dimerem. Amfifyzin, endofilin, BRAP1 / bin2 a nadrin jsou příklady takových proteinů obsahujících N-BAR. Drosophila amphiphysin N-BAR (DA-N-BAR) je příkladem proteinu s preferencí negativně nabitých povrchů.[1]
F-BAR (EFC) doména
F-BAR domény (pro FCH-BAR nebo EFC pro rozšířenou FCH homologii) jsou BAR domény, které jsou rozšířením již zavedené FCH domény. Často se nacházejí na amino konci proteinů. Mohou vázat lipidové membrány a mohou tubulovat lipidy in vitro a in vivo, ale jejich přesná fyziologická role je stále předmětem výzkumu.[2] Příklady rodiny domén F-BAR jsou CIP4 / FBP17 / Toca-1, Syndapiny (také nazývané PACSIN) a munisciny. Genový knock-out syndapinu I u myší odhalil, že tato mozkem obohacená izoforma rodiny syndapinu je klíčová pro správnou kontrolu velikosti synaptických vezikul a tím skutečně pomáhá definovat zakřivení membrány fyziologický proces. Práce laboratoře Britty Qualmann také prokázala, že syndapin I je zásadní pro správné zacílení na velkou GTPázu dynamin na membrány.[3]
Třídění nexinů
The třídění nexinu rodina proteinů zahrnuje několik členů, kteří mají doménu BAR, včetně dobře charakterizované SNX1 a SNX9.
Lidské proteiny obsahující tuto doménu
AMPH; ARHGAP17; BIN1; BIN2; BIN3; DNMBP; GMIP; RICH2; SH3BP1;SH3GL1; SH3GL2; SH3GL3; SH3GLB1; SH3GLB2;
Viz také
externí odkazy
Reference
- ^ A b Peter BJ, Kent HM, Mills IG a kol. (Leden 2004). "BAR domény jako senzory zakřivení membrány: amfifyzinová BAR struktura". Věda. 303 (5657): 495–9. doi:10.1126 / science.1092586. PMID 14645856. S2CID 6104655.
- ^ Qualmann B, Koch D, Kessels MM (srpen 2011). „Pojďme banány: přehodnocení endocytického BAR kódu“. EMBO J.. 30 (17): 3501–15. doi:10.1038 / emboj.2011.266. PMC 3181480. PMID 21878992.
- ^ Koch D, Spiwoks-Becker I, Sabanov V, Sinning A, Dugladze T, Stellmacher A, Ahuja R, Grimm J, Schüler S, Müller A, Angenstein F, Ahmed T, Diesler A, Moser M, Tom Dieck S, Spessert R , Boeckers TM, Fässler R, Hübner CA, Balschun D, Gloveli T, Kessels MM, Qualmann B (prosinec 2011). „Správná tvorba synaptických vezikul a aktivita neuronové sítě se zásadně spoléhají na syndapin I“. EMBO J.. 30 (24): 4955–69. doi:10.1038 / emboj.2011.339. PMC 3243622. PMID 21926968.
Další čtení
- Leventis PA, Chow BM, Stewart BA, Iyengar B, Campos AR, Boulianne GL (listopad 2001). „Drosophila Amphiphysin je postsynaptický protein potřebný pro normální pohyb, ale nikoli endocytózu“. Provoz. 2 (11): 839–50. doi:10.1034 / j.1600-0854.2001.21113.x. PMID 11733051.
- Zhang B, Zelhof AC (červenec 2002). „Amfifyziny: zvýšení BAR pro recyklaci synaptických vezikul a dynamiku membrány. Bin-Amphiphysin-Rvsp“. Provoz. 3 (7): 452–60. doi:10.1034 / j.1600-0854.2002.30702.x. PMID 12047553.Posouzení.
- Zelhof AC, Bao H, Hardy RW, Razzaq A, Zhang B, Doe CQ (prosinec 2001). „Drosophila Amphiphysin se podílí na lokalizaci proteinů a membránové morfogenezi, ale ne na synaptickou vesikulární endocytózu“. Rozvoj. 128 (24): 5005–15. PMID 11748137.
- Mathew D, Popescu A, Budnik V (listopad 2003). „Funkce Drosophila amphiphysin během synaptického cyklování membrány Fasciclin II“. J. Neurosci. 23 (33): 10710–6. doi:10.1523 / JNEUROSCI.23-33-10710.2003. PMC 6740931. PMID 14627656.
- Peter BJ, Kent HM, Mills IG a kol. (Leden 2004). "BAR domény jako senzory zakřivení membrány: amfifyzinová BAR struktura". Věda. 303 (5657): 495–9. doi:10.1126 / science.1092586. PMID 14645856. S2CID 6104655.
- Weissenhorn W (srpen 2005). "Krystalová struktura domény BAR endofilin-A1". J. Mol. Biol. 351 (3): 653–61. doi:10.1016 / j.jmb.2005.06.013. PMID 16023669.
- Gallop JL, Jao CC, Kent HM a kol. (Červen 2006). "Mechanismus zakřivení membrány zprostředkované doménou N-BAR endofilinem". EMBO J.. 25 (12): 2898–910. doi:10.1038 / sj.emboj.7601174. PMC 1500843. PMID 16763559.
- Masuda M, Takeda S, Sone M a kol. (Červen 2006). „Doména endofilin BAR řídí zakřivení membrány dvěma nově identifikovanými mechanismy založenými na struktuře“. EMBO J.. 25 (12): 2889–97. doi:10.1038 / sj.emboj.7601176. PMC 1500852. PMID 16763557.
- Frost A, Perera R, Roux A a kol. (Březen 2008). "Strukturní základ membránové invaginace doménami F-BAR". Buňka. 132 (5): 807–17. doi:10.1016 / j.cell.2007.12.041. PMC 2384079. PMID 18329367.