Sluchový systém - Auditory system
| Sluchový systém | |
|---|---|
| Anatomická terminologie |

The sluchový systém je senzorický systém pro smysl sluch. Zahrnuje jak smyslové orgány (uši) a sluchové části senzorický systém.
Přehled systému
Trychtýře vnějšího ucha zvuk vibrace na ušní bubínek, což zvyšuje akustický tlak ve středofrekvenčním rozsahu. Ossicles středního ucha dále zhruba 20krát zesilují vibrační tlak. Základna svorek spojuje vibrace do kochle přes oválné okénko, které vibruje kapalinu perilymfou (přítomnou v celém vnitřním uchu) a způsobuje, že se kulaté okénko vypoukne, když se oválné okénko vyboulí dovnitř.
Vestibulární a tympanické kanály jsou naplněny perilymfou a menší kochleární kanál mezi nimi je naplněn endolymfou, tekutinou s velmi odlišnou koncentrací a napětím iontů.[1][2][3] Vibrace vestibulárního potrubí perilymfy ohýbají orgán vnějších buněk Corti (4 řádky), což způsobuje uvolňování prestinu ve špičkách buněk. To způsobí, že buňky budou chemicky prodlouženy a zmenšeny (somatický motor) a vlasové svazky se posunou, což zase elektricky ovlivní pohyb bazilární membrány (motor vlasových svazků). Tyto motory (vnější vlasové buňky) zesilují amplitudy pohyblivých vln více než 40krát.[4] Vnější vlasové buňky (OHC) jsou minimálně inervovány spirálním gangliem v pomalých (nemyelinizovaných) vzájemných komunikačních svazcích (30+ vlasů na nervové vlákno); to kontrastuje s vnitřními vlasovými buňkami (IHC), které mají pouze aferentní inervaci (30+ nervových vláken na jeden vlas), ale jsou silně spojeny. Existuje třikrát až čtyřikrát tolik OHC než IHC. Bazilární membrána (BM) je bariéra mezi šupinami, podél jejichž okraje sedí IHC a OHC. Šířka a tuhost bazilární membrány se mění, aby řídila frekvence nejlépe snímané IHC. Na kochleární základně je BM nejužší a nejpevnější (vysoké frekvence), zatímco na kochleárním vrcholu je nejširší a nejméně tuhá (nízké frekvence). Tektoriální membrána (TM) pomáhá usnadnit kochleární zesílení stimulací OHC (přímá) a IHC (prostřednictvím vibrací endolymfy). Šířka a tuhost TM se vyrovná BM a podobně pomáhá při frekvenční diferenciaci.[5][6][7][8][9][10][11][12][13]
The vynikající olivový komplex (SOC), v mostech, je první konvergence levého a pravého kochleárního pulzu. SOC má 14 popsaných jader; zde se používá jejich zkratka (viz Vynikající olivový komplex pro jejich plná jména). MSO určuje úhel, ze kterého zvuk vycházel, měřením časových rozdílů v levé a pravé informaci. LSO normalizuje hladinu zvuku mezi ušima; používá zvukové intenzity k určení zvukového úhlu. LSO inervuje IHC. VNTB inervuje OHC. MNTB inhibuje LSO prostřednictvím glycinu. LNTB jsou imunní vůči glycinu a používají se pro rychlou signalizaci. DPO jsou vysokofrekvenční a tonotopická. DLPO jsou nízkofrekvenční a tonotopické. VLPO mají stejnou funkci jako DPO, ale působí v jiné oblasti. PVO, CPO, RPO, VMPO, ALPO a SPON (inhibované glycinem) jsou různá signální a inhibiční jádra.[14][15][16][17]
Lichoběžníkové tělo je místo, kde se většina vláken kochleárního jádra (CN) rozpadá (kříž zleva doprava a naopak); tento kříž pomáhá při zvukové lokalizaci.[18] CN se dělí na ventrální (VCN) a hřbetní (DCN) regiony. VCN má tři jádra.[je zapotřebí objasnění ] Husté buňky přenášejí časovací informace, jejich tvarové průměry časují nervozitu. Stellate (chopper) buňky kódují zvuková spektra (vrcholy a údolí) rychlostmi prostorového nervového výboje na základě síly vstupního zvuku (spíše než frekvence). Buňky chobotnice mají při střelbě téměř nejlepší časovou přesnost, dekódují sluchový časovací kód. DCN má 2 jádra. DCN také přijímá informace z VCN. Fusiformní buňky integrují informace k určení spektrálních signálů k místům (například zda zvuk vycházel zepředu nebo zezadu). Kochleární nervová vlákna (více než 30 000) mají nejcitlivější frekvenci a reagují v širokém rozsahu úrovní.[19][20]
Zjednodušené signály nervových vláken jsou transportovány hustými buňkami do binaurálních oblastí v olivový komplex, zatímco signální vrcholy a údolí jsou zaznamenány hvězdicovými buňkami a načasování signálu je extrahováno buňkami chobotnice. Postranní lemniscus má tři jádra: hřbetní jádra nejlépe reagují na bilaterální vstup a mají reakce vyladěné podle složitosti; střední jádra mají široké ladicí reakce; a ventrální jádra mají široké a středně složité ladicí křivky. Ventrální jádra laterálního lemnisku pomáhají inferior colliculus (IC) dekódovat zvuky modulované amplitudou tím, že dávají jak fázické, tak tonické odpovědi (krátké, respektive dlouhé noty). IC přijímá vstupy, které nejsou zobrazeny, včetně vizuálních (pretektální oblast: pohání oči ke zvuku. Superior colliculus: orientace a chování k objektům, stejně jako pohyby očí (saccade)), pons (vynikající mozeček stopky: thalamus na mozeček připojení / slyšet zvuk a učit se odezvu chování), míchu (periaqueductal šedá: slyšet zvuk a instinktivně se pohybovat) a thalamus. Výše uvedené je to, co implikuje IC v „úlekové reakci“ a očních reflexech. Kromě multisenzorické integrace IC reaguje na konkrétní frekvence modulace amplitudy, což umožňuje detekci výšky tónu. IC také určuje časové rozdíly v binaurálním sluchu.[21] Mediální geniculární jádro se dělí na ventrální (reléové a relé-inhibiční buňky: frekvence, intenzita a binaurální informace topograficky předávané), dorzální (široká a složitá vyladěná jádra: spojení se somatosenzorickými informacemi) a střední (široká, komplexní a úzce laděná) jádra: intenzita přenosu a doba trvání zvuku). Sluchová kůra (AC) přináší zvuk do vědomí / vnímání. AC identifikuje zvuky (rozpoznávání zvukového jména) a také identifikuje místo původu zvuku. AC je topografická frekvenční mapa se svazky reagujícími na různé harmonie, načasování a výšku tónu. Pravostranný střídavý proud je citlivější na tonalitu, levý střídavý proud je citlivější na minutové sekvenční rozdíly ve zvuku.[22][23] Rostromediální a ventrolaterální prefrontální kůry se účastní aktivace během tonálního prostoru a ukládání krátkodobých vzpomínek.[24] Heschlův gyrus / transverzální temporální gyrus zahrnuje Wernickeho oblast a funkčnost, je silně zapojen do procesů emocí-zvuk, emoce-mimika a zvuk-paměť. Entorhinální kůra je součástí „systému hipokampu“, který pomáhá a ukládá vizuální a sluchové paměti.[25][26] Supramarginal gyrus (SMG) pomáhá porozumět jazyku a je zodpovědný za soucitné reakce. SMG spojuje zvuky se slovy s hranatým gyrusem a pomáhá při výběru slov. SMG integruje hmatové, vizuální a sluchové informace.[27][28]
Struktura

Vnější ucho
Záhyby chrupavky obklopující zvukovod se nazývají boltce. Zvukové vlny se odráží a zeslabují, když zasáhnou špičku, a tyto změny poskytují další informace, které pomohou mozku určit směr zvuku.
Zvukové vlny vstupují do zvukovod, klamně jednoduchá trubka. Ušní kanál zesiluje zvuky mezi 3 a 12 kHz. The ušní bubínek, na vzdáleném konci zvukovodu označuje začátek střední ucho.
Střední ucho
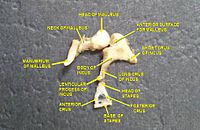
Zvukové vlny procházejí zvukovodem a zasáhnou tympanickou membránu, nebo ušní bubínek. Tato informace o vlně prochází vzduchem naplněnou dutinou středního ucha prostřednictvím řady jemných kostí: Malleus (kladivo), incus (kovadlina) a sponky (třmen). Tyto ossicles působí jako páka a převádí zvukové vibrace nízkotlakého bubínku na vysokotlaké zvukové vibrace v jiné, menší membráně zvané oválné okno nebo vestibulární okno. The manubrium (rukojeť) malleu artikuluje s tympanickou membránou, zatímco stupačka (základna) svorek artikuluje s oválným okénkem. U oválného okénka je nutný vyšší tlak než u typické membrány, protože vnitřní ucho za oválným okénkem obsahuje spíše kapalinu než vzduch. The stapediový reflex svalů středního ucha pomáhá chránit vnitřní ucho před poškozením snížením přenosu zvukové energie, když stapedius sval se aktivuje v reakci na zvuk. Střední ucho stále obsahuje zvukové informace ve formě vln; převádí se na nervové impulsy v kochlea.
Vnitřní ucho
| Kochlea | |
|---|---|
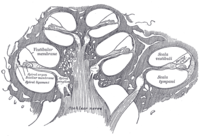 Schematický podélný řez hlemýžďem. The kochleární potrubí nebo Scala média, je označen jako ductus cochlearis vpravo. | |
| Anatomická terminologie |
Vnitřní ucho se skládá z kochlea a několik nesluchových struktur. Hlemýžď má tři části naplněné tekutinou (tj scala media, scala tympani a scala vestibuli), a podporuje fluidní vlnu poháněnou tlakem přes bazilární membrána oddělující dvě části. Je překvapivé, že jedna část, zvaná kochleární potrubí nebo Scala média, obsahuje endolymfa. Endolymfa je tekutina podobného složení jako intracelulární tekutina nacházející se uvnitř buněk. Orgán Corti se nachází v tomto potrubí na bazilární membráně a transformuje mechanické vlny na elektrické signály v neuronech. Další dvě sekce jsou známé jako scala tympani a scala vestibuli. Ty se nacházejí v kostním labyrintu, který je naplněn tekutinou zvanou perilymph, podobné složení jako mozkomíšní mok. Chemický rozdíl mezi tekutinami endolymfa a perilymph tekutiny jsou důležité pro funkci vnitřního ucha kvůli rozdílům elektrického potenciálu mezi ionty draslíku a vápníku.
Půdorys lidské kochley (typický pro všechny savčí a většina obratlovců ) ukazuje, kde se po jeho délce vyskytují specifické frekvence. Frekvence je přibližně exponenciální funkcí délky hlemýždě uvnitř Cortiho varhany. U některých druhů, jako jsou netopýři a delfíni, se vztah rozšiřuje v konkrétních oblastech, aby se podpořila jejich aktivní schopnost sonaru.
Cortiho varhany
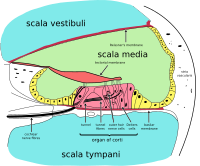
Orgán Corti tvoří pás smyslového epitelu, který běží podélně po celém kochlei Scala média. Jeho vláskové buňky transformují tekutinové vlny na nervové signály. Cesta bezpočtu nervů začíná tímto prvním krokem; odtud další zpracování vede k celé řadě sluchových reakcí a vjemů.
Vlásková buňka
Vlasové buňky jsou sloupcovité buňky, každá se svazkem 100–200 specializovaných řasy nahoře, pro které jsou pojmenováni. Existují dva typy vláskových buněk; vnitřní a vnější vlasy buňky. Vnitřní vláskové buňky jsou mechanoreceptory pro sluch: přenášejí vibrace zvuku na elektrickou aktivitu v nervová vlákna, který se přenáší do mozku. Vnější vláskové buňky jsou motorickou strukturou. Zvuková energie způsobuje změny ve tvaru těchto buněk, což slouží k zesílení zvukových vibrací kmitočtově specifickým způsobem. Lehce odpočívá na nejdelší řasy vnitřních vlasových buněk je tectoriální membrána, který se pohybuje každým cyklem zvuku tam a zpět, naklánění řasinek, což je to, co vyvolává elektrické reakce vlasových buněk.
Vnitřní vláskové buňky, jako fotoreceptorové buňky oka, ukázat a odstupňovaná odpověď, místo toho hroty typické pro jiné neurony. Tyto odstupňované potenciály nejsou vázány vlastnostmi akčního potenciálu „všechny nebo žádné“.
V tomto okamžiku je možné se zeptat, jak takové kroucení vlasového svazku spouští rozdíl v membránovém potenciálu. Současný model je takový, že řasinky jsou navzájem spojeny pomocí „odkazy na tipy ", struktury, které spojují špičky jednoho cilium s druhým. Protahováním a komprimací mohou spojky špičky otevírat iontový kanál a produkovat receptorový potenciál ve vláskové buňce. Nedávno se ukázalo, že kadherin-23 CDH23 a protocadherin-15 PCDH15 jsou adhezní molekuly spojené s těmito odkazy na špičky.[29] Předpokládá se, že a vápník poháněný motor způsobuje zkrácení těchto vazeb k regeneraci napětí. Tato regenerace napětí umožňuje obavu z prodloužené sluchové stimulace.[30]
Neurony
Aferentní neurony inervují kochleární vnitřní vlasové buňky na synapsích, kde je neurotransmiter glutamát komunikuje signály z vlasových buněk do dendritů primárních sluchových neuronů.
V kochle je mnohem méně vnitřních vlasových buněk než aferentní nervová vlákna - mnoho sluchových nervových vláken inervuje každou vlasovou buňku. Nervové dendrity patří k neuronům Sluchový nerv, což se zase připojuje k vestibulární nerv tvořit vestibulocochlear nerv nebo lebeční nerv číslo VIII.[31]Oblast bazilární membrány dodávající vstupy do konkrétního aferentního nervového vlákna lze považovat za jeho receptivní pole.
Při vnímání zvuku hrají roli i eferentní projekce z mozku do hlemýždě, i když tomu není dobře rozuměno. Eferentní synapse se vyskytují na vnějších vlasových buňkách a na aferentních (směrem k mozku) dendritech pod vnitřními vlasovými buňkami
Neuronální struktura
Kochleární jádro
The kochleární jádro je prvním místem neuronového zpracování nově převedených „digitálních“ dat z vnitřního ucha (viz také binaurální fúze ). U savců je tato oblast anatomicky a fyziologicky rozdělena do dvou oblastí: hřbetní kochleární jádro (DCN) a ventrální kochleární jádro (VCN). VCN se dále dělí nervovým kořenem na posteroventrální kochleární jádro (PVCN) a anteroventrální kochleární jádro (AVCN).[32]
Trapézové tělo
The lichoběžníkové tělo je balíček rozhodující vlákna ve ventrálních mostech, která přenášejí informace používané pro binaurální výpočty v mozkovém kmeni. Některé z těchto axony pocházejí z kochleární jádro a přejít na druhou stranu před cestou na nadřazené olivové jádro. Předpokládá se, že to pomůže lokalizace zvuku.[33]
Vynikající olivový komplex
The vynikající olivový komplex se nachází v pons, a přijímá projekce převážně z ventrálního kochleárního jádra, ačkoli tam také vychází projekce dorzálního kochleárního jádra, prostřednictvím ventrální akustické strie. V rámci vynikající olivový komplex leží laterální nadřazená oliva (LSO) a střední nadřazená oliva (MSO). První z nich je důležitý pro detekci rozdílů na úrovni interaurálů, zatímco druhý je důležitý pro rozlišení interaurálních časových rozdílů.[16]
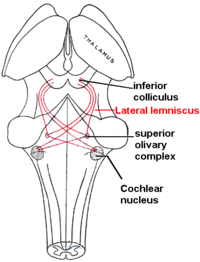
Boční lemniscus
Postranní lemniscus je trakt axony v mozkový kmen který přenáší informace o zvuku z kochleární jádro do různých jader mozkového kmene a nakonec do kontralaterálu inferior colliculus z střední mozek.
Inferior colliculi
Dolní colliculi (IC) se nacházejí těsně pod centry vizuálního zpracování známými jako superior colliculi. Centrální jádro integrovaného obvodu je téměř povinným relé ve vzestupném sluchovém systému a s největší pravděpodobností jedná o integraci informací (konkrétně o lokalizaci zdroje zvuku z vynikající olivový komplex[15] a hřbetní kochleární jádro ) před odesláním do thalamus a kůra.[34] Dolní colliculus také přijímá sestupné vstupy z sluchová kůra a sluchové thalamus (nebo mediální geniculate jádro ). [35]
Mediální geniculární jádro
The mediální geniculate jádro je součástí systému thalamického relé.
Primární sluchová kůra
The primární sluchová kůra je první region z mozková kůra přijímat sluchový vstup.
Vnímání zvuku je spojeno s levým zadním superior temporální gyrus (STG). Nadřazený temporální gyrus obsahuje několik důležitých struktur mozku, včetně Brodmann oblasti 41 a 42 s vyznačením polohy primární sluchová kůra, kortikální oblast odpovědná za vnímání základních charakteristik zvuku, jako je výška tónu a rytmus. Z výzkumu u nelidských primátů víme, že primární sluchovou kůru lze pravděpodobně rozdělit dále na funkčně diferencovatelné podoblasti.[36][37][38][39][40][41][42]Lze předpokládat, že mají neurony primární sluchové kůry receptivní pole pokrývající řadu sluchové frekvence a mají selektivní reakce na harmonické výšky tónu.[43] Neurony integrující informace ze dvou uší mají receptivní pole pokrývající určitou oblast sluchového prostoru.
Primární sluchová kůra je obklopena sekundární sluchovou kůrou a je s ní propojena. Tyto sekundární oblasti se propojují s dalšími zpracovatelskými oblastmi v EU superior temporální gyrus v hřbetní bance superior temporální sulcus a v čelní lalok. U lidí je spojení těchto oblastí s střední temporální gyrus jsou pravděpodobně důležité pro vnímání řeči. Frontotemporální systém, který je základem sluchového vnímání, nám umožňuje rozlišit zvuky jako řeč, hudbu nebo hluk.
Sluchové ventrální a hřbetní proudy
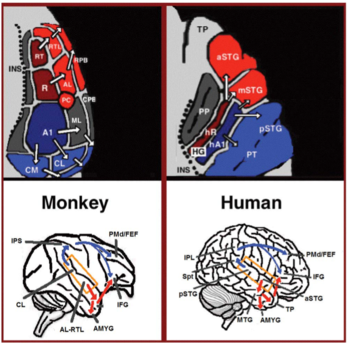
Z primární sluchové kůry vycházejí dvě oddělené cesty: sluchový ventrální proud a sluchový hřbetní proud.[44] Sluchový ventrální proud zahrnuje přední horní temporální gyrus, přední horní temporální sulcus, střední temporální gyrus a temporální pól. Neurony v těchto oblastech jsou odpovědné za zvukové rozpoznání a extrakci významu z vět. Sluchový dorzální proud zahrnuje zadní horní temporální gyrus a sulcus, nižší temenní lalůček a intra-parietální sulcus. Obě cesty se u lidí promítají do horního frontálního gyrusu. Nejvíce zavedenou rolí sluchového dorzálního proudu u primátů je zvuková lokalizace. U lidí je sluchový hřbetní proud v levé hemisféře také zodpovědný za opakování a artikulaci řeči, fonologické dlouhodobé kódování jmen slov a verbální pracovní paměť.
Klinický význam
Správná funkce sluchového systému je vyžadována, aby bylo možné snímat, zpracovávat a rozumět zvuku z okolí. Potíže se snímáním, zpracováním a porozuměním zvukového vstupu mají potenciál nepříznivě ovlivnit schopnost jednotlivce denně komunikovat, učit se a efektivně plnit rutinní úkoly.[45]
U dětí je včasná diagnostika a léčba narušené funkce sluchového systému důležitým faktorem při zajišťování splnění klíčových sociálních, akademických a řečových / jazykových vývojových milníků.[46]
Porucha sluchového systému může zahrnovat některou z následujících možností:
- Sluchová odezva mozkového kmene a ABR audiometrický test pro novorozenecký sluch
- Porucha sluchového zpracování
- Účinky na zdraví způsobené hlukem
- Tinnitus
- Endaurální jevy
Viz také
Reference
- ^ Tillotson JK, McCann S (2013). Kartičky lékařské anatomie Kaplan. Kaplan Publishing. ISBN 978-1-60714-984-2.
- ^ Ashwell K (2016). Barronovy anatomické flash karty. Barronova vzdělávací série. ISBN 978-1-4380-7717-8.
- ^ „Jak funguje můj sluch?“. Audiologická společnost NZ. Archivovány od originál dne 23. srpna 2019. Citováno 27. března 2016.
- ^ Zheng J, Shen W, He DZ, Long KB, Madison LD, Dallos P (květen 2000). „Prestin je motorický protein kochleárních vnějších vlasových buněk“. Příroda. 405 (6783): 149–55. Bibcode:2000Natur.405..149Z. doi:10.1038/35012009. PMID 10821263.
- ^ Zwislocki JJ, Cefaratti LK (listopad 1989). „Tectorial membrána. II: Měření tuhosti in vivo“. Výzkum sluchu. 42 (2–3): 211–27. doi:10.1016/0378-5955(89)90146-9. PMID 2606804.
- ^ Richter CP, Emadi G, Getnick G, Quesnel A, Dallos P (září 2007). "Tektoriální přechody tuhosti membrány". Biofyzikální deník. 93 (6): 2265–76. Bibcode:2007BpJ .... 93.2265R. doi:10.1529 / biophysj.106.094474. PMC 1959565. PMID 17496047.
- ^ Meaud J, Grosh K (březen 2010). "Efekt tectoriální membrány a podélné vazby bazilární membrány v kochleární mechanice". The Journal of the Acoustical Society of America. 127 (3): 1411–21. Bibcode:2010ASAJ..127.1411M. doi:10.1121/1.3290995. PMC 2856508. PMID 20329841.
- ^ Gueta R, Barlam D, Shneck RZ, Rousso I (říjen 2006). "Měření mechanických vlastností izolované tektoriální membrány pomocí mikroskopie atomové síly". Sborník Národní akademie věd Spojených států amerických. 103 (40): 14790–5. Bibcode:2006PNAS..10314790G. doi:10.1073 / pnas.0603429103. PMC 1595430. PMID 17001011.
- ^ Freeman DM, Abnet CC, Hemmert W, Tsai BS, Weiss TF (červen 2003). Msgstr "Dynamické materiálové vlastnosti tektorální membrány: shrnutí". Výzkum sluchu. 180 (1–2): 1–10. doi:10.1016 / S0378-5955 (03) 00073-X. PMID 12782348.
- ^ Legan PK, Lukashkina VA, Goodyear RJ, Kössi M, Russell IJ, Richardson GP (říjen 2000). „Cílená delece v alfa-tektorinu odhaluje, že pro získání a načasování kochleární zpětné vazby je nutná tektorální membrána.“ Neuron. 28 (1): 273–85. doi:10.1016 / S0896-6273 (00) 00102-1. PMID 11087000.
- ^ Canlon B (1988). „Účinek akustického traumatu na tectoriální membránu, stereocilii a citlivost sluchu: možné mechanismy poškození, zotavení a ochrany“. Skandinávská audiologie. Supplementum. 27: 1–45. PMID 3043645.
- ^ Zwislocki JJ (1979). "Tectorial membrána: možný ostřící účinek na frekvenční analýzu v hlemýždi". Acta Oto-Laryngologica. 87 (3–4): 267–9. doi:10.3109/00016487909126419. PMID 443008.
- ^ Teudt IU, Richter CP (říjen 2014). "Tuhost bazilární membrány a tectoriální membrány u myši CBA / CaJ". Časopis Asociace pro výzkum v otolaryngologii. 15 (5): 675–94. doi:10.1007 / s10162-014-0463-r. PMC 4164692. PMID 24865766.
- ^ Thompson AM, Schofield BR (listopad 2000). "Aferentní projekce vynikajícího olivárského komplexu". Mikroskopický výzkum a technika. 51 (4): 330–54. doi:10.1002 / 1097-0029 (20001115) 51: 4 <330 :: AID-JEMT4> 3.0.CO; 2-X. PMID 11071718.
- ^ A b Oliver DL (listopad 2000). "Vzestupné eferentní projekce vynikajícího olivárského komplexu". Mikroskopický výzkum a technika. 51 (4): 355–63. doi:10.1002 / 1097-0029 (20001115) 51: 4 <355 :: AID-JEMT5> 3.0.CO; 2-J. PMID 11071719.
- ^ A b Moore JK (listopad 2000). "Organizace lidského vyššího olivárského komplexu". Mikroskopický výzkum a technika. 51 (4): 403–12. doi:10.1002 / 1097-0029 (20001115) 51: 4 <403 :: AID-JEMT8> 3.0.CO; 2-Q. PMID 11071722.
- ^ Yang L, Monsivais P, Rubel EW (březen 1999). „Nadřazené olivární jádro a jeho vliv na nucleus laminaris: zdroj inhibiční zpětné vazby pro detekci shody náhod v mozkovém kmeni ptáků“. The Journal of Neuroscience. 19 (6): 2313–25. doi:10.1523 / JNEUROSCI.19-06-02313.1999. PMC 6782562. PMID 10066281.
- ^ Paolini AG, FitzGerald JV, Burkitt AN, Clark GM (září 2001). „Časové zpracování ze sluchového nervu do mediálního jádra lichoběžníkového těla u krysy“. Výzkum sluchu. 159 (1–2): 101–16. doi:10.1016 / S0378-5955 (01) 00327-6. PMID 11520638.
- ^ Bajo VM, Merchán MA, Malmierca MS, Nodal FR, Bjaalie JG (květen 1999). "Topografická organizace hřbetního jádra laterálního lemnisku u kočky". The Journal of Comparative Neurology. 407 (3): 349–66. doi:10.1002 / (SICI) 1096-9861 (19990510) 407: 3 <349 :: AID-CNE4> 3.0.CO; 2-5. PMID 10320216.
- ^ Young ED, Davis KA (2002). "Obvody a funkce hřbetního kochleárního jádra". V Oertel D, Fay RR, Popper AN (eds.). Integrační funkce v savčí sluchové dráze. Springer Handbook of Auditory Research. 15. New York, NY: Springer. 160–206. doi:10.1007/978-1-4757-3654-0_5. ISBN 978-1-4757-3654-0.
- ^ Oliver DL (2005). "Neuronální organizace v podřízeném colliculus". Ve Winer JA, Schreiner CE (eds.). Nižší colliculus. New York, NY: Springer. str. 69–114. doi:10.1007/0-387-27083-3_2. ISBN 978-0-387-27083-8.
- ^ Janata P, Birk JL, Van Horn JD, Leman M, Tillmann B, Bharucha JJ (prosinec 2002). "Kortikální topografie tonálních struktur, které jsou základem západní hudby". Věda. 298 (5601): 2167–70. Bibcode:2002Sci ... 298.2167J. doi:10.1126 / science.1076262. PMID 12481131.
- ^ Morosan P, Rademacher J, Schleicher A, Amunts K, Schormann T, Zilles K (duben 2001). "Primární lidská sluchová kůra: cytoarchitektonické dělení a mapování do prostorového referenčního systému". NeuroImage. 13 (4): 684–701. CiteSeerX 10.1.1.420.7633. doi:10,1006 / nimg.2000.0715. PMID 11305897.
- ^ Romanski LM, Tian B, Fritz J, Mishkin M, Goldman-Rakic PS, Rauschecker JP (prosinec 1999). „Duální proudy sluchových aferentů cílí na více domén v prefrontální kůře primátů“. Přírodní neurovědy. 2 (12): 1131–6. doi:10.1038/16056. PMC 2778291. PMID 10570492.
- ^ Badre D, Wagner AD (říjen 2007). "Levá ventrolaterální prefrontální kůra a kognitivní kontrola paměti". Neuropsychologie. 45 (13): 2883–901. doi:10.1016 / j.neuropsychologia.2007.06.015. PMID 17675110.
- ^ Amunts K, Kedo O, Kindler M, Pieperhoff P, Mohlberg H, Shah NJ, Habel U, Schneider F, Zilles K (prosinec 2005). „Cytoarchitektonické mapování lidské amygdaly, hipokampální oblasti a entorhinální kůry: mezisubjektová variabilita a mapy pravděpodobnosti“. Anatomie a embryologie. 210 (5–6): 343–52. doi:10.1007 / s00429-005-0025-5. PMID 16208455.
- ^ Penniello MJ, Lambert J, Eustache F, Petit-Taboué MC, Barré L, Viader F, Morin P, Lechevalier B, Baron JC (červen 1995). „PET studie funkční neuroanatomie poruchy psaní při Alzheimerově chorobě. Role levého supramarginálního a levého úhlového gyri“. Brain: Journal of Neurology. 118 (Pt 3) (3): 697–706. doi:10.1093 / mozek / 118.3.697. PMID 7600087.
- ^ Stoeckel C, Gough PM, Watkins KE, Devlin JT (říjen 2009). „Zapojení Supramarginal gyrus do vizuálního rozpoznávání slov“. Kůra; Časopis věnovaný studiu nervového systému a chování. 45 (9): 1091–6. doi:10.1016 / j.cortex.2008.12.004. PMC 2726132. PMID 19232583.
- ^ Lelli A, Kazmierczak P, Kawashima Y, Müller U, Holt JR (srpen 2010). „Vývoj a regenerace senzorické transdukce ve sluchových vlasových buňkách vyžaduje funkční interakci mezi kadherinem-23 a protocadherinem-15“. The Journal of Neuroscience. 30 (34): 11259–69. doi:10.1523 / JNEUROSCI.1949-10.2010. PMC 2949085. PMID 20739546.
- ^ Peng AW, Salles FT, Pan B, Ricci AJ (listopad 2011). „Integrace biofyzikálních a molekulárních mechanismů sluchové mechanotransdukce vlasových buněk“. Příroda komunikace. 2: 523. Bibcode:2011NatCo ... 2..523P. doi:10.1038 / ncomms1533. PMC 3418221. PMID 22045002.
- ^ Meddean - CN VIII. Vestibulocochlear Nerve
- ^ Middle Brooks JC (2009). "Sluchový systém: centrální cesty". V Squire LF (ed.). Encyclopedia of Neuroscience. Akademický tisk. str. 745–752, zde: str. 745 f. ISBN 978-0-08-044617-2.
- ^ Mendoza JE (2011). "Trapézové tělo". In Kreutzer JS, DeLuca J, Caplan B (eds.). Encyklopedie klinické neuropsychologie. New York: Springer. str. 2549. doi:10.1007/978-0-387-79948-3_807. ISBN 978-0-387-79947-6.
- ^ Demanez JP, Demanez L (2003). "Anatomofyziologie centrálního sluchového nervového systému: základní pojmy". Acta Oto-Rhino-Laryngologica Belgica. 57 (4): 227–36. PMID 14714940.
- ^ Schreiner, Christoph (2005). Nižší colliculus. Springer Science + Business Media, Incorporated.
- ^ Pandya DN (1995). "Anatomie sluchové kůry". Revue Neurologique. 151 (8–9): 486–94. PMID 8578069.
- ^ Kaas JH, Hackett TA (1998). "Členění sluchové kůry a úrovně zpracování u primátů". Audiologie a neuro-otologie. 3 (2–3): 73–85. doi:10.1159/000013783. PMID 9575378.
- ^ Kaas JH, Hackett TA, Tramo MJ (duben 1999). "Sluchové zpracování v mozkové kůře primátů". Aktuální názor v neurobiologii. 9 (2): 164–70. doi:10.1016 / S0959-4388 (99) 80022-1. PMID 10322185.
- ^ Kaas JH, Hackett TA (říjen 2000). "Členění sluchové kůry a zpracování proudů u primátů". Sborník Národní akademie věd Spojených států amerických. 97 (22): 11793–9. Bibcode:2000PNAS ... 9711793K. doi:10.1073 / pnas.97.22.11793. PMC 34351. PMID 11050211.
- ^ Hackett TA, Preuss TM, Kaas JH (prosinec 2001). "Architektonická identifikace oblasti jádra ve sluchové kůře makaků, šimpanzů a lidí". The Journal of Comparative Neurology. 441 (3): 197–222. doi:10.1002 / cne.1407. PMID 11745645.
- ^ Scott SK, Johnsrude IS (únor 2003). "Neuroanatomická a funkční organizace vnímání řeči". Trendy v neurovědách. 26 (2): 100–7. CiteSeerX 10.1.1.323.8534. doi:10.1016 / S0166-2236 (02) 00037-1. PMID 12536133.
- ^ Tian B, Reser D, Durham A, Kustov A, Rauschecker JP (duben 2001). "Funkční specializace na sluchovou kůru opice rhesus". Věda. 292 (5515): 290–3. Bibcode:2001Sci ... 292..290T. doi:10.1126 / science.1058911. PMID 11303104.
- ^ Wang X (prosinec 2013). „Harmonická organizace sluchové kůry“. Frontiers in Systems Neuroscience. 7: 114. doi:10.3389 / fnsys.2013.00114. PMC 3865599. PMID 24381544.
- ^ Hickok G, Poeppel D (květen 2007). "Kortikální organizace zpracování řeči". Recenze přírody. Neurovědy. 8 (5): 393–402. doi:10.1038 / nrn2113. PMID 17431404.
- ^ „Ztráta sluchu“ (PDF). HearingLoss.org. National Academy on an Aging Society. Citováno 28. ledna 2018.
- ^ Ciorba A, Corazzi V, Negossi L, Tazzari R, Bianchini C, Aimoni C (prosinec 2017). „Středně těžká ztráta sluchu u dětí: diagnostická a rehabilitační výzva“. The Journal of International Advanced Otology. 13 (3): 407–413. doi:10.5152 / iao.2017.4162. PMID 29360094.
Další čtení
- Kandel, Eric R. (2012). Principy neurální vědy. New York: McGraw-Hill. ISBN 978-0-07-139011-8. OCLC 795553723.
externí odkazy
- Promenáda kolem kochley
- Sluchový systém - Výuka neurověd Washingtonské univerzity
- Lincoln Gray. „Kapitola 13: Sluchový systém: Cesty a reflexe“. Neuroscience Online, otevřená elektronická učebnice neurovědy. University of Texas Health Science Center v Houstonu (UTHealth). Archivovány od originál dne 12. 11. 2016. Citováno 27. dubna 2014.