Zpracování jazyka v mozku - Language processing in the brain
Tento článek má několik problémů. Prosím pomozte vylepši to nebo diskutovat o těchto otázkách na internetu diskusní stránka. (Zjistěte, jak a kdy tyto zprávy ze šablony odebrat) (Zjistěte, jak a kdy odstranit tuto zprávu šablony)
|
Zpracování jazyka odkazuje na způsob, jakým lidé používají slova ke sdělování myšlenek a pocitů, a na to, jak jsou tyto komunikace zpracovávány a chápány. Zpracování jazyka je považováno za jedinečně lidskou schopnost, která není produkována se stejným gramatickým porozuměním nebo systematičností ani u nejbližších příbuzných primátů.[1]
Po celé 20. století dominantní model[2] pro zpracování jazyka v mozku byla Geschwind-Lichteim-Wernicke model, který je založen především na analýze pacientů s poškozením mozku. Avšak díky zdokonalení intrakortikálních elektrofyziologických záznamů opičích a lidských mozků a také neinvazivním technikám, jako jsou fMRI, PET, MEG a EEG, je duální sluchová cesta[3][4] bylo odhaleno. V souladu s tímto modelem existují dvě cesty, které spojují sluchovou kůru s frontálním lalokem, přičemž každá cesta odpovídá za různé jazykové role. The sluchový ventrální proud cesta je zodpovědná za rozpoznávání zvuku a je proto známá jako sluchová cesta „co“. The sluchový hřbetní proud u člověka i jiných subhumánních primátů zvuková lokalizace, a je proto známá jako sluchová cesta „kde“. U lidí je tato cesta (zejména v levé hemisféře) také zodpovědná za produkci řeči, opakování řeči, čtení rtů a fonologickou pracovní paměť a dlouhodobou paměť. V souladu s modelem jazykové evoluce „odkud do čeho“.[5][6] důvodem, proč se ADS vyznačuje tak širokou škálou funkcí, je to, že každá označuje jinou fázi vývoje jazyka.
K rozdělení dvou proudů dochází nejprve v Sluchový nerv kde přední větev vstupuje do přední kochleární jádro v mozkovém kmeni, který vede ke sluchovému ventrálnímu proudu. Zadní větev vstupuje do dorzálního a posteroventrálního kochleárního jádra za vzniku sluchového dorzálního proudu.[7]:8
Ke zpracování jazyka může dojít také ve vztahu k podepsané jazyky nebo písemný obsah.
Rané neurolingvistické modely

Po celé 20. století dominovaly našim znalostem zpracování jazyka v mozku model Wernicke-Lichtheim-Geschwind.[8][2][9] Wernicke-Lichtheim-Geschwindův model je primárně založen na výzkumu prováděném na jedincích s poškozením mozku, u nichž se údajně vyskytly různé poruchy související s jazykem. V souladu s tímto modelem jsou slova vnímána prostřednictvím specializovaného centra pro příjem slov (Wernickeho oblast ), který se nachází vlevo temporoparietální spojení. Tento region poté promítne do centra produkce slov (Brocova oblast ), který se nachází vlevo dolní čelní gyrus. Protože se předpokládalo, že téměř veškerý jazykový vstup trychtýřuje přes Wernickovu oblast a veškerý jazykový výstup trychtýřově přes Brocovu oblast, bylo extrémně obtížné identifikovat základní vlastnosti každého regionu. Tento nedostatek jasné definice příspěvku regionů Wernicke a Broca k lidské řeči způsobil, že bylo nesmírně obtížné identifikovat jejich homology u jiných primátů.[10] S příchodem MRI a jeho aplikací na mapování lézí se však ukázalo, že tento model je založen na nesprávných korelacích mezi příznaky a lézemi.[11][12][13][14][15][16][17] Vyvrácení tak vlivného a dominantního modelu otevřelo dveře novým modelům zpracování jazyka v mozku.
Současné neurolingvistické modely
Anatomie
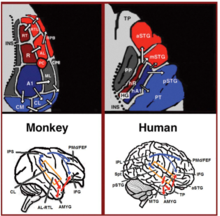
V posledních dvou desetiletích došlo k významnému pokroku v našem chápání nervového zpracování zvuků u primátů. Zpočátku zaznamenáváním nervové aktivity ve sluchových kůrách opic[18][19] a později zpracovány pomocí histologického barvení[20][21][22] a fMRI skenovací studie,[23] V primární sluchové kůře byly identifikovány 3 sluchová pole a bylo prokázáno, že je obklopuje 9 asociativních sluchových polí (obrázek 1 vlevo nahoře). Anatomické sledování a studie lézí dále naznačují oddělení mezi předním a zadním sluchovým polem, přičemž přední primární sluchová pole (oblasti R-RT) vyčnívají do předních asociativních sluchových polí (oblasti AL-RTL) a zadní primární sluchové pole (oblast A1) promítající do zadních asociativních sluchových polí (oblasti CL-CM).[20][24][25][26] Nedávno se nashromáždily důkazy, které naznačují homologii mezi sluchovými poli člověka a opice. U lidí odhalily studie histologického barvení dvě samostatná sluchová pole v primární sluchové oblasti Heschlův gyrus,[27][28] a mapováním tonotopické organizace lidských primárních sluchových polí s fMRI s vysokým rozlišením a jejím porovnáním s tonotopickou organizací primárních sluchových polí opice byla vytvořena homologie mezi předním primárním sluchovým polem člověka a oblastí opic R (u lidí označována jako oblast hR) a zadní lidské sluchové pole zadní a oblast opice A1 (u lidí označena jako oblast hA1).[29][30][31][32][33] Intrakortikální záznamy od člověka sluchová kůra dále demonstroval podobné vzorce propojení se sluchovou kůrou opice. Záznam z povrchu sluchové kůry (nadčasová rovina) uvádí, že přední Heschlův gyrus (oblast hR) vyčnívá primárně do střední přední superior temporální gyrus (mSTG-aSTG) a zadní Heschlův gyrus (oblast hA1) vyčnívá primárně k zadnímu hornímu temporálnímu gyrusu (pSTG) a planum temporale (oblast PT; obrázek 1 vpravo nahoře).[34][35] V souladu s připojením z oblasti hR k aSTG a hA1 k pSTG je studie fMRI u pacienta se zhoršeným rozpoznáváním zvuku (sluchová agnosie ), u kterého byla prokázána snížená bilaterální aktivace v oblastech hR a aSTG, ale se ušetřenou aktivací v mSTG-pSTG.[36] Tento vzor konektivity je také potvrzen studií, která zaznamenávala aktivaci z postranního povrchu sluchové kůry a hlásila simultánní nepřekrývající se aktivační klastry v pSTG a mSTG-aSTG při poslechu zvuků.[37]
Po proudu do sluchové kůry anatomické sledovací studie u opic vymezily projekce od předních asociativních sluchových polí (oblasti AL-RTL) po ventrální prefrontální a premotorické kůry v dolní čelní gyrus (IFG)[38][39] a amygdala.[40] Studie kortikálního záznamu a funkčního zobrazování u opic makaků byly dále rozpracovány na tomto proudu zpracování tím, že ukázaly, že akustické informace proudí z přední sluchové kůry k temporálnímu pólu (TP) a poté k IFG.[41][42][43][44][45][46] Tato cesta se běžně označuje jako sluchový ventrální proud (AVS; obrázek 1, vlevo dole červené šipky). Na rozdíl od předních sluchových polí studie trasování uváděly, že zadní sluchová pole (oblasti CL-CM) se promítají primárně do dorsolaterálních prefrontálních a premotorických kortik (ačkoli některé projekce v IFG končí).[47][39] Kortikální záznamy a anatomické sledovací studie u opic dále poskytly důkazy o tom, že tento zpracovatelský proud proudí ze zadních sluchových polí do čelního laloku přes reléovou stanici v nitro-parietálním sulku (IPS).[48][49][50][51][52][53] Tato cesta se běžně označuje jako sluchový hřbetní proud (ADS; obrázek 1, vlevo dole modré šipky). Porovnání drah bílé hmoty zapojených do komunikace u lidí a opic s difúzní tenzorové zobrazování techniky naznačuje podobné spojení AVS a ADS u těchto dvou druhů (opice,[52] Člověk[54][55][56][57][58][59]). U lidí se ukázalo, že pSTG promítá do temenního laloku (sylvian temenně-temporální spojení -nižší temenní lalůček; Spt-IPL ), a odtud do dorsolaterálních prefrontálních a premotorických kortikal (obrázek 1, vpravo dole modré šipky) a aSTG bylo prokázáno, že vyčnívají do předního temporálního laloku (střední temporální gyrus-temporální pól; MTG-TP) a odtud do IFG (obrázek 1 vpravo dole červené šipky).
Sluchový ventrální proud
Sluchový ventrální proud (AVS) spojuje sluchová kůra s střední temporální gyrus a temporální pól, což se zase spojuje s dolní čelní gyrus. Tato cesta je zodpovědná za rozpoznávání zvuku a je proto známá jako sluchová cesta „co“. Mezi funkce AVS patří následující.
Rozpoznávání zvuku
Akumulativní konvergující důkazy naznačují, že AVS je zapojen do rozpoznávání sluchových objektů. Na úrovni primární sluchové kůry ukázaly nahrávky opic vyšší procento neuronů selektivních pro naučené melodické sekvence v oblasti R než v oblasti A1,[60] a studie na lidech prokázala větší selektivitu pro slyšené slabiky v předním Heschlově gyrusu (oblast hR) než zadní Heschlovy gyrus (oblast hA1).[61] V následných asociativních sluchových polích studie od opic i lidí uvádějí, že hranice mezi předním a zadním sluchovým polem (PC na ploše 1 u opice a mSTG u člověka) zpracovává atributy výšky tónu, které jsou nezbytné pro rozpoznávání sluchových objektů .[18] Přední sluchová pole opic byla také prokázána se selektivitou pro kon-specifické vokalizace s intra-kortikálními záznamy.[41][19][62] a funkční zobrazování[63][42][43] Jedna studie na opicích fMRI dále prokázala roli aSTG při rozpoznávání jednotlivých hlasů.[42] Role lidského mSTG-aSTG při rozpoznávání zvuku byla demonstrována pomocí funkčních zobrazovacích studií, které korelovaly aktivitu v této oblasti s izolací sluchových objektů od hluku pozadí,[64][65] as rozpoznáváním mluvených slov,[66][67][68][69][70][71][72] hlasy,[73] melodie,[74][75] zvuky prostředí,[76][77][78] a komunikativní zvuky bez řeči.[79] A metaanalýza studií fMRI[80] dále prokázal funkční disociaci mezi levým mSTG a aSTG, přičemž první zpracovává krátké řečové jednotky (fonémy) a druhý zpracovává delší jednotky (např. slova, zvuky prostředí). Studie, která zaznamenávala nervovou aktivitu přímo z levého pSTG a aSTG, uváděla, že aSTG, ale ne pSTG, byl aktivnější, když pacient poslouchal řeč v jejím rodném jazyce než v neznámém cizím jazyce.[81] Důsledkem je, že elektrostimulace k aSTG tohoto pacienta vedla ke zhoršení vnímání řeči[81] (viz také[82][83] pro podobné výsledky). Intrakortikální záznamy z pravé a levé strany aSTG dále ukázaly, že řeč je zpracována laterálně na hudbu.[81] Studie fMRI u pacienta se zhoršeným rozpoznáváním zvuku (sluchová agnosie ) kvůli mozkový kmen poškození bylo také prokázáno se sníženou aktivací v oblastech hR a aSTG obou hemisfér při slyšení mluvených slov a zvuků prostředí.[36] Záznamy z přední sluchové kůry opic při zachování naučených zvuků v pracovní paměti,[46] a oslabující účinek indukovaných lézí na tuto oblast na vyvolání pracovní paměti,[84][85][86] dále zapojit AVS do udržování vnímaných sluchových objektů v pracovní paměti. U lidí byla oblast mSTG-aSTG také hlášena jako aktivní během zkoušky slyšených slabik pomocí MEG.[87] a fMRI[88] Druhá studie dále prokázala, že pracovní paměť v AVS slouží pro akustické vlastnosti mluvených slov a že je nezávislá na pracovní paměti v ADS, která zprostředkovává vnitřní řeč. Studie pracovní paměti u opic také naznačují, že u opic je na rozdíl od lidí AVS dominantním úložištěm pracovní paměti.[89]
U lidí, po proudu od aSTG, se předpokládá, že MTG a TP tvoří sémantický lexikon, což je úložiště dlouhodobé paměti audiovizuálních reprezentací, které jsou vzájemně propojeny na základě sémantických vztahů. (Viz také recenze od[3][4] o tomto tématu). Primárním důkazem této role MTG-TP je, že pacienti s poškozením této oblasti (např. Pacienti s sémantická demence nebo herpes simplex virus encefalitida ) jsou hlášeny[90][91] se zhoršenou schopností popisovat vizuální a sluchové objekty a tendencí k páchání sémantických chyb při pojmenovávání objektů (tj. sémantická parafázie ). Sémantické parafázie vyjádřili také afázičtí pacienti s poškozením MTG-TP vlevo[14][92] a bylo prokázáno, že se vyskytují u neafázických pacientů po elektrostimulaci do této oblasti.[93][83] nebo podkladová dráha bílé hmoty[94] Dvě metaanalýzy literatury fMRI také uváděly, že přední MTG a TP byly trvale aktivní během sémantické analýzy řeči a textu;[66][95] a intrakortikální záznamová studie korelovala neurální výboj v MTG s porozuměním srozumitelných vět.[96]
Věta s porozuměním
Kromě extrakce významu zvuků se zdá, že oblast MTG-TP AVS má roli v porozumění větám, možná spojením pojmů dohromady (např. Sloučením pojmu „modrá“ a „košile“ za účelem vytvoření pojmu „ modré triko'). Role MTG při získávání významu z vět byla prokázána ve funkčních zobrazovacích studiích, které uvádějí silnější aktivaci v přední MTG, když jsou správné věty porovnány se seznamy slov, větami v cizím nebo nesmyslném jazyce, míchanými větami, větami se sémantickým nebo syntaktickým porušení a věty podobné sekvence zvuků prostředí.[97][98][99][100][101][102][103][104] Jedna studie fMRI[105] ve kterém byli účastníci instruováni, aby si přečetli příběh, který dále koreloval aktivitu v přední MTG s množstvím sémantického a syntaktického obsahu, které každá věta obsahovala. Studie EEG[106] který kontrastoval s kortikální aktivitou při čtení vět se syntaktickým porušením a bez něj u zdravých účastníků a pacientů s poškozením MTG-TP, dospěl k závěru, že MTG-TP v obou hemisférách se účastní automatické (na základě pravidel) fáze syntaktické analýzy (složka ELAN) a že levý MTG-TP je také zapojen do pozdější kontrolované fáze syntaktické analýzy (součást P600). U pacientů s poškozením oblasti MTG-TP byly také hlášeny poruchy porozumění větám.[14][107][108] Viz recenze[109] pro více informací o tomto tématu.
Dvoustrannost
V rozporu s modelem Wernicke-Lichtheim-Geschwind, který implikuje rozpoznávání zvuku pouze v levé hemisféře, studie, které izolovaně zkoumaly vlastnosti pravé nebo levé hemisféry pomocí jednostranné hemisférické anestézie (tj. Postup WADA[110]) nebo intrakortikální záznamy z každé polokoule[96] poskytl důkaz, že rozpoznávání zvuku je zpracováván bilaterálně. Studie, která instruovala pacienty s odpojenými hemisférami (tj. split-brain pacientů), aby odpovídala mluvená slova psaným slovům prezentovaným na pravé nebo levé hemifield, hlášená slovní zásoba v pravé hemisféře, která se téměř shoduje s velikostí levé hemisféry[111] (Slovník pravé hemisféry byl ekvivalentní slovníku zdravého 11letého dítěte). Toto dvoustranné rozpoznávání zvuků je rovněž v souladu se zjištěním, že jednostranná léze sluchové kůry málokdy vede k deficitu sluchového porozumění (tj. sluchová agnosie ), zatímco druhá léze na zbývající hemisféře (která by se mohla objevit o několik let později) ano.[112][113] A konečně, jak již bylo zmíněno dříve, skenování fMRI u pacienta se sluchovou agnosií prokázalo bilaterální sníženou aktivaci v předních sluchových kůrách,[36] a bilaterální elektrostimulace do těchto oblastí v obou hemisférách vedla k narušení rozpoznávání řeči.[81]
Sluchový hřbetní proud
Sluchový hřbetní proud spojuje sluchovou kůru s temenní lalok, což se zase spojuje s dolní čelní gyrus. U lidí i subhumánních primátů je sluchový hřbetní proud zodpovědný za zdravou lokalizaci a je proto znám jako sluchová cesta „kde“. U lidí je tato cesta (zejména v levé hemisféře) také zodpovědná za produkci řeči, opakování řeči, čtení rtů a fonologickou pracovní paměť a dlouhodobou paměť.
Produkce řeči
Studie současných lidí prokázaly roli ADS v produkci řeči, zejména v hlasovém vyjádření jmen objektů. Například v sérii studií, ve kterých byla přímo stimulována subkortikální vlákna[94] interference v levém pSTG a IPL vedlo k chybám během úkolů pojmenování objektů a interference v levém IFG vedla k zadržení řeči. Magnetická interference v pSTG a IFG zdravých účastníků také způsobovala chyby řeči a zástavu řeči[114][115] Jedna studie také uvádí, že elektrická stimulace levice IPL způsobil, že pacienti věřili, že mluvili, když ne, a že stimulace IFG způsobila, že pacienti nevědomky hýbali rty.[116] Příspěvek ADS k procesu artikulace názvů objektů mohl záviset na příjmu aferentů ze sémantického lexikonu AVS, protože intrakortikální záznamová studie uváděla aktivaci v zadní MTG před aktivací ve Spt -IPL oblast, kdy pacienti pojmenovali objekty na obrázcích[117] Intrakortikální studie elektrické stimulace rovněž uváděly, že elektrická interference se zadním MTG korelovala se zhoršeným pojmenováním objektů[118][82]
Vokální mimikry
Ačkoli vnímání zvuku je primárně připisováno AVS, zdá se, že ADS je spojena s několika aspekty vnímání řeči. Například v metaanalýze studií fMRI[119] (Turkeltaub a Coslett, 2010), ve kterém sluchové vnímání fonémy byla porovnána s těsně se shodujícími zvuky a studie byly hodnoceny pro požadovanou úroveň pozornosti, autoři dospěli k závěru, že pozornost k fonémům koreluje se silnou aktivací v oblasti pSTG-pSTS. Studie intrakortikálního záznamu, ve které byli účastníci instruováni k identifikaci slabik, také korelovala slyšení každé slabiky s vlastním aktivačním vzorem v pSTG.[120] Zapojení ADS do vnímání i produkce řeči bylo dále osvětleno v několika průkopnických funkčních zobrazovacích studiích, které kontrastovaly vnímání řeči se zjevnou nebo skrytou produkcí řeči.[121][122][123] Tyto studie prokázaly, že pSTS je aktivní pouze během vnímání řeči, zatímco oblast Spt je aktivní jak při vnímání, tak při produkci řeči. Autoři dospěli k závěru, že pSTS se promítá do oblasti Spt, která převádí sluchový vstup na artikulační pohyby.[124][125] Podobné výsledky byly získány ve studii, ve které byly elektricky stimulovány spánkové a temenní laloky účastníků. Tato studie uvádí, že elektrická stimulace oblasti pSTG interferuje s porozuměním vět a že stimulace IPL interferuje se schopností vokalizovat názvy objektů.[83] Autoři také uvádějí, že stimulace v oblasti Spt a nižší IPL způsobovala interference jak při pojmenovávání objektů, tak při porozumění řeči. Role ADS v opakování řeči je také shodná s výsledky ostatních funkčních zobrazovacích studií, které lokalizovaly aktivaci během úkolů opakování řeči do oblastí ADS.[126][127][128] Studie intrakortikálního záznamu, která zaznamenávala aktivitu ve většině temporálních, parietálních a frontálních laloků, také uváděla aktivaci v pSTG, Spt, IPL a IFG, když je opakování řeči v kontrastu s vnímáním řeči.[129] Neuropsychologické studie také zjistily, že jedinci s deficitem opakování řeči, ale zachovaným sluchovým porozuměním (tj. vodivostní afázie ) trpí ohraničeným poškozením oblasti Spt-IPL[130][131][132][133][134][135][136] nebo poškození výčnělků vycházejících z této oblasti a zaměřených na čelní lalok[137][138][139][140] Studie také uvádějí přechodný stav opakování řeči deficit u pacientů po přímé intrakortikální elektrické stimulaci do stejné oblasti.[11][141][142] Vhled do účelu opakování řeči v ADS poskytují longitudinální studie dětí, které korelovaly učení cizí slovní zásoby se schopností opakovat nesmyslná slova.[143][144]
Monitorování řeči
Kromě opakování a vytváření řeči se zdá, že služba ADS hraje roli při sledování kvality výstupu řeči. Neuroanatomické důkazy naznačují, že ADS je vybaven sestupnými spojeními z IFG do pSTG, které přenášejí informace o motorické aktivitě (tj. Vedlejších výbojích) v hlasovém aparátu (ústa, jazyk, hlasové záhyby). Tato zpětná vazba označuje zvuk vnímaný během produkce řeči jako vlastní produkci a lze ji použít k úpravě hlasového aparátu ke zvýšení podobnosti mezi vnímanými a vydávanými hovory. Důkazy pro sestupné spojení z IFG na pSTG poskytla studie, která elektricky stimulovala IFG během chirurgických operací a hlásila šíření aktivace do oblasti pSTG-pSTS-Spt[145] Studie[146] srovnávající schopnost afázických pacientů s poškozením čelního, temenního nebo spánkového laloku rychle a opakovaně formulovat řetězec slabik uvádí, že poškození čelního laloku narušilo artikulaci obou identických slabikových řetězců („Bababa“) a neidentických slabikových řetězce („Badaga“), zatímco u pacientů s poškozením temporálního nebo temenního laloku došlo k poškození pouze při artikulaci neidentických slabikových řetězců. Protože pacienti s poškozením temporálního a parietálního laloku byli schopni v prvním úkolu opakovat slabičnou strunu, zdá se, že jejich vnímání a produkce řeči jsou relativně zachovány, a jejich deficit ve druhém úkolu je proto důsledkem zhoršeného sledování. Studie fMRI, která demonstrovala roli sestupných spojení ADS při monitorování vysílaných hovorů, instruovala účastníky, aby mluvili za normálních podmínek nebo při poslechu upravené verze vlastního hlasu (zpožděný první formant) a uvedli, že slyšení zkreslené verze vlastního hlasu vede ve zvýšené aktivaci v pSTG.[147] Další ukázkou toho, že ADS usnadňuje motorickou zpětnou vazbu během mimiky, je intrakortikální záznamová studie, která kontrastuje s vnímáním a opakováním řeči.[129] Autoři uvádějí, že kromě aktivace v IPL a IFG je opakování řeči charakterizováno silnější aktivací v pSTG než během vnímání řeči.
Integrace fonémů s pohyby rtů
Ačkoli vnímání zvuku je primárně připisováno AVS, zdá se, že ADS je spojena s několika aspekty vnímání řeči. Například v metaanalýze studií fMRI[119] ve kterém sluchové vnímání fonémů bylo porovnáno s blízce se shodujícími zvuky a studie byly hodnoceny na požadovanou úroveň pozornosti, autoři dospěli k závěru, že pozornost na fonémy koreluje se silnou aktivací v oblasti pSTG-pSTS. Studie intrakortikálního záznamu, ve které byli účastníci instruováni k identifikaci slabik, také korelovala slyšení každé slabiky s vlastním aktivačním vzorem v pSTG.[148] V souladu s rolí ADS při rozlišování fonémů,[119] studie připisují integraci fonémů a jejich odpovídajících pohybů rtů (tj. vizuů) pSTS z ADS. Například studie fMRI[149] koreloval aktivaci v pSTS s McGurkovou iluzí (ve které slyšení slabiky „ba“ při pohledu na viseme „ga“ vede k vnímání slabiky „da“). Další studie zjistila, že použití magnetické stimulace k narušení zpracování v této oblasti dále narušuje McGurkovu iluzi.[150] Spojení pSTS s audiovizuální integrací řeči bylo také prokázáno ve studii, která účastníkům poskytla obrázky tváří a mluvených slov různé kvality. Studie uvádí, že pSTS vybírá kombinované zvýšení jasnosti tváří a mluvených slov.[151] Potvrzující důkazy poskytla studie fMRI[152] který kontrastoval s vnímáním audiovizuální řeči s audiovizuálním nemluvním (obrázky a zvuky nástrojů). Tato studie uvádí detekci kompartmentů selektivních pro řeč v pSTS. Kromě toho studie fMRI[153] který kontrastoval s kongruentní audiovizuální řečí s nesourodou řečí (obrázky statických obličejů) uváděl aktivaci pSTS. Přehled představující další sbližující se důkazy týkající se role pSTS a ADS v integraci fonémů a vizém viz.[154]
Fonologická dlouhodobá paměť
Rostoucí množství důkazů naznačuje, že lidé mají kromě dlouhodobého úložiště slovních významů umístěných v MTG-TP AVS (tj. Sémantický lexikon) také dlouhodobý úložiště názvů objektů. nachází se v oblasti Spt-IPL ADS (tj. fonologický lexikon). Například studie[155][156] zkoumání pacientů s poškozením AVS (poškození MTG) nebo poškozením ADS (poškození IPL) uvádí, že poškození MTG vede k tomu, že jednotlivci nesprávně identifikují předměty (např. nazývají „kozu“ „ovcí“, příklad sémantická parafázie ). Naopak poškození IPL vede k tomu, že jednotlivci objekt správně identifikují, ale nesprávně vysloví jeho název (např. „Gof“ místo „goat“, příklad phonemic paraphasia ). Sémantické chyby parafázie byly také hlášeny u pacientů, kteří dostávali intrakortikální elektrickou stimulaci AVS (MTG), a fonemické chyby parafázie byly hlášeny u pacientů, jejichž ADS (pSTG, Spt a IPL) dostávali intrakortikální elektrickou stimulaci.[83][157][94] Další podporou role ADS při pojmenovávání objektů je studie MEG, která lokalizovala aktivitu v IPL během učení a během vyvolávání názvů objektů.[158] Studie, která vyvolala magnetickou interferenci v IPL účastníků, zatímco odpovídali na otázky týkající se objektu, uváděla, že účastníci byli schopni odpovědět na otázky týkající se charakteristik objektu nebo vjemových atributů, ale byli narušeni, když se zeptali, zda slovo obsahuje dvě nebo tři slabiky.[159] Studie MEG také korelovala zotavení z anomie (porucha charakterizovaná zhoršenou schopností pojmenovat objekty) se změnami v aktivaci IPL.[160] Další podporu role IPL v kódování zvuků slov představují studie uvádějící, že ve srovnání s jednojazyčnými mají dvojjazyční v IPL větší kortikální hustotu, ale ne v MTG.[161][162] Protože důkazy ukazují, že v dvojjazyční, různé fonologické reprezentace stejného slova mají stejné sémantické zastoupení,[163] toto zvýšení hustoty v IPL ověřuje existenci fonologického lexikonu: očekává se, že sémantický lexikon dvojjazyčných bude mít podobnou velikost jako sémantický lexikon jednojazyčných, zatímco jejich fonologický lexikon by měl být dvakrát větší. V souladu s tímto zjištěním koreluje kortikální hustota v IPL jednojazyčných také s velikostí slovníku.[164][165] Zejména funkční disociace AVS a ADS v úlohách pojmenování objektů je podporována kumulativními důkazy z výzkumu čtení, které ukazují, že sémantické chyby korelují s poruchou MTG a phonemic chyby s poruchou IPL. Na základě těchto asociací byla sémantická analýza textu spojena s inferior-temporal gyrus a MTG a fonologická analýza textu byla spojena s pSTG-Spt-IPL[166][167][168]
Fonologická pracovní paměť
Pracovní paměť se často považuje za dočasnou aktivaci reprezentací uložených v dlouhodobé paměti, které se používají pro řeč (fonologická reprezentace). Toto sdílení zdrojů mezi pracovní pamětí a řečí je zřejmé z nálezu[169][170] že mluvení během zkoušky má za následek výrazné snížení počtu položek, které lze z pracovní paměti vyvolat (artikulační potlačení ). O zapojení fonologického lexikonu do pracovní paměti svědčí také tendence jednotlivců dělat více chyb při vybavování slov z nedávno naučeného seznamu fonologicky podobných slov než ze seznamu fonologicky odlišných slov ( účinek fonologické podobnosti ).[169] Studie také zjistily, že řečové chyby spáchané během čtení jsou pozoruhodně podobné chybám řeči, které vznikly při vyvolání nedávno naučených, fonologicky podobných slov z pracovní paměti.[171] Bylo také pozorováno, že pacienti s poškozením IPL vykazují jak chyby produkce řeči, tak zhoršenou pracovní paměť[172][173][174][175] Nakonec je názor, že verbální pracovní paměť je výsledkem dočasné aktivace fonologických reprezentací v ADS, kompatibilní s nedávnými modely popisujícími pracovní paměť jako kombinaci udržování reprezentací v mechanismu pozornosti souběžně s dočasnou aktivací reprezentací v dlouhodobé paměti.[170][176][177][178] Bylo argumentováno, že role ADS při nácviku seznamů slov je důvodem, proč je tato cesta aktivní při porozumění větám[179] Přehled role ADS v pracovní paměti najdete v části.[180]

Vývoj jazyka
Sluchový hřbetní proud má také funkce nesouvisející s jazykem, jako je lokalizace zvuku[181][182][183][184][185] a vedení pohybů očí.[186][187] Nedávné studie také naznačují roli ADS při lokalizaci členů rodiny / kmene jako studie[188] zaznamenané z kůry epileptického pacienta uvádějí, že pSTG, ale nikoli aSTG, je selektivní pro přítomnost nových řečníků. FMRI[189] Studie plodů v jejich třetím trimestru také prokázaly, že oblast Spt je selektivnější pro ženskou řeč než čisté tóny a podsekce Spt je selektivní pro řeč jejich matky na rozdíl od neznámých ženských hlasů.
V současné době není známo, proč je tolik funkcí připisováno lidským ADS. Pokus o sjednocení těchto funkcí do jednoho rámce byl proveden v modelu evoluce jazyka „Odkud do jaké“[190][191] V souladu s tímto modelem každá funkce ADS naznačuje jinou přechodnou fázi ve vývoji jazyka. Role zvukové lokalizace a integrace zvukového umístění s hlasy a sluchovými objekty je interpretována jako důkaz toho, že původem řeči je výměna kontaktních hovorů (hovorů používaných k hlášení polohy v případě oddělení) mezi matkami a potomky. Role ADS při vnímání a produkci intonací je interpretována jako důkaz, že řeč začala úpravou kontaktních hovorů intonacemi, případně pro rozlišení poplachových kontaktních volání od bezpečných kontaktních volání. Role ADS v kódování názvů objektů (fonologická dlouhodobá paměť) je interpretována jako důkaz postupného přechodu od modifikace hovorů intonací k úplné hlasové kontrole. Role ADS v integraci pohybů rtů s fonémy a v opakování řeči je interpretována jako důkaz toho, že mluvená slova se naučili kojenci napodobující vokalizace svých rodičů, původně napodobováním jejich pohybů rtů. The role of the ADS in phonological working memory is interpreted as evidence that the words learned through mimicry remained active in the ADS even when not spoken. This resulted with individuals capable of rehearsing a list of vocalizations, which enabled the production of words with several syllables. Further developments in the ADS enabled the rehearsal of lists of words, which provided the infra-structure for communicating with sentences.
Sign language in the brain
Neuroscientific research has provided a scientific understanding of how sign language is processed in the brain. There are over 135 discrete sign languages around the world- making use of different accents formed by separate areas of a country.[192]
By resorting to lesion analyses and neuroimaging, neuroscientists have discovered that whether it be spoken or sign language, human brains process language in general, in a similar manner regarding which area of the brain is being used. [192]Lesion analyses are used to examine the consequences of damage to specific brain regions involved in language while neuroimaging explore regions that are engaged in the processing of language.[192]
Previous hypotheses have been made that damage to Broca's area or Wernicke’s area does not affect sign language being perceived; however, it is not the case. Studies have shown that damage to these areas are similar in results in spoken language where sign errors are present and/or repeated. [192]In both types of languages, they are affected by damage to the left hemisphere of the brain rather than the right -usually dealing with the arts.
There are obvious patterns for utilizing and processing language. In sign language, Broca’s area is activated while processing sign language employs Wernicke’s area similar to that of spoken language [192]
There have been other hypotheses about the lateralization of the two hemispheres. Specifically, the right hemisphere was thought to contribute to the overall communication of a language globally whereas the left hemisphere would be dominant in generating the language locally.[193] Through research in aphasias, RHD signers were found to have a problem maintaining the spatial portion of their signs, confusing similar signs at different locations necessary to communicate with another properly.[193] LHD signers, on the other hand, had similar results to those of hearing patients. Furthermore, other studies have emphasized that sign language is present bilaterally but will need to continue researching to reach a conclusion.[193]
Writing in the brain
There is a comparatively small body of research on the neurology of reading and writing.[194] Most of the studies performed deal with reading rather than writing or spelling, and the majority of both kinds focus solely on the English language.[195] Anglický pravopis is less transparent than that of other languages using a Latinské písmo.[194] Another difficulty is that some studies focus on spelling words of English and omit the few logographic characters found in the script.[194]
In terms of spelling, English words can be divided into three categories – regular, irregular, and “novel words” or “nonwords.” Regular words are those in which there is a regular, one-to-one correspondence between grafém a foném in spelling. Irregular words are those in which no such correspondence exists. Nonwords are those that exhibit the expected orthography of regular words but do not carry meaning, such as nonce words a onomatopoeia.[194]
An issue in the cognitive and neurological study of reading and spelling in English is whether a single-route or dual-route model best describes how literate speakers are able to read and write all three categories of English words according to accepted standards of orthographic correctness. Single-route models posit that lexical memory is used to store all spellings of words for retrieval in a single process. Dual-route models posit that lexical memory is employed to process irregular and high-frequency regular words, while low-frequency regular words and nonwords are processed using a sub-lexical set of phonological rules.[194]
The single-route model for reading has found support in computer modelling studies, which suggest that readers identify words by their orthographic similarities to phonologically alike words.[194] However, cognitive and lesion studies lean towards the dual-route model. Cognitive spelling studies on children and adults suggest that spellers employ phonological rules in spelling regular words and nonwords, while lexical memory is accessed to spell irregular words and high-frequency words of all types.[194] Similarly, lesion studies indicate that lexical memory is used to store irregular words and certain regular words, while phonological rules are used to spell nonwords.[194]
More recently, neuroimaging studies using pozitronová emisní tomografie a fMRI have suggested a balanced model in which the reading of all word types begins in the visual word form area, but subsequently branches off into different routes depending upon whether or not access to lexical memory or semantic information is needed (which would be expected with irregular words under a dual-route model).[194] A 2007 fMRI study found that subjects asked to produce regular words in a spelling task exhibited greater activation in the left posterior STG, an area used for phonological processing, while the spelling of irregular words produced greater activation of areas used for lexical memory and semantic processing, such as the left IFG a vlevo SMG and both hemispheres of the MTG.[194] Spelling nonwords was found to access members of both pathways, such as the left STG and bilateral MTG and ITG.[194] Significantly, it was found that spelling induces activation in areas such as the left fusiform gyrus and left SMG that are also important in reading, suggesting that a similar pathway is used for both reading and writing.[194]
Far less information exists on the cognition and neurology of non-alphabetic and non-English scripts. Every language has a morfologické a a fonologický component, either of which can be recorded by a systém psaní. Scripts recording words and morphemes are considered logografické, while those recording phonological segments, such as syllabaries a abecedy, are phonographic.[195] Most systems combine the two and have both logographic and phonographic characters.[195]
In terms of complexity, writing systems can be characterized as “transparent” or “opaque” and as “shallow” or “deep.” A “transparent” system exhibits an obvious correspondence between grapheme and sound, while in an “opaque” system this relationship is less obvious. The terms “shallow” and “deep” refer to the extent that a system’s orthography represents morphemes as opposed to phonological segments.[195] Systems that record larger morphosyntactic or phonological segments, such as logographic systems and syllabaries put greater demand on the memory of users.[195] It would thus be expected that an opaque or deep writing system would put greater demand on areas of the brain used for lexical memory than would a system with transparent or shallow orthography.
Viz také
Reference
- ^ Seidenberg MS, Petitto LA (1987). "Communication, symbolic communication, and language: Comment on Savage-Rumbaugh, McDonald, Sevcik, Hopkins, and Rupert (1986)". Journal of Experimental Psychology: General. 116 (3): 279–287. doi:10.1037/0096-3445.116.3.279.
- ^ A b Geschwind N (June 1965). "Disconnexion syndromes in animals and man. I". Posouzení. Mozek. 88 (2): 237–94. doi:10.1093/brain/88.2.237. PMID 5318481.
- ^ A b Hickok G, Poeppel D (May 2007). "The cortical organization of speech processing". Posouzení. Recenze přírody. Neurovědy. 8 (5): 393–402. doi:10.1038/nrn2113. PMID 17431404. S2CID 6199399.
- ^ A b Gow DW (June 2012). "The cortical organization of lexical knowledge: a dual lexicon model of spoken language processing". Posouzení. Mozek a jazyk. 121 (3): 273–88. doi:10.1016/j.bandl.2012.03.005. PMC 3348354. PMID 22498237.
- ^ Poliva O (2017-09-20). "From where to what: a neuroanatomically based evolutionary model of the emergence of speech in humans". Posouzení. F1000Výzkum. 4: 67. doi:10.12688/f1000research.6175.3. PMC 5600004. PMID 28928931.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ Poliva O (2016). "From Mimicry to Language: A Neuroanatomically Based Evolutionary Model of the Emergence of Vocal Language". Posouzení. Frontiers in Neuroscience. 10: 307. doi:10.3389/fnins.2016.00307. PMC 4928493. PMID 27445676. Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
- ^ Pickles JO (2015). "Chapter 1: Auditory pathways: anatomy and physiology". In Aminoff MJ, Boller F, Swaab DF (eds.). Příručka klinické neurologie. Posouzení. 129. s. 3–25. doi:10.1016/B978-0-444-62630-1.00001-9. ISBN 978-0-444-62630-1. PMID 25726260.
- ^ Lichteim L (1885-01-01). "On Aphasia". Mozek. 7 (4): 433–484. doi:10.1093/brain/7.4.433.
- ^ Wernicke C (1974). Der aphasische Symptomenkomplex. Springer Berlin Heidelberg. s. 1–70. ISBN 978-3-540-06905-8.
- ^ Aboitiz F, García VR (December 1997). "The evolutionary origin of the language areas in the human brain. A neuroanatomical perspective". Výzkum mozku. Recenze výzkumu mozku. 25 (3): 381–96. doi:10.1016/s0165-0173(97)00053-2. PMID 9495565. S2CID 20704891.
- ^ A b Anderson JM, Gilmore R, Roper S, Crosson B, Bauer RM, Nadeau S, Beversdorf DQ, Cibula J, Rogish M, Kortencamp S, Hughes JD, Gonzalez Rothi LJ, Heilman KM (October 1999). "Conduction aphasia and the arcuate fasciculus: A reexamination of the Wernicke-Geschwind model". Mozek a jazyk. 70 (1): 1–12. doi:10.1006/brln.1999.2135. PMID 10534369. S2CID 12171982.
- ^ DeWitt I, Rauschecker JP (November 2013). "Wernicke's area revisited: parallel streams and word processing". Mozek a jazyk. 127 (2): 181–91. doi:10.1016/j.bandl.2013.09.014. PMC 4098851. PMID 24404576.
- ^ Dronkers NF (January 2000). "The pursuit of brain-language relationships". Mozek a jazyk. 71 (1): 59–61. doi:10.1006/brln.1999.2212. PMID 10716807. S2CID 7224731.
- ^ A b C Dronkers NF, Wilkins DP, Van Valin RD, Redfern BB, Jaeger JJ (May 2004). "Lesion analysis of the brain areas involved in language comprehension". Poznání. 92 (1–2): 145–77. doi:10.1016/j.cognition.2003.11.002. hdl:11858/00-001M-0000-0012-6912-A. PMID 15037129. S2CID 10919645.
- ^ Mesulam MM, Thompson CK, Weintraub S, Rogalski EJ (August 2015). "The Wernicke conundrum and the anatomy of language comprehension in primary progressive aphasia". Mozek. 138 (Pt 8): 2423–37. doi:10.1093/brain/awv154. PMC 4805066. PMID 26112340.
- ^ Poeppel D, Emmorey K, Hickok G, Pylkkänen L (October 2012). „Směrem k nové neurobiologii jazyka“. The Journal of Neuroscience. 32 (41): 14125–31. doi:10.1523/jneurosci.3244-12.2012. PMC 3495005. PMID 23055482.
- ^ Vignolo LA, Boccardi E, Caverni L (March 1986). "Unexpected CT-scan findings in global aphasia". Kůra; Časopis věnovaný studiu nervového systému a chování. 22 (1): 55–69. doi:10.1016/s0010-9452(86)80032-6. PMID 2423296. S2CID 4479679.
- ^ A b Bendor D, Wang X (August 2006). "Cortical representations of pitch in monkeys and humans". Aktuální názor v neurobiologii. 16 (4): 391–9. doi:10.1016/j.conb.2006.07.001. PMC 4325365. PMID 16842992.
- ^ A b Rauschecker JP, Tian B, Hauser M (April 1995). "Processing of complex sounds in the macaque nonprimary auditory cortex". Věda. 268 (5207): 111–4. doi:10.1126/science.7701330. PMID 7701330.
- ^ A b de la Mothe LA, Blumell S, Kajikawa Y, Hackett TA (May 2006). "Cortical connections of the auditory cortex in marmoset monkeys: core and medial belt regions". The Journal of Comparative Neurology. 496 (1): 27–71. doi:10.1002/cne.20923. PMID 16528722.
- ^ de la Mothe LA, Blumell S, Kajikawa Y, Hackett TA (May 2012). "Cortical connections of auditory cortex in marmoset monkeys: lateral belt and parabelt regions". Anatomický záznam. 295 (5): 800–21. doi:10.1002/ar.22451. PMC 3379817. PMID 22461313.
- ^ Kaas JH, Hackett TA (October 2000). "Subdivisions of auditory cortex and processing streams in primates". Sborník Národní akademie věd Spojených států amerických. 97 (22): 11793–9. doi:10.1073/pnas.97.22.11793. PMC 34351. PMID 11050211.
- ^ Petkov CI, Kayser C, Augath M, Logothetis NK (July 2006). "Functional imaging reveals numerous fields in the monkey auditory cortex". PLOS Biology. 4 (7): e215. doi:10.1371/journal.pbio.0040215. PMC 1479693. PMID 16774452.
- ^ Morel A, Garraghty PE, Kaas JH (September 1993). "Tonotopic organization, architectonic fields, and connections of auditory cortex in macaque monkeys". The Journal of Comparative Neurology. 335 (3): 437–59. doi:10.1002/cne.903350312. PMID 7693772.
- ^ Rauschecker JP, Tian B (October 2000). "Mechanisms and streams for processing of "what" and "where" in auditory cortex". Sborník Národní akademie věd Spojených států amerických. 97 (22): 11800–6. doi:10.1073/pnas.97.22.11800. PMC 34352. PMID 11050212.
- ^ Rauschecker JP, Tian B, Pons T, Mishkin M (May 1997). "Serial and parallel processing in rhesus monkey auditory cortex". The Journal of Comparative Neurology. 382 (1): 89–103. doi:10.1002/(sici)1096-9861(19970526)382:1<89::aid-cne6>3.3.co;2-y. PMID 9136813.
- ^ Sweet RA, Dorph-Petersen KA, Lewis DA (October 2005). "Mapping auditory core, lateral belt, and parabelt cortices in the human superior temporal gyrus". The Journal of Comparative Neurology. 491 (3): 270–89. doi:10.1002/cne.20702. PMID 16134138.
- ^ Wallace MN, Johnston PW, Palmer AR (April 2002). "Histochemical identification of cortical areas in the auditory region of the human brain". Experimentální výzkum mozku. 143 (4): 499–508. doi:10.1007/s00221-002-1014-z. PMID 11914796. S2CID 24211906.
- ^ Da Costa S, van der Zwaag W, Marques JP, Frackowiak RS, Clarke S, Saenz M (October 2011). "Human primary auditory cortex follows the shape of Heschl's gyrus". The Journal of Neuroscience. 31 (40): 14067–75. doi:10.1523/jneurosci.2000-11.2011. PMC 6623669. PMID 21976491.
- ^ Humphries C, Liebenthal E, Binder JR (April 2010). "Tonotopic organization of human auditory cortex". NeuroImage. 50 (3): 1202–11. doi:10.1016/j.neuroimage.2010.01.046. PMC 2830355. PMID 20096790.
- ^ Langers DR, van Dijk P (September 2012). "Mapping the tonotopic organization in human auditory cortex with minimally salient acoustic stimulation". Mozková kůra. 22 (9): 2024–38. doi:10.1093/cercor/bhr282. PMC 3412441. PMID 21980020.
- ^ Striem-Amit E, Hertz U, Amedi A (March 2011). "Extensive cochleotopic mapping of human auditory cortical fields obtained with phase-encoding FMRI". PLOS ONE. 6 (3): e17832. doi:10.1371/journal.pone.0017832. PMC 3063163. PMID 21448274.
- ^ Woods DL, Herron TJ, Cate AD, Yund EW, Stecker GC, Rinne T, Kang X (2010). "Functional properties of human auditory cortical fields". Frontiers in Systems Neuroscience. 4: 155. doi:10.3389/fnsys.2010.00155. PMC 3001989. PMID 21160558.
- ^ Gourévitch B, Le Bouquin Jeannès R, Faucon G, Liégeois-Chauvel C (March 2008). "Temporal envelope processing in the human auditory cortex: response and interconnections of auditory cortical areas" (PDF). Hearing Research. 237 (1–2): 1–18. doi:10.1016/j.heares.2007.12.003. PMID 18255243. S2CID 15271578.
- ^ Guéguin M, Le Bouquin-Jeannès R, Faucon G, Chauvel P, Liégeois-Chauvel C (February 2007). "Evidence of functional connectivity between auditory cortical areas revealed by amplitude modulation sound processing". Mozková kůra. 17 (2): 304–13. doi:10.1093/cercor/bhj148. PMC 2111045. PMID 16514106.
- ^ A b C Poliva O, Bestelmeyer PE, Hall M, Bultitude JH, Koller K, Rafal RD (September 2015). "Functional Mapping of the Human Auditory Cortex: fMRI Investigation of a Patient with Auditory Agnosia from Trauma to the Inferior Colliculus" (PDF). Cognitive and Behavioral Neurology. 28 (3): 160–80. doi:10.1097/wnn.0000000000000072. PMID 26413744. S2CID 913296.
- ^ Chang EF, Edwards E, Nagarajan SS, Fogelson N, Dalal SS, Canolty RT, Kirsch HE, Barbaro NM, Knight RT (June 2011). "Cortical spatio-temporal dynamics underlying phonological target detection in humans". Journal of Cognitive Neuroscience. 23 (6): 1437–46. doi:10.1162/jocn.2010.21466. PMC 3895406. PMID 20465359.
- ^ Muñoz M, Mishkin M, Saunders RC (September 2009). "Resection of the medial temporal lobe disconnects the rostral superior temporal gyrus from some of its projection targets in the frontal lobe and thalamus". Mozková kůra. 19 (9): 2114–30. doi:10.1093/cercor/bhn236. PMC 2722427. PMID 19150921.
- ^ A b Romanski LM, Bates JF, Goldman-Rakic PS (January 1999). "Auditory belt and parabelt projections to the prefrontal cortex in the rhesus monkey". The Journal of Comparative Neurology. 403 (2): 141–57. doi:10.1002/(sici)1096-9861(19990111)403:2<141::aid-cne1>3.0.co;2-v. PMID 9886040.
- ^ Tanaka D (June 1976). "Thalamic projections of the dorsomedial prefrontal cortex in the rhesus monkey (Macaca mulatta)". Výzkum mozku. 110 (1): 21–38. doi:10.1016/0006-8993(76)90206-7. PMID 819108. S2CID 21529048.
- ^ A b Perrodin C, Kayser C, Logothetis NK, Petkov CI (August 2011). "Voice cells in the primate temporal lobe". Aktuální biologie. 21 (16): 1408–15. doi:10.1016/j.cub.2011.07.028. PMC 3398143. PMID 21835625.
- ^ A b C Petkov CI, Kayser C, Steudel T, Whittingstall K, Augath M, Logothetis NK (March 2008). "A voice region in the monkey brain". Přírodní neurovědy. 11 (3): 367–74. doi:10.1038/nn2043. PMID 18264095. S2CID 5505773.
- ^ A b Poremba A, Malloy M, Saunders RC, Carson RE, Herscovitch P, Mishkin M (January 2004). "Species-specific calls evoke asymmetric activity in the monkey's temporal poles". Příroda. 427 (6973): 448–51. doi:10.1038/nature02268. PMID 14749833. S2CID 4402126.
- ^ Romanski LM, Averbeck BB, Diltz M (February 2005). "Neural representation of vocalizations in the primate ventrolateral prefrontal cortex". Journal of Neurophysiology. 93 (2): 734–47. doi:10.1152/jn.00675.2004. PMID 15371495.
- ^ Russ BE, Ackelson AL, Baker AE, Cohen YE (January 2008). "Coding of auditory-stimulus identity in the auditory non-spatial processing stream". Journal of Neurophysiology. 99 (1): 87–95. doi:10.1152/jn.01069.2007. PMC 4091985. PMID 18003874.
- ^ A b Tsunada J, Lee JH, Cohen YE (June 2011). "Representation of speech categories in the primate auditory cortex". Journal of Neurophysiology. 105 (6): 2634–46. doi:10.1152/jn.00037.2011. PMC 3118748. PMID 21346209.
- ^ Cusick CG, Seltzer B, Cola M, Griggs E (September 1995). "Chemoarchitectonics and corticocortical terminations within the superior temporal sulcus of the rhesus monkey: evidence for subdivisions of superior temporal polysensory cortex". The Journal of Comparative Neurology. 360 (3): 513–35. doi:10.1002/cne.903600312. PMID 8543656.
- ^ Cohen YE, Russ BE, Gifford GW, Kiringoda R, MacLean KA (December 2004). "Selectivity for the spatial and nonspatial attributes of auditory stimuli in the ventrolateral prefrontal cortex". The Journal of Neuroscience. 24 (50): 11307–16. doi:10.1523/jneurosci.3935-04.2004. PMC 6730358. PMID 15601937.
- ^ Deacon TW (February 1992). "Cortical connections of the inferior arcuate sulcus cortex in the macaque brain". Výzkum mozku. 573 (1): 8–26. doi:10.1016/0006-8993(92)90109-m. ISSN 0006-8993. PMID 1374284. S2CID 20670766.
- ^ Lewis JW, Van Essen DC (December 2000). "Corticocortical connections of visual, sensorimotor, and multimodal processing areas in the parietal lobe of the macaque monkey". The Journal of Comparative Neurology. 428 (1): 112–37. doi:10.1002/1096-9861(20001204)428:1<112::aid-cne8>3.0.co;2-9. PMID 11058227.
- ^ Roberts AC, Tomic DL, Parkinson CH, Roeling TA, Cutter DJ, Robbins TW, Everitt BJ (May 2007). "Forebrain connectivity of the prefrontal cortex in the marmoset monkey (Callithrix jacchus): an anterograde and retrograde tract-tracing study". The Journal of Comparative Neurology. 502 (1): 86–112. doi:10.1002/cne.21300. PMID 17335041.
- ^ A b Schmahmann JD, Pandya DN, Wang R, Dai G, D'Arceuil HE, de Crespigny AJ, Wedeen VJ (March 2007). "Association fibre pathways of the brain: parallel observations from diffusion spectrum imaging and autoradiography". Mozek. 130 (Pt 3): 630–53. doi:10.1093/brain/awl359. PMID 17293361.
- ^ Seltzer B, Pandya DN (July 1984). "Further observations on parieto-temporal connections in the rhesus monkey". Experimentální výzkum mozku. 55 (2): 301–12. doi:10.1007/bf00237280. PMID 6745368. S2CID 20167953.
- ^ Catani M, Jones DK, ffytche DH (January 2005). "Perisylvian language networks of the human brain". Annals of Neurology. 57 (1): 8–16. doi:10.1002/ana.20319. PMID 15597383.
- ^ Frey S, Campbell JS, Pike GB, Petrides M (November 2008). "Dissociating the human language pathways with high angular resolution diffusion fiber tractography". The Journal of Neuroscience. 28 (45): 11435–44. doi:10.1523/jneurosci.2388-08.2008. PMC 6671318. PMID 18987180.
- ^ Makris N, Papadimitriou GM, Kaiser JR, Sorg S, Kennedy DN, Pandya DN (April 2009). "Delineation of the middle longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study". Mozková kůra. 19 (4): 777–85. doi:10.1093/cercor/bhn124. PMC 2651473. PMID 18669591.
- ^ Menjot de Champfleur N, Lima Maldonado I, Moritz-Gasser S, Machi P, Le Bars E, Bonafé A, Duffau H (January 2013). "Middle longitudinal fasciculus delineation within language pathways: a diffusion tensor imaging study in human". European Journal of Radiology. 82 (1): 151–7. doi:10.1016/j.ejrad.2012.05.034. PMID 23084876.
- ^ Turken AU, Dronkers NF (2011). "The neural architecture of the language comprehension network: converging evidence from lesion and connectivity analyses". Frontiers in Systems Neuroscience. 5: 1. doi:10.3389/fnsys.2011.00001. PMC 3039157. PMID 21347218.
- ^ Saur D, Kreher BW, Schnell S, Kümmerer D, Kellmeyer P, Vry MS, Umarova R, Musso M, Glauche V, Abel S, Huber W, Rijntjes M, Hennig J, Weiller C (November 2008). "Ventral and dorsal pathways for language". Sborník Národní akademie věd Spojených států amerických. 105 (46): 18035–40. doi:10.1073/pnas.0805234105. PMC 2584675. PMID 19004769.
- ^ Yin P, Mishkin M, Sutter M, Fritz JB (December 2008). "Early stages of melody processing: stimulus-sequence and task-dependent neuronal activity in monkey auditory cortical fields A1 and R". Journal of Neurophysiology. 100 (6): 3009–29. doi:10.1152/jn.00828.2007. PMC 2604844. PMID 18842950.
- ^ Steinschneider M, Volkov IO, Fishman YI, Oya H, Arezzo JC, Howard MA (February 2005). "Intracortical responses in human and monkey primary auditory cortex support a temporal processing mechanism for encoding of the voice onset time phonetic parameter". Mozková kůra. 15 (2): 170–86. doi:10.1093/cercor/bhh120. PMID 15238437.
- ^ Russ BE, Ackelson AL, Baker AE, Cohen YE (January 2008). "Coding of auditory-stimulus identity in the auditory non-spatial processing stream". Journal of Neurophysiology. 99 (1): 87–95. doi:10.1152/jn.01069.2007. PMC 4091985. PMID 18003874.
- ^ Joly O, Pallier C, Ramus F, Pressnitzer D, Vanduffel W, Orban GA (September 2012). "Processing of vocalizations in humans and monkeys: a comparative fMRI study" (PDF). NeuroImage. 62 (3): 1376–89. doi:10.1016/j.neuroimage.2012.05.070. PMID 22659478. S2CID 9441377.
- ^ Scheich H, Baumgart F, Gaschler-Markefski B, Tegeler C, Tempelmann C, Heinze HJ, Schindler F, Stiller D (February 1998). "Functional magnetic resonance imaging of a human auditory cortex area involved in foreground-background decomposition". Evropský žurnál neurovědy. 10 (2): 803–9. doi:10.1046/j.1460-9568.1998.00086.x. PMID 9749748.
- ^ Zatorre RJ, Bouffard M, Belin P (April 2004). "Sensitivity to auditory object features in human temporal neocortex". The Journal of Neuroscience. 24 (14): 3637–42. doi:10.1523/jneurosci.5458-03.2004. PMC 6729744. PMID 15071112.
- ^ A b Binder JR, Desai RH, Graves WW, Conant LL (December 2009). "Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies". Mozková kůra. 19 (12): 2767–96. doi:10.1093/cercor/bhp055. PMC 2774390. PMID 19329570.
- ^ Davis MH, Johnsrude IS (April 2003). "Hierarchical processing in spoken language comprehension". The Journal of Neuroscience. 23 (8): 3423–31. doi:10.1523/jneurosci.23-08-03423.2003. PMC 6742313. PMID 12716950.
- ^ Liebenthal E, Binder JR, Spitzer SM, Possing ET, Medler DA (October 2005). "Neural substrates of phonemic perception". Mozková kůra. 15 (10): 1621–31. doi:10.1093/cercor/bhi040. PMID 15703256.
- ^ Narain C, Scott SK, Wise RJ, Rosen S, Leff A, Iversen SD, Matthews PM (December 2003). "Defining a left-lateralized response specific to intelligible speech using fMRI". Mozková kůra. 13 (12): 1362–8. doi:10.1093/cercor/bhg083. PMID 14615301.
- ^ Obleser J, Boecker H, Drzezga A, Haslinger B, Hennenlotter A, Roettinger M, Eulitz C, Rauschecker JP (July 2006). "Vowel sound extraction in anterior superior temporal cortex". Mapování lidského mozku. 27 (7): 562–71. doi:10.1002/hbm.20201. PMC 6871493. PMID 16281283.
- ^ Obleser J, Zimmermann J, Van Meter J, Rauschecker JP (October 2007). "Multiple stages of auditory speech perception reflected in event-related FMRI". Mozková kůra. 17 (10): 2251–7. doi:10.1093/cercor/bhl133. PMID 17150986.
- ^ Scott SK, Blank CC, Rosen S, Wise RJ (December 2000). "Identification of a pathway for intelligible speech in the left temporal lobe". Mozek. 123 (12): 2400–6. doi:10.1093/brain/123.12.2400. PMC 5630088. PMID 11099443.
- ^ Belin P, Zatorre RJ (November 2003). "Adaptation to speaker's voice in right anterior temporal lobe". NeuroReport. 14 (16): 2105–2109. doi:10.1097/00001756-200311140-00019. PMID 14600506. S2CID 34183900.
- ^ Benson RR, Whalen DH, Richardson M, Swainson B, Clark VP, Lai S, Liberman AM (September 2001). "Parametrically dissociating speech and nonspeech perception in the brain using fMRI". Mozek a jazyk. 78 (3): 364–96. doi:10.1006/brln.2001.2484. PMID 11703063. S2CID 15328590.
- ^ Leaver AM, Rauschecker JP (June 2010). "Cortical representation of natural complex sounds: effects of acoustic features and auditory object category". The Journal of Neuroscience. 30 (22): 7604–12. doi:10.1523/jneurosci.0296-10.2010. PMC 2930617. PMID 20519535.
- ^ Lewis JW, Phinney RE, Brefczynski-Lewis JA, DeYoe EA (August 2006). "Lefties get it "right" when hearing tool sounds". Journal of Cognitive Neuroscience. 18 (8): 1314–30. doi:10.1162/jocn.2006.18.8.1314. PMID 16859417. S2CID 14049095.
- ^ Maeder PP, Meuli RA, Adriani M, Bellmann A, Fornari E, Thiran JP, Pittet A, Clarke S (October 2001). "Distinct pathways involved in sound recognition and localization: a human fMRI study" (PDF). NeuroImage. 14 (4): 802–16. doi:10.1006/nimg.2001.0888. PMID 11554799. S2CID 1388647.
- ^ Viceic D, Fornari E, Thiran JP, Maeder PP, Meuli R, Adriani M, Clarke S (November 2006). "Human auditory belt areas specialized in sound recognition: a functional magnetic resonance imaging study" (PDF). NeuroReport. 17 (16): 1659–62. doi:10.1097/01.wnr.0000239962.75943.dd. PMID 17047449. S2CID 14482187.
- ^ Shultz S, Vouloumanos A, Pelphrey K (May 2012). "The superior temporal sulcus differentiates communicative and noncommunicative auditory signals". Journal of Cognitive Neuroscience. 24 (5): 1224–32. doi:10.1162/jocn_a_00208. PMID 22360624. S2CID 10784270.
- ^ DeWitt I, Rauschecker JP (February 2012). "Phoneme and word recognition in the auditory ventral stream". Sborník Národní akademie věd Spojených států amerických. 109 (8): E505-14. doi:10.1073/pnas.1113427109. PMC 3286918. PMID 22308358.
- ^ A b C d Lachaux JP, Jerbi K, Bertrand O, Minotti L, Hoffmann D, Schoendorff B, Kahane P (October 2007). "A blueprint for real-time functional mapping via human intracranial recordings". PLOS ONE. 2 (10): e1094. doi:10.1371/journal.pone.0001094. PMC 2040217. PMID 17971857.
- ^ A b Matsumoto R, Imamura H, Inouchi M, Nakagawa T, Yokoyama Y, Matsuhashi M, Mikuni N, Miyamoto S, Fukuyama H, Takahashi R, Ikeda A (April 2011). "Left anterior temporal cortex actively engages in speech perception: A direct cortical stimulation study". Neuropsychologie. 49 (5): 1350–1354. doi:10.1016/j.neuropsychologia.2011.01.023. hdl:2433/141342. PMID 21251921. S2CID 1831334.
- ^ A b C d Roux FE, Miskin K, Durand JB, Sacko O, Réhault E, Tanova R, Démonet JF (October 2015). "Electrostimulation mapping of comprehension of auditory and visual words". Kůra; Časopis věnovaný studiu nervového systému a chování. 71: 398–408. doi:10.1016/j.cortex.2015.07.001. PMID 26332785. S2CID 39964328.
- ^ Fritz J, Mishkin M, Saunders RC (June 2005). "In search of an auditory engram". Sborník Národní akademie věd Spojených států amerických. 102 (26): 9359–64. doi:10.1073/pnas.0503998102. PMC 1166637. PMID 15967995.
- ^ Stepien LS, Cordeau JP, Rasmussen T (1960). "The effect of temporal lobe and hippocampal lesions on auditory and visual recent memory in monkeys". Mozek. 83 (3): 470–489. doi:10.1093/brain/83.3.470. ISSN 0006-8950.
- ^ Strominger NL, Oesterreich RE, Neff WD (June 1980). "Sequential auditory and visual discriminations after temporal lobe ablation in monkeys". Fyziologie a chování. 24 (6): 1149–56. doi:10.1016/0031-9384(80)90062-1. PMID 6774349. S2CID 7494152.
- ^ Kaiser J, Ripper B, Birbaumer N, Lutzenberger W (October 2003). "Dynamics of gamma-band activity in human magnetoencephalogram during auditory pattern working memory". NeuroImage. 20 (2): 816–27. doi:10.1016/s1053-8119(03)00350-1. PMID 14568454. S2CID 19373941.
- ^ Buchsbaum BR, Olsen RK, Koch P, Berman KF (November 2005). "Human dorsal and ventral auditory streams subserve rehearsal-based and echoic processes during verbal working memory". Neuron. 48 (4): 687–97. doi:10.1016/j.neuron.2005.09.029. PMID 16301183. S2CID 13202604.
- ^ Scott BH, Mishkin M, Yin P (July 2012). "Monkeys have a limited form of short-term memory in audition". Sborník Národní akademie věd Spojených států amerických. 109 (30): 12237–41. doi:10.1073/pnas.1209685109. PMC 3409773. PMID 22778411.
- ^ Noppeney U, Patterson K, Tyler LK, Moss H, Stamatakis EA, Bright P, Mummery C, Price CJ (April 2007). "Temporal lobe lesions and semantic impairment: a comparison of herpes simplex virus encephalitis and semantic dementia". Mozek. 130 (Pt 4): 1138–47. doi:10.1093/brain/awl344. PMID 17251241.
- ^ Patterson K, Nestor PJ, Rogers TT (December 2007). "Where do you know what you know? The representation of semantic knowledge in the human brain". Recenze přírody. Neurovědy. 8 (12): 976–87. doi:10.1038/nrn2277. PMID 18026167. S2CID 7310189.
- ^ Schwartz MF, Kimberg DY, Walker GM, Faseyitan O, Brecher A, Dell GS, Coslett HB (December 2009). "Anterior temporal involvement in semantic word retrieval: voxel-based lesion-symptom mapping evidence from aphasia". Mozek. 132 (Pt 12): 3411–27. doi:10.1093/brain/awp284. PMC 2792374. PMID 19942676.
- ^ Hamberger MJ, McClelland S, McKhann GM, Williams AC, Goodman RR (March 2007). "Distribution of auditory and visual naming sites in nonlesional temporal lobe epilepsy patients and patients with space-occupying temporal lobe lesions". Epilepsie. 48 (3): 531–8. doi:10.1111/j.1528-1167.2006.00955.x. PMID 17326797.
- ^ A b C Duffau H (March 2008). "The anatomo-functional connectivity of language revisited. New insights provided by electrostimulation and tractography". Neuropsychologie. 46 (4): 927–34. doi:10.1016/j.neuropsychologia.2007.10.025. PMID 18093622. S2CID 40514753.
- ^ Vigneau M, Beaucousin V, Hervé PY, Duffau H, Crivello F, Houdé O, Mazoyer B, Tzourio-Mazoyer N (May 2006). "Meta-analyzing left hemisphere language areas: phonology, semantics, and sentence processing". NeuroImage. 30 (4): 1414–32. doi:10.1016/j.neuroimage.2005.11.002. PMID 16413796. S2CID 8870165.
- ^ A b Creutzfeldt O, Ojemann G, Lettich E (October 1989). "Neuronal activity in the human lateral temporal lobe. I. Responses to speech". Experimentální výzkum mozku. 77 (3): 451–75. doi:10.1007/bf00249600. PMID 2806441. S2CID 19952034.
- ^ Mazoyer BM, Tzourio N, Frak V, Syrota A, Murayama N, Levrier O, Salamon G, Dehaene S, Cohen L, Mehler J (October 1993). "The cortical representation of speech" (PDF). Journal of Cognitive Neuroscience. 5 (4): 467–79. doi:10.1162/jocn.1993.5.4.467. PMID 23964919. S2CID 22265355.
- ^ Humphries C, Love T, Swinney D, Hickok G (October 2005). "Response of anterior temporal cortex to syntactic and prosodic manipulations during sentence processing". Mapování lidského mozku. 26 (2): 128–38. doi:10.1002/hbm.20148. PMC 6871757. PMID 15895428.
- ^ Humphries C, Willard K, Buchsbaum B, Hickok G (June 2001). "Role of anterior temporal cortex in auditory sentence comprehension: an fMRI study". NeuroReport. 12 (8): 1749–52. doi:10.1097/00001756-200106130-00046. PMID 11409752. S2CID 13039857.
- ^ Vandenberghe R, Nobre AC, Price CJ (May 2002). "The response of left temporal cortex to sentences". Journal of Cognitive Neuroscience. 14 (4): 550–60. doi:10.1162/08989290260045800. PMID 12126497. S2CID 21607482.
- ^ Friederici AD, Rüschemeyer SA, Hahne A, Fiebach CJ (February 2003). "The role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes". Mozková kůra. 13 (2): 170–7. doi:10.1093/cercor/13.2.170. PMID 12507948.
- ^ Xu J, Kemeny S, Park G, Frattali C, Braun A (2005). "Language in context: emergent features of word, sentence, and narrative comprehension". NeuroImage. 25 (3): 1002–15. doi:10.1016/j.neuroimage.2004.12.013. PMID 15809000. S2CID 25570583.
- ^ Rogalsky C, Hickok G (April 2009). "Selective attention to semantic and syntactic features modulates sentence processing networks in anterior temporal cortex". Mozková kůra. 19 (4): 786–96. doi:10.1093/cercor/bhn126. PMC 2651476. PMID 18669589.
- ^ Pallier C, Devauchelle AD, Dehaene S (February 2011). "Cortical representation of the constituent structure of sentences". Sborník Národní akademie věd Spojených států amerických. 108 (6): 2522–7. doi:10.1073/pnas.1018711108. PMC 3038732. PMID 21224415.
- ^ Brennan J, Nir Y, Hasson U, Malach R, Heeger DJ, Pylkkänen L (February 2012). "Syntactic structure building in the anterior temporal lobe during natural story listening". Mozek a jazyk. 120 (2): 163–73. doi:10.1016/j.bandl.2010.04.002. PMC 2947556. PMID 20472279.
- ^ Kotz SA, von Cramon DY, Friederici AD (October 2003). "Differentiation of syntactic processes in the left and right anterior temporal lobe: Event-related brain potential evidence from lesion patients". Mozek a jazyk. 87 (1): 135–136. doi:10.1016/s0093-934x(03)00236-0. S2CID 54320415.
- ^ Martin RC, Shelton JR, Yaffee LS (February 1994). "Language processing and working memory: Neuropsychological evidence for separate phonological and semantic capacities". Deník paměti a jazyka. 33 (1): 83–111. doi:10.1006/jmla.1994.1005.
- ^ Magnusdottir S, Fillmore P, den Ouden DB, Hjaltason H, Rorden C, Kjartansson O, Bonilha L, Fridriksson J (October 2013). "Damage to left anterior temporal cortex predicts impairment of complex syntactic processing: a lesion-symptom mapping study". Mapování lidského mozku. 34 (10): 2715–23. doi:10.1002/hbm.22096. PMC 6869931. PMID 22522937.
- ^ Bornkessel-Schlesewsky I, Schlesewsky M, Small SL, Rauschecker JP (March 2015). "Neurobiological roots of language in primate audition: common computational properties". Trendy v kognitivních vědách. 19 (3): 142–50. doi:10.1016/j.tics.2014.12.008. PMC 4348204. PMID 25600585.
- ^ Hickok G, Okada K, Barr W, Pa J, Rogalsky C, Donnelly K, Barde L, Grant A (December 2008). "Bilateral capacity for speech sound processing in auditory comprehension: evidence from Wada procedures". Mozek a jazyk. 107 (3): 179–84. doi:10.1016/j.bandl.2008.09.006. PMC 2644214. PMID 18976806.
- ^ Zaidel E (September 1976). "Auditory Vocabulary of the Right Hemisphere Following Brain Bisection or Hemidecortication". Kůra. 12 (3): 191–211. doi:10.1016/s0010-9452(76)80001-9. ISSN 0010-9452. PMID 1000988. S2CID 4479925.
- ^ Poeppel D (October 2001). "Pure word deafness and the bilateral processing of the speech code". Kognitivní věda. 25 (5): 679–693. doi:10.1016/s0364-0213(01)00050-7.
- ^ Ulrich G (May 1978). "Interhemispheric functional relationships in auditory agnosia. An analysis of the preconditions and a conceptual model". Mozek a jazyk. 5 (3): 286–300. doi:10.1016/0093-934x(78)90027-5. PMID 656899. S2CID 33841186.
- ^ Stewart L, Walsh V, Frith U, Rothwell JC (March 2001). "TMS produces two dissociable types of speech disruption" (PDF). NeuroImage. 13 (3): 472–8. doi:10.1006/nimg.2000.0701. PMID 11170812. S2CID 10392466.
- ^ Acheson DJ, Hamidi M, Binder JR, Postle BR (June 2011). "A common neural substrate for language production and verbal working memory". Journal of Cognitive Neuroscience. 23 (6): 1358–67. doi:10.1162/jocn.2010.21519. PMC 3053417. PMID 20617889.
- ^ Desmurget M, Reilly KT, Richard N, Szathmari A, Mottolese C, Sirigu A (May 2009). "Movement intention after parietal cortex stimulation in humans". Věda. 324 (5928): 811–3. doi:10.1126/science.1169896. PMID 19423830. S2CID 6555881.
- ^ Edwards E, Nagarajan SS, Dalal SS, Canolty RT, Kirsch HE, Barbaro NM, Knight RT (March 2010). "Spatiotemporal imaging of cortical activation during verb generation and picture naming". NeuroImage. 50 (1): 291–301. doi:10.1016/j.neuroimage.2009.12.035. PMC 2957470. PMID 20026224.
- ^ Boatman D, Gordon B, Hart J, Selnes O, Miglioretti D, Lenz F (August 2000). "Transcortical sensory aphasia: revisited and revised". Mozek. 123 (8): 1634–42. doi:10.1093/brain/123.8.1634. PMID 10908193.
- ^ A b C Turkeltaub PE, Coslett HB (July 2010). "Localization of sublexical speech perception components". Mozek a jazyk. 114 (1): 1–15. doi:10.1016/j.bandl.2010.03.008. PMC 2914564. PMID 20413149.
- ^ Chang EF, Rieger JW, Johnson K, Berger MS, Barbaro NM, Knight RT (November 2010). "Categorical speech representation in human superior temporal gyrus". Přírodní neurovědy. 13 (11): 1428–32. doi:10.1038/nn.2641. PMC 2967728. PMID 20890293.
- ^ Buchsbaum BR, Hickok G, Humphries C (September 2001). "Role of left posterior superior temporal gyrus in phonological processing for speech perception and production". Kognitivní věda. 25 (5): 663–678. doi:10.1207/s15516709cog2505_2. ISSN 0364-0213.
- ^ Wise RJ, Scott SK, Blank SC, Mummery CJ, Murphy K, Warburton EA (January 2001). "Separate neural subsystems within 'Wernicke's area'". Mozek. 124 (Pt 1): 83–95. doi:10.1093/brain/124.1.83. PMID 11133789.
- ^ Hickok G, Buchsbaum B, Humphries C, Muftuler T (July 2003). "Auditory-motor interaction revealed by fMRI: speech, music, and working memory in area Spt". Journal of Cognitive Neuroscience. 15 (5): 673–82. doi:10.1162/089892903322307393. PMID 12965041.
- ^ Warren JE, Wise RJ, Warren JD (December 2005). "Sounds do-able: auditory-motor transformations and the posterior temporal plane". Trendy v neurovědách. 28 (12): 636–43. doi:10.1016/j.tins.2005.09.010. PMID 16216346. S2CID 36678139.
- ^ Hickok G, Poeppel D (May 2007). "The cortical organization of speech processing". Recenze přírody. Neurovědy. 8 (5): 393–402. doi:10.1038/nrn2113. PMID 17431404. S2CID 6199399.
- ^ Karbe H, Herholz K, Weber-Luxenburger G, Ghaemi M, Heiss WD (June 1998). "Cerebral networks and functional brain asymmetry: evidence from regional metabolic changes during word repetition". Mozek a jazyk. 63 (1): 108–21. doi:10.1006/brln.1997.1937. PMID 9642023. S2CID 31335617.
- ^ Giraud AL, Price CJ (August 2001). "The constraints functional neuroimaging places on classical models of auditory word processing". Journal of Cognitive Neuroscience. 13 (6): 754–65. doi:10.1162/08989290152541421. PMID 11564320. S2CID 13916709.
- ^ Graves WW, Grabowski TJ, Mehta S, Gupta P (September 2008). "The left posterior superior temporal gyrus participates specifically in accessing lexical phonology". Journal of Cognitive Neuroscience. 20 (9): 1698–710. doi:10.1162/jocn.2008.20113. PMC 2570618. PMID 18345989.
- ^ A b Towle VL, Yoon HA, Castelle M, Edgar JC, Biassou NM, Frim DM, Spire JP, Kohrman MH (August 2008). "ECoG gamma activity during a language task: differentiating expressive and receptive speech areas". Mozek. 131 (Pt 8): 2013–27. doi:10.1093/brain/awn147. PMC 2724904. PMID 18669510.
- ^ Selnes OA, Knopman DS, Niccum N, Rubens AB (June 1985). "The critical role of Wernicke's area in sentence repetition". Annals of Neurology. 17 (6): 549–57. doi:10.1002/ana.410170604. PMID 4026225.
- ^ Axer H, von Keyserlingk AG, Berks G, von Keyserlingk DG (March 2001). "Supra- and infrasylvian conduction aphasia". Mozek a jazyk. 76 (3): 317–31. doi:10.1006/brln.2000.2425. PMID 11247647. S2CID 25406527.
- ^ Bartha L, Benke T (April 2003). "Acute conduction aphasia: an analysis of 20 cases". Mozek a jazyk. 85 (1): 93–108. doi:10.1016/s0093-934x(02)00502-3. PMID 12681350. S2CID 18466425.
- ^ Baldo JV, Katseff S, Dronkers NF (March 2012). "Brain Regions Underlying Repetition and Auditory-Verbal Short-term Memory Deficits in Aphasia: Evidence from Voxel-based Lesion Symptom Mapping". Afaziologie. 26 (3–4): 338–354. doi:10.1080/02687038.2011.602391. PMC 4070523. PMID 24976669.
- ^ Baldo JV, Klostermann EC, Dronkers NF (May 2008). "It's either a cook or a baker: patients with conduction aphasia get the gist but lose the trace". Mozek a jazyk. 105 (2): 134–40. doi:10.1016/j.bandl.2007.12.007. PMID 18243294. S2CID 997735.
- ^ Fridriksson J, Kjartansson O, Morgan PS, Hjaltason H, Magnusdottir S, Bonilha L, Rorden C (August 2010). "Impaired speech repetition and left parietal lobe damage". The Journal of Neuroscience. 30 (33): 11057–61. doi:10.1523/jneurosci.1120-10.2010. PMC 2936270. PMID 20720112.
- ^ Buchsbaum BR, Baldo J, Okada K, Berman KF, Dronkers N, D'Esposito M, Hickok G (December 2011). "Conduction aphasia, sensory-motor integration, and phonological short-term memory - an aggregate analysis of lesion and fMRI data". Mozek a jazyk. 119 (3): 119–28. doi:10.1016/j.bandl.2010.12.001. PMC 3090694. PMID 21256582.
- ^ Yamada K, Nagakane Y, Mizuno T, Hosomi A, Nakagawa M, Nishimura T (March 2007). "MR tractography depicting damage to the arcuate fasciculus in a patient with conduction aphasia". Neurologie. 68 (10): 789. doi:10.1212/01.wnl.0000256348.65744.b2. PMID 17339591.
- ^ Breier JI, Hasan KM, Zhang W, Men D, Papanicolaou AC (March 2008). "Language dysfunction after stroke and damage to white matter tracts evaluated using diffusion tensor imaging". AJNR. American Journal of Neuroradiology. 29 (3): 483–7. doi:10.3174/ajnr.A0846. PMC 3073452. PMID 18039757.
- ^ Zhang Y, Wang C, Zhao X, Chen H, Han Z, Wang Y (September 2010). "Diffusion tensor imaging depicting damage to the arcuate fasciculus in patients with conduction aphasia: a study of the Wernicke-Geschwind model". Neurologický výzkum. 32 (7): 775–8. doi:10.1179/016164109x12478302362653. PMID 19825277. S2CID 22960870.
- ^ Jones OP, Prejawa S, Hope TM, Oberhuber M, Seghier ML, Leff AP, Green DW, Price CJ (2014). "Sensory-to-motor integration during auditory repetition: a combined fMRI and lesion study". Frontiers in Human Neuroscience. 8: 24. doi:10.3389/fnhum.2014.00024. PMC 3908611. PMID 24550807.
- ^ Quigg M, Fountain NB (March 1999). "Conduction aphasia elicited by stimulation of the left posterior superior temporal gyrus". Časopis neurologie, neurochirurgie a psychiatrie. 66 (3): 393–6. doi:10.1136/jnnp.66.3.393. PMC 1736266. PMID 10084542.
- ^ Quigg M, Geldmacher DS, Elias WJ (May 2006). "Conduction aphasia as a function of the dominant posterior perisylvian cortex. Report of two cases". Journal of Neurosurgery. 104 (5): 845–8. doi:10.3171/jns.2006.104.5.845. PMID 16703895.
- ^ Service E, Kohonen V (April 1995). "Is the relation between phonological memory and foreign language learning accounted for by vocabulary acquisition?". Aplikovaná psycholingvistika. 16 (2): 155–172. doi:10.1017/S0142716400007062.
- ^ Service E (July 1992). "Phonology, working memory, and foreign-language learning". Quarterly Journal of Experimental Psychology. A, Experimentální psychologie člověka. 45 (1): 21–50. doi:10.1080/14640749208401314. PMID 1636010. S2CID 43268252.
- ^ Matsumoto R, Nair DR, LaPresto E, Najm I, Bingaman W, Shibasaki H, Lüders HO (říjen 2004). „Funkční konektivita v systému lidského jazyka: studie kortikokortikálního evokovaného potenciálu“. Mozek. 127 (Pt 10): 2316–30. doi:10.1093 / mozek / awh246. PMID 15269116.
- ^ Kimura D, Watson N (listopad 1989). "Vztah mezi ovládáním orálního pohybu a řečí". Mozek a jazyk. 37 (4): 565–90. doi:10.1016 / 0093-934x (89) 90112-0. PMID 2479446. S2CID 39913744.
- ^ Tourville JA, Reilly KJ, Guenther FH (únor 2008). "Neurální mechanismy, které jsou základem kontroly zpětné vazby řeči". NeuroImage. 39 (3): 1429–43. doi:10.1016 / j.neuroimage.2007.09.054. PMC 3658624. PMID 18035557.
- ^ Chang EF, Rieger JW, Johnson K, Berger MS, Barbaro NM, Knight RT (listopad 2010). "Kategorická reprezentace řeči v lidském nadřazeném temporálním gyrusu". Přírodní neurovědy. 13 (11): 1428–32. doi:10.1038 / č. 2641. PMC 2967728. PMID 20890293.
- ^ Nath AR, Beauchamp MS (leden 2012). „Nervový základ pro interindividuální rozdíly v McGurkově efektu, multisenzorická řečová iluze“. NeuroImage. 59 (1): 781–7. doi:10.1016 / j.neuroimage.2011.07.024. PMC 3196040. PMID 21787869.
- ^ Beauchamp MS, Nath AR, Pasalar S (únor 2010). „Transkraniální magnetická stimulace řízená fMRI odhaluje, že horní temporální sulcus je kortikální lokus McGurkova efektu“. The Journal of Neuroscience. 30 (7): 2414–7. doi:10.1523 / JNEUROSCI.4865-09.2010. PMC 2844713. PMID 20164324.
- ^ McGettigan C, Faulkner A, Altarelli I, Obleser J, Baverstock H, Scott SK (duben 2012). „S porozuměním řeči napomáhají různé modality: behaviorální a neurální interakce“. Neuropsychologie. 50 (5): 762–76. doi:10.1016 / j.neuropsychologia.2012.01.010. PMC 4050300. PMID 22266262.
- ^ Stevenson RA, James TW (únor 2009). „Audiovizuální integrace v nadřazeném temporálním sulku člověka: Inverzní účinnost a neurální zpracování rozpoznávání řeči a objektů“. NeuroImage. 44 (3): 1210–23. doi:10.1016 / j.neuroimage.2008.09.034. PMID 18973818. S2CID 8342349.
- ^ Bernstein LE, Jiang J, Pantazis D, Lu ZL, Joshi A (říjen 2011). „Vizuální fonetické zpracování lokalizované pomocí gest řeči a nedůvěrných tváří ve videu a bodovém osvětlení“. Mapování lidského mozku. 32 (10): 1660–76. doi:10,1002 / hbm.21139. PMC 3120928. PMID 20853377.
- ^ Campbell R (březen 2008). „Zpracování audiovizuální řeči: empirické a neurální základy“. Filozofické transakce Královské společnosti v Londýně. Série B, Biologické vědy. 363 (1493): 1001–10. doi:10.1098 / rstb.2007.2155. PMC 2606792. PMID 17827105.
- ^ Schwartz MF, Faseyitan O, Kim J, Coslett HB (prosinec 2012). „Příspěvek hřbetního proudu k fonologickému vyhledávání v pojmenovávání objektů“. Mozek. 135 (Pt 12): 3799–814. doi:10.1093 / mozek / aws300. PMC 3525060. PMID 23171662.
- ^ Schwartz MF, Kimberg DY, Walker GM, Faseyitan O, Brecher A, Dell GS, Coslett HB (prosinec 2009). „Přední časové zapojení do sémantického vyhledávání slov: důkazy mapující léze a příznaky na základě voxelů z afázie“. Mozek. 132 (Pt 12): 3411–27. doi:10.1093 / mozek / awp284. PMC 2792374. PMID 19942676.
- ^ Ojemann GA (červen 1983). "Organizace mozku pro jazyk z pohledu mapování elektrické stimulace". Behaviorální a mozkové vědy. 6 (2): 189–206. doi:10.1017 / S0140525X00015491. ISSN 1469-1825.
- ^ Cornelissen K, Laine M, Renvall K, Saarinen T, Martin N, Salmelin R (červen 2004). "Učení nových jmen pro nové objekty: kortikální efekty měřené magnetoencefalografií". Mozek a jazyk. 89 (3): 617–22. doi:10.1016 / j.bandl.2003.12.007. PMID 15120553. S2CID 32224334.
- ^ Hartwigsen G, Baumgaertner A, Price CJ, Koehnke M, Ulmer S, Siebner HR (září 2010). „Fonologická rozhodnutí vyžadují levý i pravý supramarginální gyri“. Sborník Národní akademie věd Spojených států amerických. 107 (38): 16494–9. doi:10.1073 / pnas.1008121107. PMC 2944751. PMID 20807747.
- ^ Cornelissen K, Laine M, Tarkiainen A, Järvensivu T, Martin N, Salmelin R (duben 2003). „Plastickost mozku dospělých vyvolaná léčbou anomie“. Journal of Cognitive Neuroscience. 15 (3): 444–61. doi:10.1162/089892903321593153. PMID 12729495. S2CID 1597939.
- ^ Mechelli A, Crinion JT, Noppeney U, O'Doherty J, Ashburner J, Frackowiak RS, Price CJ (říjen 2004). "Neurolingvistika: strukturní plasticita v dvojjazyčném mozku". Příroda. 431 (7010): 757. doi:10.1038 / 431757a. hdl:11858 / 00-001M-0000-0013-D79B-1. PMID 15483594. S2CID 4338340.
- ^ Green DW, Crinion J, Price CJ (červenec 2007). „Zkoumání účinků vícejazyčné slovní zásoby na struktury mozku pomocí morfometrie založené na voxelech“. Dvojjazyčnost. 10 (2): 189–199. doi:10.1017 / S1366728907002933. PMC 2312335. PMID 18418473.
- ^ Willness C (2016-01-08). „Oxfordská příručka organizačního klimatu a kultury, autor: Benjamin Schneider & Karen M. Barbera (Eds.), New York, NY: Oxford University Press, 2014. ISBN 978-0-19-986071-5". Recenze knih. British Journal of Psychology. 107 (1): 201–202. doi:10.1111 / bjop.12170.
- ^ Lee H, Devlin JT, Shakeshaft C, Stewart LH, Brennan A, Glensman J, Pitcher K, Crinion J, Mechelli A, Frackowiak RS, Green DW, Price CJ (leden 2007). „Anatomické stopy po osvojení slovní zásoby v adolescentním mozku“. The Journal of Neuroscience. 27 (5): 1184–9. doi:10.1523 / JNEUROSCI.4442-06.2007. PMC 6673201. PMID 17267574.
- ^ Richardson FM, Thomas MS, Filippi R, Harth H, Price CJ (květen 2010). „Kontrastní účinky znalostí slovní zásoby na dočasnou a temenní mozkovou strukturu po celou dobu životnosti“. Journal of Cognitive Neuroscience. 22 (5): 943–54. doi:10.1162 / jocn.2009.21238. PMC 2860571. PMID 19366285.
- ^ Jobard G, Crivello F, Tzourio-Mazoyer N (říjen 2003). „Hodnocení teorie dvojí cesty čtení: metanalýza 35 neuroimagingových studií“. NeuroImage. 20 (2): 693–712. doi:10.1016 / s1053-8119 (03) 00343-4. PMID 14568445. S2CID 739665.
- ^ Bolger DJ, Perfetti CA, Schneider W (květen 2005). „Znovu navštívený mezikulturní účinek na mozek: univerzální struktury plus variace psacího systému“. Mapování lidského mozku (francouzsky). 25 (1): 92–104. doi:10.1002 / hbm.20124. PMC 6871743. PMID 15846818.
- ^ Brambati SM, Ogar J, Neuhaus J, Miller BL, Gorno-Tempini ML (červenec 2009). „Poruchy čtení u primární progresivní afázie: studie chování a neuroimagingu“. Neuropsychologie. 47 (8–9): 1893–900. doi:10.1016 / j.neuropsychologia.2009.02.033. PMC 2734967. PMID 19428421.
- ^ A b Baddeley A, Lewis V, Vallar G (květen 1984). „Zkoumání artikulační smyčky“. Sekce Čtvrtletník experimentální psychologie A. 36 (2): 233–252. doi:10.1080/14640748408402157. S2CID 144313607.
- ^ A b Cowan N (únor 2001). „Kouzelné číslo 4 v krátkodobé paměti: přehodnocení kapacity duševní paměti“. Behaviorální a mozkové vědy. 24 (1): 87–114, diskuse 114–85. doi:10.1017 / S0140525X01003922. PMID 11515286.
- ^ Caplan D, Rochon E, Waters GS (srpen 1992). "Artikulační a fonologické determinanty efektů délky slova v úlohách rozsahu". Quarterly Journal of Experimental Psychology. A, Experimentální psychologie člověka. 45 (2): 177–92. doi:10.1080/14640749208401323. PMID 1410554. S2CID 32594562.
- ^ Waters GS, Rochon E, Caplan D (únor 1992). „Role plánování řeči na vysoké úrovni při zkoušce: Důkazy od pacientů s apraxií řeči“. Deník paměti a jazyka. 31 (1): 54–73. doi:10.1016 / 0749-596x (92) 90005-i.
- ^ Cohen L, Bachoud-Levi AC (září 1995). "Role výstupního fonologického bufferu při řízení načasování řeči: jediná případová studie". Kůra; Časopis věnovaný studiu nervového systému a chování. 31 (3): 469–86. doi:10.1016 / s0010-9452 (13) 80060-3. PMID 8536476. S2CID 4480375.
- ^ Shallice T, Rumiati RI, Zadini A (září 2000). Msgstr "Selektivní poškození fonologické výstupní vyrovnávací paměti". Kognitivní neuropsychologie. 17 (6): 517–46. doi:10.1080/02643290050110638. PMID 20945193. S2CID 14811413.
- ^ Shu H, Xiong H, Han Z, Bi Y, Bai X (2005). „Selektivní poškození fonologické výstupní vyrovnávací paměti: důkazy od čínského pacienta“. Behaviorální neurologie. 16 (2–3): 179–89. doi:10.1155/2005/647871. PMC 5478832. PMID 16410633.
- ^ Oberauer K (2002). "Přístup k informacím v pracovní paměti: Zkoumání ohniska pozornosti". Journal of Experimental Psychology: Learning, Memory, and Cognition. 28 (3): 411–421. doi:10.1037/0278-7393.28.3.411. PMID 12018494.
- ^ Unsworth N, Engle RW (leden 2007). "Povaha jednotlivých rozdílů v kapacitě pracovní paměti: aktivní údržba v primární paměti a řízené vyhledávání ze sekundární paměti". Psychologický přehled. 114 (1): 104–32. doi:10.1037 / 0033-295x.114.1.104. PMID 17227183.
- ^ Barrouillet P, Camos V (prosinec 2012). "Postupem času". Současné směry v psychologické vědě. 21 (6): 413–419. doi:10.1177/0963721412459513. S2CID 145540189.
- ^ Bornkessel-Schlesewsky I, Schlesewsky M, Small SL, Rauschecker JP (březen 2015). „Neurobiologické kořeny jazyka v konkurzu primátů: společné výpočetní vlastnosti“. Trendy v kognitivních vědách. 19 (3): 142–50. doi:10.1016 / j.tics.2014.12.008. PMC 4348204. PMID 25600585.
- ^ Buchsbaum BR, D'Esposito M (květen 2008). Msgstr "Hledání fonologického obchodu: od smyčky po konvoluci". Journal of Cognitive Neuroscience. 20 (5): 762–78. doi:10.1162 / jocn.2008.20501. PMID 18201133. S2CID 17878480.
- ^ Miller LM, Recanzone GH (duben 2009). „Populace sluchových kortikálních neuronů mohou přesně kódovat akustický prostor napříč intenzitou stimulu“. Sborník Národní akademie věd Spojených států amerických. 106 (14): 5931–5. doi:10.1073 / pnas.0901023106. PMC 2667094. PMID 19321750.
- ^ Tian B, Reser D, Durham A, Kustov A, Rauschecker JP (duben 2001). "Funkční specializace na sluchovou kůru opice rhesus". Věda. 292 (5515): 290–3. doi:10.1126 / science.1058911. PMID 11303104. S2CID 32846215.
- ^ Alain C, Arnott SR, Hevenor S, Graham S, Grady CL (říjen 2001). ""Co "a" kde "v lidském sluchovém systému". Sborník Národní akademie věd Spojených států amerických. 98 (21): 12301–6. doi:10.1073 / pnas.211209098. PMC 59809. PMID 11572938.
- ^ De Santis L, Clarke S, Murray MM (leden 2007). „Automatické a vnitřní sluchové„ co “a„ kde “zpracování u lidí odhalené elektrickým neuroimagingem. Mozková kůra. 17 (1): 9–17. doi:10.1093 / cercor / bhj119. PMID 16421326.
- ^ Barrett DJ, Hall DA (srpen 2006). "Předvolby odpovědí na" co "a" kde "v lidské neprimární sluchové kůře". NeuroImage. 32 (2): 968–77. doi:10.1016 / j.neuroimage.2006.03.050. PMID 16733092. S2CID 19988467.
- ^ Linden JF, Grunewald A, Andersen RA (červenec 1999). „Odpovědi na sluchové podněty v postranní intraparietální oblasti makaků. II. Modulace chování“. Journal of Neurophysiology. 82 (1): 343–58. doi:10.1152 / jn.1999.82.1.343. PMID 10400963.
- ^ Mazzoni P, Bracewell RM, Barash S, Andersen RA (březen 1996). "Prostorově vyladěné sluchové odpovědi v oblasti LIP makaků provádějících zpožděné paměťové sakády na akustické cíle". Journal of Neurophysiology. 75 (3): 1233–41. doi:10.1152 / jn.1996.75.3.1233. PMID 8867131.
- ^ Lachaux JP, Jerbi K, Bertrand O, Minotti L, Hoffmann D, Schoendorff B, Kahane P (říjen 2007). „Plán pro funkční mapování v reálném čase prostřednictvím lidských intrakraniálních záznamů“. PLOS ONE. 2 (10): e1094. doi:10.1371 / journal.pone.0001094. PMC 2040217. PMID 17971857.
- ^ Jardri R, Houfflin-Debarge V, Delion P, Pruvo JP, Thomas P, Pins D (duben 2012). "Hodnocení reakce plodu na řeč matky pomocí neinvazivní funkční zobrazovací techniky mozku". International Journal of Developmental Neuroscience. 30 (2): 159–61. doi:10.1016 / j.ijdevneu.2011.11.002. PMID 22123457. S2CID 2603226.
- ^ Poliva O (2017-09-20). „Odkud k čemu: neuroanatomicky založený evoluční model vzniku řeči u lidí“. F1000Výzkum. 4: 67. doi:10.12688 / F1000Research.6175.3. PMC 5600004. PMID 28928931.
- ^ Poliva O (2016-06-30). „Od mimikry k jazyku: neuroanatomicky založený evoluční model vzniku hlasového jazyka“. Frontiers in Neuroscience. 10: 307. doi:10.3389 / fnins.2016.00307. PMC 4928493. PMID 27445676.
- ^ A b C d E Suri, Sana. „Jaká znaková řeč nás učí o mozku“. Konverzace. Citováno 2019-10-07.
- ^ A b C Scientific American. (2002). Znaková řeč v mozku. [Brožura]. Citováno z http://lcn.salk.edu/Brochure/SciAM%20ASL.pdf
- ^ A b C d E F G h i j k l Norton ES, Kovelman I, Petitto LA (březen 2007). „Existují oddělené neurální systémy pro pravopis? Nové poznatky o roli pravidel a paměti v pravopisu při zobrazování funkční magnetickou rezonancí“. Mysl, mozek a vzdělání. 1 (1): 48–59. doi:10.1111 / j.1751-228X.2007.00005.x. PMC 2790202. PMID 20011680.
- ^ A b C d E Treiman R, Kessler B (2007). Psací systémy a vývoj pravopisu. The Science of Reading: A Handbook. Blackwell Publishing Ltd. str. 120–134. doi:10.1002 / 9780470757642.ch7. ISBN 978-0-470-75764-2.