Superior colliculus - Superior colliculus
| Superior colliculus | |
|---|---|
 Schéma superior colliculus (L) člověka střední mozek (zobrazeno červeně) a okolní oblasti. Nadřazený colliculus je obklopen červeným kroužkem a průhledným červeným kruhem, který označuje jeho umístění. | |
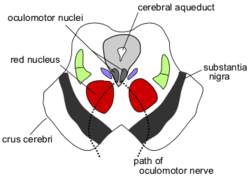 Řez středním mozkem na úrovni superior colliculus ukazující cestu okulomotorický nerv | |
| Detaily | |
| Část | Tectum |
| Identifikátory | |
| latinský | Colliculus superior |
| Pletivo | D013477 |
| NeuroNames | 473 |
| NeuroLex ID | birnlex_1040 |
| TA98 | A14.1.06.015 |
| TA2 | 5912 |
| TH | H3.11.03.3.01002 |
| TE | E5.14.3.3.1.4.4 |
| FMA | 62403 |
| Anatomické pojmy neuroanatomie | |
The superior colliculus (latinský pro "horní kopec") je stavba ležící na střecha savců střední mozek.[1] U nesavců obratlovců, homologní struktura je známá jako optické tektumnebo optický lalok.[1][2][3] Forma adjektiva tektální se běžně používá pro obě struktury.
U savců tvoří superior colliculus hlavní součást středního mozku. Jedná se o spárovanou strukturu a společně se spárovanými inferior colliculi tvoří corpora quadrigemina. Superior colliculus je vrstvená struktura, s řadou vrstev, které se liší podle druhů. Vrstvy lze seskupit do povrchových vrstev (stratum opticum a výše) a hlubší zbývající vrstvy. Neurony v povrchových vrstvách dostávají přímý vstup ze sítnice a reagují téměř výlučně na vizuální podněty. Mnoho neuronů v hlubších vrstvách také reaguje na jiné modality a některé reagují na podněty v různých modalitách.[4] Hlubší vrstvy také obsahují populaci neuronů souvisejících s motorem, schopných aktivovat pohyby očí i další reakce.[5]
Obecnou funkcí tektálního systému je nasměrovat behaviorální reakce na konkrétní body v egocentrickém („střed těla“) prostoru. Každá vrstva obsahuje a topografická mapa okolního světa v retinotopický souřadnice a aktivace neuronů v určitém bodě mapy evokuje reakci směřující k odpovídajícímu bodu ve vesmíru. U primátů byl superior colliculus studován hlavně s ohledem na jeho roli při řízení pohybů očí. Vizuální vstup ze sítnice nebo „příkazový“ vstup z mozkové kůry vytvoří „bouli“ aktivity v tektální mapě, která, je-li dostatečně silná, vyvolá sakadický pohyb očí. Dokonce i u primátů se však superior colliculus také podílí na generování prostorově zaměřených otočení hlavy, pohybů dosahujících paží,[6] a posuny pozornosti, které nezahrnují žádné zjevné pohyby.[7] U jiných druhů je superior colliculus zapojen do široké škály odpovědí, včetně obratů celého těla u kráčících potkanů. U savců, a zejména u primátů, masivní expanze mozkové kůry redukuje colliculus superior na mnohem menší část celého mozku. Zůstává nicméně důležitý z hlediska funkce primárního integračního centra pro pohyby očí.
U druhů, které nejsou savci, je optické tektum zapojeno do mnoha reakcí, včetně plavání v rybách, létání u ptáků, úderů jazykem za kořistí u žab a úderů tesáků u hadů. U některých druhů, včetně ryb a ptáků, je optické tektum, známé také jako optický lalok, jednou z největších složek mozku.
Poznámka k terminologii: Tento článek sleduje terminologii zavedenou v literatuře, používající výraz „superior colliculus“ při diskusi o savcích a „optické tektum“ při diskusi o konkrétních jiných než savčích druzích nebo obecně o obratlovcích.
Struktura


Superior colliculus je synaptická vrstvená struktura.[8] Dva vynikající colliculi sedí pod thalamus a obklopit epifýza v savčí střední mozek. Zahrnuje hřbetní aspekt střední mozek, zadní k periaqueductal šedá a okamžitě lepší než inferior colliculus. Inferior a superior colliculi jsou souhrnně označovány jako corpora quadrigemina (Latinský, čtyřnásobná těla). Superior colliculi jsou větší než inferior colliculi, ačkoli nižší colliculi jsou výraznější.[9] The brachium superior colliculus (nebo vynikající brachium) je větev, která sahá bočně od superior colliculus, a procházející mezi pulvinar a mediální geniculate tělo, částečně pokračuje do eminence zvané boční geniculate tělo, a částečně do optický trakt.
Nadřazený colliculus je spojen s blízkou strukturou zvanou parabigeminální jádro, často označovaný jako jeho satelit. V optickém tektu je tato blízká struktura známá jako nucleus isthmi.
Neurální obvody
Mikrostruktura superior colliculus a optického tektum se u jednotlivých druhů liší. Obecně platí, že vždy existuje jasný rozdíl mezi povrchovými vrstvami, které přijímají vstup primárně z vizuálního systému a vykazují primárně vizuální reakce, a hlubšími vrstvami, které přijímají mnoho typů vstupů a promítají se do mnoha oblastí mozku souvisejících s motorem. Rozdíl mezi těmito dvěma zónami je tak jasný a konzistentní, že někteří anatomové navrhli, že by měly být považovány za samostatné mozkové struktury.
U savců neuroanatomisté obvykle identifikují sedm vrstev[10] Horní tři vrstvy se nazývají povrchní:
- Lamina I. nebo SZ, stratum zonale, je tenká vrstva skládající se z malých myelinovaných axonů spolu s okrajovými a vodorovnými buňkami.
- Lamina II nebo SGS, stratum griseum superficiale („povrchová šedá vrstva“), obsahuje mnoho neuronů různých tvarů a velikostí.
- Lamina III nebo TAK, stratum opticum ("optická vrstva"), se skládá hlavně z axonů pocházejících z optický trakt.
Dále přijdou dva mezivrstvy:
- Lamina IV nebo SGI, stratum griseum intermedium („střední šedá vrstva“), je nejsilnější vrstvou a je vyplněna mnoha neurony mnoha velikostí. Tato vrstva je často stejně silná jako všechny ostatní vrstvy dohromady. Často se dělí na „horní“ a „spodní“ část.
- Lamina V nebo SAI, stratum album intermedium („mezilehlá bílá vrstva“), sestává hlavně z vláken z různých zdrojů.
Nakonec přijdou dva hluboké vrstvy:
- Lamina VI nebo SGP, stratum griseum profundum („temně šedá vrstva“), sestává z volně zabalených neuronů a myelinizovaných vláken.
- Lamina VII nebo MÍZA, stratum album profundum ("hluboká bílá vrstva"), ležící přímo nad periaqueductal šedá, sestává výhradně z vláken.
Povrchové vrstvy přijímají vstup hlavně ze sítnice, z oblastí mozkové kůry souvisejících se zrakem a ze dvou tektálních struktur nazývaných pretectum a parabigeminální jádro. Vstup sítnice zahrnuje celou povrchovou zónu a je oboustranný, i když kontralaterální část je rozsáhlejší. Kortikální vstup pochází nejvíce z primární zraková kůra (oblast 17), sekundární zraková kůra (oblasti 18 a 19 ) a pole čelního oka. Parabigeminální jádro hraje velmi důležitou roli v tektální funkci, která je popsána níže.
Na rozdíl od zrakově ovládaných vstupů do povrchových vrstev přijímají mezilehlé a hluboké vrstvy vstupy z velmi rozmanité sady senzorických a motorických struktur. Většina oblastí mozkové kůry vyčnívá do těchto vrstev, ačkoli vstup z „asociačních“ oblastí má tendenci být těžší než vstup z primárních senzorických nebo motorických oblastí.[Citace je zapotřebí ] Ovšem zúčastněné kortikální oblasti a síla jejich relativních výčnělků se u jednotlivých druhů liší.[11] Další důležitý vstup pochází z substantia nigra, pars reticulata, součást bazální ganglia. Tato projekce využívá inhibiční neurotransmiter GABA, a předpokládá se, že působí „hradlovým“ účinkem na nadřazený colliculus. Mezilehlé a hluboké vrstvy také přijímají vstup z spinální trigeminální jádro, který přenáší somatosenzorické informace z obličeje, stejně jako hypotalamus, zona incerta, thalamus, a inferior colliculus.
Kromě svých charakteristických vstupů mají povrchové a hluboké zóny superior colliculus také výrazné výstupy. Jeden z nejdůležitějších výstupů jde do pulvinar a boční mezilehlé oblasti thalamu, které se zase promítají do oblastí mozkové kůry, které se podílejí na ovládání pohybů očí. Existují také projekce z povrchové zóny do pretektálních jader, boční geniculate jádro thalamu a parabigeminálního jádra. Projekce z hlubších vrstev jsou rozsáhlejší. Existují dvě velké sestupné dráhy, cestování do mozkového kmene a míchy, a četné vzestupné projekce do různých senzorických a motorických center, včetně několika, které se podílejí na generování pohybů očí.
Oba colliculi mají také sestupné projekce na paramediánní pontinovou retikulární formaci a míchu, a proto mohou být zapojeny do odpovědí na podněty rychleji, než by kortikální zpracování umožňovalo.
Mozaiková struktura
Při podrobném prozkoumání kolikulární vrstvy ve skutečnosti nejsou hladké listy, ale jsou rozděleny do voštinového uspořádání diskrétních sloupců.[12] Nejjasnější indikace sloupcové struktury pochází z cholinergních vstupů vznikajících z parabigeminálního jádra, jehož terminály tvoří rovnoměrně rozmístěné shluky, které se táhnou shora dolů tektum.[13] Několik dalších neurochemických markerů včetně kalretininu, parvalbuminu, GAP-43 a NMDA receptorů a spojení s mnoha dalšími mozkovými strukturami v mozkovém kmeni a diencephalonu také vykazuje odpovídající nehomogenitu.[14] Celkový počet sloupců se odhaduje na přibližně 100.[12] Funkční význam této sloupové architektury není jasný, ale je zajímavé, že nedávné důkazy implikují cholinergní vstupy jako součást rekurentního obvodu produkujícího v tektu dynamiku vítěz-bere vše, jak je podrobněji popsáno níže.
Všechny druhy, které byly zkoumány - včetně savců a jiných než savců - vykazují rozčlenění, ale v podrobnostech uspořádání jsou určité systematické rozdíly.[13] U druhů se sítnicí pruhovaného typu (hlavně u druhů s bočně umístěnými očima, jako jsou králíci a jeleni), oddělení pokrývá celý rozsah SC. U druhů s centrálně umístěnou foveou se však kompartmentalizace rozpadá v přední (rostrální) části SC. Tato část SC obsahuje mnoho „fixačních“ neuronů, které nepřetržitě střílejí, zatímco oči zůstávají fixovány v konstantní poloze.
Funkce
Historie vyšetřování optického tektu byla poznamenána několika velkými posuny v názorech. Asi před rokem 1970 se většina studií zabývala jinými než savci - rybami, žabami, ptáky - tedy druhy, u nichž je optická tektum dominantní strukturou přijímající vstup z očí. Obecný názor tedy byl, že optické tektum je u těchto druhů hlavním vizuálním centrem v mozku, které není savcem, a v důsledku toho se účastní široké škály chování[Citace je zapotřebí ]. V sedmdesátých a devadesátých letech se však nervové záznamy savců, většinou opic, zaměřovaly především na roli superior colliculus při kontrole pohybů očí. Tato linie vyšetřování začala ovládat literaturu do takové míry, že většinový názor byl ten, že kontrola pohybu očí je u savců jedinou důležitou funkcí, což se v mnoha současných učebnicích stále odráží.
Na konci 90. let však experimenty se zvířaty, jejichž hlavy se mohly volně pohybovat, jasně ukázaly, že SC ve skutečnosti produkuje pohledy se posunou, obvykle složené spíše z kombinovaných pohybů hlavy a očí než z pohybů očí per se. Tento objev probudil zájem o celou šíři funkcí nadřízeného colliculus a vedl ke studiu multisenzorická integrace v různých druzích a situacích. Role SC při řízení pohybů očí je však chápána v mnohem větší hloubce než jakákoli jiná funkce.
Behaviorální studie ukázaly, že SC není pro rozpoznávání objektů potřeba, ale hraje zásadní roli ve schopnosti nasměrovat chování ke konkrétním objektům a může tuto schopnost podporovat i v nepřítomnosti mozkové kůry.[15] Kočky s velkým poškozením zrakové kůry tedy nemohou rozpoznávat předměty, ale přesto mohou být schopny sledovat a orientovat se na pohybující se podněty, i když pomaleji než obvykle. Pokud je však odstraněna jedna polovina SC, kočky budou neustále kroužit směrem ke straně léze a nutně se orientovat na objekty, které se tam nacházejí, ale vůbec se neorientují na objekty umístěné v opačném poloměrem pole. Tyto deficity se časem snižují, ale nikdy nezmizí.
Pohyby očí
U primátů pohyby očí lze rozdělit do několika typů: fixace, ve kterém jsou oči směrovány k nehybnému předmětu, pohyby očí slouží pouze k vyrovnání pohybů hlavy; hladké pronásledování, ve kterém se oči neustále pohybují, aby sledovaly pohybující se předmět; sakády, ve kterém se oči velmi rychle pohybují z jednoho místa na druhé; a vergence, ve kterém se oči pohybují současně v opačných směrech za účelem získání nebo udržení jediného binokulárního vidění. Ve všech je zapojen superior colliculus, ale jeho role v saccade byla studována nejintenzivněji.
Každý ze dvou colliculi - jeden na každé straně mozku - obsahuje dvourozměrnou mapu představující polovinu zorného pole. The fovea - oblast maximální citlivosti - je znázorněna na předním okraji mapy a na okraji na zadním okraji. Pohyby očí jsou vyvolávány aktivitou v hlubokých vrstvách SC. Během fixace jsou neurony blízko předního okraje - foveální zóny - tonicky aktivní. Během plynulého pronásledování se aktivují neurony v malé vzdálenosti od předního okraje, což vede k malým pohybům očí. U saccade jsou neurony aktivovány v oblasti, která představuje bod, do kterého bude saccade nasměrována. Těsně před saccade se aktivita rychle hromadí v cílovém místě a klesá v jiných částech SC. Kódování je poměrně široké, takže pro jakoukoli danou sakádu profil aktivity tvoří „kopec“, který zahrnuje podstatnou část kolikulární mapy: Umístění vrcholu tohoto „kopce“ představuje cíl sakády.
SC kóduje cíl posunu pohledu, ale nezdá se, že by specifikoval přesné pohyby potřebné k dosažení cíle.[16] Rozklad posunu pohledu na pohyby hlavy a očí a přesná trajektorie oka během sakády závisí na integraci kolikulárních a nekolikulárních signálů po motorických oblastech po proudu, a to způsoby, které dosud nejsou dobře pochopeny. Bez ohledu na to, jak je pohyb vyvoláván nebo prováděn, jej SC kóduje do „retinotopických“ souřadnic: to znamená, že poloha kopce SC odpovídá pevnému umístění na sítnici. Zdá se, že to odporuje pozorování, že stimulace jediný bod na SC může mít za následek různé směry posunu pohledu, v závislosti na počáteční orientaci očí. Ukázalo se však, že je to proto, že umístění stimulu v sítnici je nelineární funkcí umístění cíle, orientace oka a sférická geometrie oka.[17]
Existují určité diskuse o tom, zda SC pouze ovládá pohyby očí a ponechává popravu jiným strukturám, nebo zda se aktivně podílí na výkonu sakády. V roce 1991 tvrdili Munoz a kol. Na základě údajů, které shromáždili, že během sakády se „kopec“ aktivity v SC pohybuje postupně, aby odrážel měnící se posun oka od cílového místa, zatímco sakáda postupuje.[18] V současné době převládá názor, že ačkoli se „kopec“ během sakády mírně posouvá, nemění se stabilním a přiměřeným způsobem, jak předpovídá hypotéza „pohyblivého kopce“.[19] Pohybující se kopce však mohou hrát v nadřazeném colliculus jinou roli; novější experimenty prokázaly nepřetržitě se pohybující kopec aktivity vizuální paměti, když se oči pohybují pomalu, zatímco je zachován samostatný sakádový cíl.[20]
Výstup z motorického sektoru SC jde do sady jader středního mozku a mozkového kmene, které transformují kód „place“ používaný SC na kód „rate“ používaný okulomotorickými neurony. Pohyby očí jsou generovány šesti svaly uspořádanými do tří ortogonálně vyrovnaných párů. Na úrovni konečné společné dráhy jsou tedy pohyby očí zakódovány v podstatě do kartézského souřadnicového systému.
Ačkoli SC dostává silný vstup přímo ze sítnice, u primátů je z velké části pod kontrolou mozkové kůry, která obsahuje několik oblastí, které se podílejí na určování pohybů očí.[21] The pole čelního oka, část motorické kůry, se účastní spouštění úmyslných sakád a sousední oblast, doplňková oční pole, se účastní organizování skupin sakád do sekvencí. Temenní oční pole, která jsou dále v mozku, jsou zapojena hlavně do reflexních váčků vytvořených v reakci na změny v pohledu.
SC pouze přijímá vizuální vstupy v jeho povrchových vrstvách, zatímco hlubší vrstvy colliculus přijímají také sluchové a somatosenzorické vstupy a jsou spojeny s mnoha senzomotorickými oblastmi mozku. Koliculus jako celek je myšlenka pomoci orientovat hlavu a oči směrem k něčemu viděnému a slyšenému.[7][22][23][24]
Nadřazený colliculus také přijímá sluchové informace z dolního colliculus. Tato sluchová informace je integrována s vizuálními informacemi, které jsou již přítomny, aby se vytvořil břichomluvecký efekt.
Roztržitost
Kromě souvislosti s pohyby očí se zdá, že SC hraje důležitou roli v obvodech podporujících rozptýlení. Při normálním stárnutí dochází ke zvýšené roztržitosti [25] a je také ústředním prvkem řady zdravotních stavů, včetně porucha pozornosti s hyperaktivitou (ADHD).[26] Výzkum ukázal, že léze SC u mnoha druhů mohou vést ke zvýšené roztržitosti[27][28] a, u lidí, odstranění inhibiční kontroly na SC z pre-frontální kůry, tedy zvýšení aktivity v oblasti, také zvyšuje roztržitost.[29] Výzkum na zvířecím modelu ADHD, spontánně hypertenzního potkana, také ukazuje změněné chování závislé na kolikulární struktuře[30][31] a fyziologie.[31] Amfetamin (hlavní léčba ADHD) navíc potlačuje aktivitu v colliculus u zdravých zvířat.[32]
Ostatní zvířata
Ostatní savci
Primáti
Obvykle se uznává, že primát superior colliculus je mezi savci, protože neobsahuje úplnou mapu zorného pole viděného kontralaterálním okem. Místo toho jako vizuální kůra a boční geniculate jádro, každý colliculus představuje pouze kontralaterální polovinu zorné pole, až do středové čáry, a vylučuje reprezentaci ipsilaterální poloviny.[33] Tato funkční charakteristika je vysvětlena absencí anatomických spojení mezi primáty gangliové buňky sítnice v časové polovině sítnice a kontralaterální superior colliculus. U ostatních savců se gangliové buňky sítnice skrz kontralaterální sítnici promítají do kontralaterálního colliculus. Toto rozlišení mezi primáty a jinými primáty je jednou z klíčových linií důkazů na podporu teorie létajících primátů navrhl australský neurolog Jack Pettigrew v roce 1986 poté, co zjistil, že létající lišky (megabaty ) připomínají primáty, pokud jde o vzor anatomických spojení mezi sítnicí a superior colliculus.[34]
Kočky
V kočce vyčnívá přes colliculus superior retikulární formace a interaguje s motorickými neurony v mozkový kmen.[35]
Netopýři
Netopýři nejsou ve skutečnosti slepí, ale mnohem více závisí na echolokaci než na vidění pro navigaci a zachycení kořisti. Získávají informace o okolním světě vydáváním sonarových pípnutí a následným posloucháním ozvěn. Jejich mozky jsou pro tento proces vysoce specializované a některé z těchto specializací se objevují v superior colliculus.[36] U netopýrů zaujímá projekce sítnice pouze tenkou zónu těsně pod povrchem, ale existují rozsáhlé vstupy ze sluchových oblastí a výstupy do motorických oblastí schopných orientovat uši, hlavu nebo tělo. Ozvěny přicházející z různých směrů aktivují neurony na různých místech v kolikulárních vrstvách,[37] a aktivace kolikulárních neuronů ovlivňuje cvrlikání, které vydávají netopýři. Existuje tedy silný případ, že superior colliculus vykonává stejné druhy funkcí pro sluchově řízené chování netopýrů, které provádí pro vizuálně vedené chování jiných druhů.
Netopýři se obvykle dělí do dvou hlavních skupin: Microchiroptera (nejpočetnější a běžně se vyskytující po celém světě) a Megachiroptera (kaloni, vyskytující se v Asii, Africe a Austrálii). Až na jednu výjimku Megabaty nevystavujte echolokaci a při navigaci se spoléhejte na rozvinutý smysl pro vidění. Vizuální receptivní pole neuronů v superior colliculus u těchto zvířat tvoří přesnou mapu sítnice, podobný tomu nalezenému v kočky a primáti.
Hlodavci
Předpokládá se, že superior colliculus u hlodavců zprostředkovává smyslově řízený přístup a vyhýbací chování.[38][39] Studie využívající nástroje pro analýzu obvodů na myši superior colliculus odhalily několik důležitých funkcí.[8]
Non-savčí obratlovci
Optické tektum


The optické tektum je vizuální centrum v mozku jiných než savců, které se vyvíjí z výstražná deska mezencephalonu. U savců jsou spojení z optického tektu důležitá pro rozpoznávání a reakci na objekty různých velikostí, což usnadňují vysílače excitačních optických nervů, jako jsou L-glutamát.[41]
Narušující vizuální zážitek na začátku roku zebrafish vývoj vede ke změně tektální aktivity. Změny v tektální aktivitě vyústily v neschopnost úspěšně lovit a chytat kořist.[42] Inhibiční signalizace hypotalamu do hlubokého tektálu neuropil je důležitý při tektálním zpracování u larev zebrafish. Tektální neuropil obsahuje struktury včetně periventrikulární neurony axony a dendrity. Neuropil také obsahuje GABAergický povrchové inhibiční neurony umístěné v stratum opticum.[43] Namísto velké mozkové kůry mají zebrafish relativně velké optické tektum, u kterého existuje hypotéza, že provádí některé vizuální zpracování, které kůra provádí u savců.[44]
Nedávné studie lézí naznačují, že optické tektum nemá žádný vliv na pohybové reakce vyššího řádu OMR nebo OKR,[45] ale může být integrálnější s narážkami nižšího řádu ve vnímání pohybu, jako při identifikaci malých předmětů.[46]
Optické tektum je jednou ze základních složek mozek obratlovců, existující u řady druhů.[47] Některé aspekty struktury jsou velmi konzistentní, včetně struktury složené z několika vrstev, s hustým vstupem z optických traktů do povrchových vrstev a dalším silným vstupem, který přenáší somatosenzorický vstup do hlubších vrstev. Ostatní aspekty jsou velmi variabilní, například celkový počet vrstev (od 3 u afrických plic po 15 u zlatých rybek)[48]) a počet různých typů buněk (od 2 do lungfish do 27 v house sparrow[48]).
Optické tektum je úzce spojeno se sousední strukturou zvanou nucleus isthmi, která vzbudila velký zájem, protože evidentně velmi významně přispívá k tektální funkci.[49] (V superior colliculus podobná struktura se nazývá parabigeminální jádro). Jádro isthmii je rozděleno na dvě části, tzv isthmus pars magnocellularis (Obr; "část s velkými buňkami") a isthmus pars parvocellularis (Ipc; "část s malými buňkami"). Spojení mezi třemi oblastmi - optické tektum, Ipc a Imc - jsou topografické. Neurony v povrchových vrstvách optického tektu se promítají do odpovídajících bodů v Ipc a Imc. Projekce na Ipc jsou úzce zaměřeny, zatímco projekce na Imc jsou poněkud rozptýlenější. Ipc vede k úzce zaměřeným cholinergním projekcím jak pro Imc, tak pro optické tektum. V optickém tektu se cholinergní vstupy z Ipc rozvětvují a vytvářejí terminály, které se táhnou přes celý sloupec, shora dolů. Naproti tomu Imc vede k projekcím GABAergic na Ipc a optické tektum, které se šíří v bočních rozměrech velmi široce a zahrnují většinu retinotopických map. Okruh tectum-Ipc-Imc tedy způsobí, že tektální aktivita produkuje opakovanou zpětnou vazbu, která zahrnuje úzce zaměřenou excitaci malého sloupce sousedních tektálních neuronů spolu s globální inhibicí vzdálených tektálních neuronů.
Optické tektum se účastní mnoha reakcí, včetně plavání v rybách, letu u ptáků, úderů jazykem směrem ke kořisti u žab a úderů zubů u hadů. U některých druhů, včetně ryb a ptáků, je optické tektum, známé také jako optický lalok, jednou z největších složek mozku.
U hagfish, mihule a žraloka je to relativně malá stavba, ale v teleost ryby se značně rozšiřuje, v některých případech se stává největší strukturou v mozku. U obojživelníků, plazů a zejména ptáků je také velmi významnou složkou.[48]
v hadi který dokáže detekovat infračervené záření, jako pythons a zmije, počáteční neurální vstup je přes trigeminální nerv místo optický trakt. Zbytek zpracování je podobný zpracování vizuálního smyslu a zahrnuje tedy optické tektum.[50]
Ryba

Mihule
The mihule byl rozsáhle studován, protože má relativně jednoduchý mozek, o kterém se v mnoha ohledech předpokládá, že odráží mozkovou strukturu časných předků obratlovců. Od sedmdesátých let minulého století Sten Grillner a jeho kolegové z Karolinska Institute ve Stockholmu používají mihule obecnou jako modelový systém k vypracování základních principů motorické kontroly u obratlovců, počínaje míchou a směrem nahoru do mozku.[51] V sérii studií zjistili, že nervové obvody v míše jsou schopné generovat rytmické motorické vzorce, které jsou základem plavání, že tyto obvody jsou ovládány specifickými pohybovými oblastmi v mozkovém kmeni a středním mozku a že tyto oblasti jsou zase kontrolovány vyššími strukturami mozku, včetně bazální ganglia a tektum. Ve studii o mihule tektové publikované v roce 2007[52] zjistili, že elektrická stimulace může vyvolat pohyby očí, pohyby v bočním ohybu nebo plaveckou aktivitu a že typ, amplituda a směr pohybu se mění v závislosti na místě v stimulovaném tektu. Tato zjištění byla interpretována v souladu s myšlenkou, že tektum generuje cílenou lokomoci v mihule, jako je tomu u jiných druhů.
Ptactvo

U ptáků je optické tektum zapojeno do letu a je jednou z největších složek mozku. Studie vizuálního zpracování ptáků umožnila lépe porozumět tomu u savců, včetně lidí.[53]
Viz také
Další obrázky

Schéma znázorňující centrální spojení optické nervy a optické trakty. (Superior colliculus viditelný blízko středu.)

Superior colliculus

Mozkový kmen. Pohled zezadu.



Poznámky
- ^ A b Squire, L (2013). Základní neurověda (Čtvrté vydání). str. 707. ISBN 9780123858702.
- ^ Knudsen, EI (červen 2011). „Kontrola zdola: role sítě středního mozku v prostorové pozornosti“. Evropský žurnál neurovědy. 33 (11): 1961–72. doi:10.1111 / j.1460-9568.2011.07696.x. PMC 3111946. PMID 21645092.
- ^ Liu, Tsung-Han; Chiao, Chuan-Chin (25. ledna 2017). „Mozaiková organizace kontroly tělesných vzorů v optickém laloku chobotnic“. Journal of Neuroscience. 37 (4): 768–780. doi:10.1523 / JNEUROSCI.0768-16.2016. PMC 6597019. PMID 28123014.
- ^ Wallace a kol., 1998
- ^ Gandhi a kol., 2011
- ^ Lunenburger a kol., 2001
- ^ A b Kustov a Robinson, 1996
- ^ A b Ito, S; Feldheim, DA (2018). „Moci Superior Colliculus: rozvíjející se model pro studium formování a funkce obvodu“. Hranice v neurálních obvodech. 12: 10. doi:10.3389 / fncir.2018.00010. PMC 5816945. PMID 29487505.
- ^ „IX. Neurologie. 4b. Střední mozek neboli Mesencephalon. Gray, Henry. 1918. Anatomie lidského těla“. www.bartleby.com. Citováno 10. října 2019.
- ^ Huerta & Harting, 1984
- ^ Clemo HR, Stein BE (1984). "Topografická organizace somatosenzorických kortikotektálních vlivů u koček". Journal of Neurophysiology. 51 (5): 843–858. doi:10.1152 / jn.1984.51.5.843. PMID 6726314.
- ^ A b Chavalier & Mana, 2000
- ^ A b Illing, 1996
- ^ Mana & Chevalier, 2001
- ^ Sprague, 1996
- ^ Sparks & Gandhi, 2003
- ^ Klier a kol., 2001
- ^ Munoz a kol., 1991
- ^ Soetedjo et al., 2002
- ^ Dash a kol., 2015
- ^ Pierrot-Deseilligny a kol., 2003
- ^ Klier a kol., 2003
- ^ Krauzlis a kol., 2004
- ^ Sparks, 1999
- ^ Prendergast, M. A .; Jackson, W. J .; Terry, A. V .; Kille, N.J .; Arneric, S. P .; Decker, M. W .; Buccafusco, J. J. (01.03.1998). „Věkové rozdíly v roztržitosti a reakci na methylfenidát u opic“. Mozková kůra. 8 (2): 164–172. doi:10.1093 / cercor / 8.2.164. ISSN 1047-3211. PMID 9542895.
- ^ Douglas, V (1983). Vývojová neuropsychiatrie. New York: Guildford Press. 280–329.
- ^ Goodale, M. A .; Foreman, N. P .; Milner, A. D. (01.03.1978). "Vizuální orientace u krysy: Disociace deficitů po kortikálních a kolikulárních lézích". Experimentální výzkum mozku. 31 (3): 445–457. doi:10.1007 / BF00237301. ISSN 0014-4819. PMID 648607.
- ^ Milner, A. D .; Foreman, N. P .; Goodale, M. A. (01.01.1978). "Go-left go-right diskriminační výkon a rozptýlení po lézích prefrontální kůry nebo superior colliculus u stumptail makaků". Neuropsychologie. 16 (4): 381–390. doi:10.1016/0028-3932(78)90062-3. PMID 99682.
- ^ Gaymard, Bertrand; François, Chantal; Ploner, Christoph J .; Condy, Carine; Rivaud-Péchoux, Sophie (01.04.2003). "Přímý prefrontotektální trakt proti roztržitosti v lidském mozku". Annals of Neurology. 53 (4): 542–545. doi:10,1002 / ana.10560. ISSN 1531-8249. PMID 12666125.
- ^ Dommett, Eleanor J .; Rostron, Claire L. (2011-11-01). „Abnormální chování při usměrňování vzduchu v modelu spontánně hypertenzní krysy ADHD“. Experimentální výzkum mozku. 215 (1): 45–52. doi:10.1007 / s00221-011-2869-7. ISSN 0014-4819. PMID 21931982.
- ^ A b Brace, L.R .; Kraev, I .; Rostron, C.L .; Stewart, M.G .; Overton, P.G .; Dommett, E.J. (2015). "Změněné vizuální zpracování v modelu hlodavců s poruchou pozornosti a hyperaktivity". Neurovědy. 303: 364–377. doi:10.1016 / j.neuroscience.2015.07.003. PMID 26166731.
- ^ Clements, K.M .; Devonshire, I.M .; Reynolds, J.N.J .; Overton, P.G. (2014). „Vylepšené vizuální reakce v superior colliculus na zvířecím modelu poruchy pozornosti s hyperaktivitou a jejich potlačení d-amfetaminem“. Neurovědy. 274: 289–298. doi:10.1016 / j.neuroscience.2014.05.054. PMID 24905438.
- ^ Lane a kol., 1973
- ^ Pettigrew, 1986
- ^ Precht, W. (1974). "Tektální vlivy na kočičí oční motoneurony". Výzkum mozku. 20 (1): 27–40. doi:10.1016/0006-8993(74)90890-7. PMID 4373140.
- ^ Ulanovsky & Moss, 2008
- ^ Valentine & Moss, 1997
- ^ Westby, G. W. M .; Keay, K. A .; Redgrave, P .; Dean, P .; Bannister, M. (srpen 1990). „Výstupní cesty zprostředkující přístup a vyhýbání se krysím colliculus mají různé senzorické vlastnosti“. Experimentální výzkum mozku. 81 (3): 626–638. doi:10.1007 / BF02423513. PMID 2226694.
- ^ Cohen, J. D .; Castro-Alamancos, M. A. (23. června 2010). „Neurální korelace aktivního chování při vyhýbání se u superior colliculus“. Journal of Neuroscience. 30 (25): 8502–8511. doi:10.1523 / JNEUROSCI.1497-10.2010. PMC 2905738. PMID 20573897.
- ^ Caltharp SA, Pira CU, Mishima N, Youngdale EN, McNeill DS, Liwnicz BH, Oberg KC (2007). „Indukce a lokalizace NOGO-A během vývoje mozku kuřat naznačují roli odlišnou od inhibice růstu neuritů“. BMC Dev. Biol. 7 (1): 32. doi:10.1186 / 1471-213X-7-32. PMC 1865376. PMID 17433109.
- ^ Beart, Phillip (1976). „Vyhodnocení L-glutamátu jako vysílače uvolněného z terminálů zrakového nervu holuba“. Výzkum mozku. 110 (1): 99–114. doi:10.1016/0006-8993(76)90211-0. PMID 6128.
- ^ Avitan, L .; Pujic, Z .; Mölter, J .; Van De Poll, M .; Sun, B .; Teng, H .; Amor, R .; Scott, E. K.; Goodhill, G.J. (2017). „Spontánní aktivita v Zebrafish Tectum se během vývoje reorganizuje a je ovlivněna vizuální zkušeností“. Aktuální biologie. 27 (16): 2407–2419. doi:10.1016 / j.cub.2017.06.056. PMID 28781054.
- ^ Dunn, Timothy W; et al. (2016). „Neuronové obvody podkladové k vizuálně vyvolaným únikům v Larval Zebrafish“. Neuron. 89 (3): 613–28. doi:10.1016 / j.neuron.2015.12.021. PMC 4742414. PMID 26804997.
- ^ Heap, LA; Vanwalleghem, GC; Thompson, AW; Favre-Bulle, I; Rubinsztein-Dunlop, H; Scott, EK (2018). „Hypotalamické projekce optického tektum v Larval Zebrafish“. Přední Neuroanat. 11: 135. doi:10.3389 / fnana.2017.00135. PMC 5777135. PMID 29403362.
- ^ Roeser, Tobias (2003). „Vizuomotorické chování u larválních zebrafish po laserové ablaci optického tektu řízené GFP“. Journal of Neuroscience. 23 (9): 3726–3734. doi:10.1523 / JNEUROSCI.23-09-03726.2003.
- ^ Barker, Alison (2015). „Senzomotorické rozhodování v Zebrafish Tectum“. Aktuální biologie. 25 (21): 2804–2814. doi:10.1016 / j.cub.2015.09.055. PMID 26592341.
- ^ Maximino, 2008
- ^ A b C Northcutt, 2002
- ^ Henriques, Pedro M .; Rahman, Niloy; Jackson, Samuel E .; Bianco, Isaac H. (3. června 2019). „Nucleus Isthmi je vyžadován k udržení pronásledování cíle během vizuálně vedeného lovu kořisti“. Aktuální biologie. 29 (11): 1771–1786.e5. doi:10.1016 / j.cub.2019.04.064. PMC 6557330. PMID 31104935.
- ^ Hartline a kol., 1978
- ^ Grillner, 2003
- ^ Saitoh et al., 2007
- ^ Wylie, DR; Gutierrez-Ibanez, C; Pakan, JM; Iwaniuk, AN (prosinec 2009). „Optické tektum ptáků: mapování naší cesty k porozumění vizuálnímu zpracování“. Kanadský žurnál experimentální psychologie. 63 (4): 328–38. doi:10.1037 / a0016826. PMID 20025392. S2CID 2712427.
Reference
- Chevalier, G; Mana S (2000). „Voštinová struktura mezilehlých vrstev krysy superior colliculus, s dalšími pozorováními u několika dalších savců: AChE vzorování“. J Comp Neurol. 419 (2): 137–53. doi:10.1002 / (SICI) 1096-9861 (20000403) 419: 2 <137 :: AID-CNE1> 3.0.CO; 2-6. PMID 10722995.
- Dash, S; Yang X; Wang H; Crawford JD (2015). „Průběžná aktualizace visuospatiální paměti v superior colliculus během pomalých pohybů očí“. Curr Biol. 25 (3): 267–74. doi:10.1016 / j.cub.2014.11.064. PMID 25601549.
- Dean, P; Redgrave P; Westby GW (1989). „Událost nebo nouzová situace? Dva systémy odezvy v savčím superior colliculus“. Trendy Neurosci. 12 (4): 137–47. doi:10.1016/0166-2236(89)90052-0. PMID 2470171.
- Gandhi, NJ; Katani HA (2011). „Motorické funkce Superior Colliculus“. Annu Rev Neurosci. 34: 205–231. doi:10.1146 / annurev-neuro-061010-113728. PMC 3641825. PMID 21456962.
- Grillner, S (2003). "Motorová infrastruktura: od iontových kanálů k neuronovým sítím". Recenze přírody Neurovědy. 4 (7): 573–86. doi:10.1038 / nrn1137. PMID 12838332.
- Hartline, PH; Kass L; Loop MS (1978). "Sloučení modalit v optickém tektu: infračervená a vizuální integrace u chřestýšů". Věda. 199 (4334): 1225–9. Bibcode:1978Sci ... 199.1225H. doi:10.1126 / science.628839. PMID 628839.
- Huerta, MF; Harting JK (1984). Vanegas H (ed.). Srovnávací neurologie optického tektum. New York: Plenum Press. 687–773. ISBN 978-0-306-41236-3.
- Illing, R-B (1996). Mozaiková architektura superior colliculus. Prog Brain Res. Pokrok ve výzkumu mozku. 112. str. 17–34. doi:10.1016 / S0079-6123 (08) 63318-X. ISBN 9780444823472. PMID 8979818.
- King, AJ; Schnupp JWH; Carlile S; Smith AL; Thompson ID (1996). The development of topographically-aligned maps of visual and auditory space in the superior colliculus. Prog Brain Res. Pokrok ve výzkumu mozku. 112. str. 335–350. doi:10.1016/S0079-6123(08)63340-3. ISBN 9780444823472. PMID 8979840.
- Klier, EM; Wang H; Crawford JD (2001). "The superior colliculus encodes gaze commands in retinal coordinates" (PDF). Nat Neurosci. 4 (6): 627–32. doi:10.1038/88450. PMID 11369944.
- Klier, E; Wang H; Crawford D (2003). "Three-dimensional eye-head coordination is implemented downstream from the superior colliculus". J. Neurophysiol. 89 (5): 2839–53. CiteSeerX 10.1.1.548.1312. doi:10.1152/jn.00763.2002. PMID 12740415.
- Krauzlis, R; Liston D; Carello C (2004). "Target selection and the superior colliculus: goals, choices and hypotheses". Vision Res. 44 (12): 1445–51. doi:10.1016/j.visres.2004.01.005. PMID 15066403.
- Kustov, A; Robinson D (1996). "Shared neural control of attentional shifts and eye movements". Příroda. 384 (6604): 74–77. Bibcode:1996Natur.384...74K. doi:10.1038/384074a0. PMID 8900281.
- Lane, RH; Allman JM; Kaas JH; Miezin FM (1973). "The visuotopic organization of the superior colliculus of the owl monkey (Aotus trivirgatus) and the bush baby (Galago senegalensis)". Brain Res. 60 (2): 335–49. doi:10.1016/0006-8993(73)90794-4. PMID 4202853.
- Lunenburger, L; Kleiser R; Stuphorn V; Miller LE; Hoffmann KP (2001). A possible role of the superior colliculus in eye–hand coordination. Prog Brain Res. Pokrok ve výzkumu mozku. 134. pp. 109–25. doi:10.1016/S0079-6123(01)34009-8. ISBN 9780444505866. PMID 11702538.
- Mana, S; Chevalier G (2001). "Honeycomb-like structure of the intermediate layers of the rat superior colliculus: afferent and efferent connections". Neurovědy. 103 (3): 673–93. doi:10.1016/S0306-4522(01)00026-4. PMID 11274787.
- Maximino, C; Soares, Daphne (2008). Soares, Daphne (ed.). "Evolutionary changes in the complexity of the tectum of nontetrapods: a cladistic approach". PLOS ONE. 3 (10): e385. Bibcode:2008PLoSO...3.3582M. doi:10.1371/journal.pone.0003582. PMC 2571994. PMID 18974789.
- Munoz, DP; Pélisson D; Guitton D (1991). "Movement of activity on the superior colliculus motor map during gaze shifts" (PDF). Věda. 251 (4999): 1358–60. doi:10.1126/science.2003221. PMID 2003221.
- Northcutt, RG (2002). "Understanding vertebrate brain evolution". Integr Comp Biol. 42 (4): 743–6. doi:10.1093 / icb / 42.4.743. PMID 21708771.
- Pettigrew, JD (1986). "Flying primates? Megabats have the advanced pathway from eye to midbrain". Věda. 231 (4743): 1304–6. Bibcode:1986Sci...231.1304P. doi:10.1126/science.3945827. PMID 3945827. S2CID 16582493.
- Pierrot-Deseilligny, C; Müri RM; Ploner CJ; Gaymard B; Rivaud-Péchoux S (2003). Cortical control of ocular saccades in humans: a model for motricity. Prog Brain Res. Pokrok ve výzkumu mozku. 142. s. 3–17. doi:10.1016/S0079-6123(03)42003-7. ISBN 9780444509772. PMID 12693251.
- Saitoh, K; Ménard A; Grillner S (2007). "Tectal control of locomotion, steering, and eye movements in lamprey". J. Neurophysiol. 97 (4): 3093–108. doi:10.1152/jn.00639.2006. PMID 17303814. S2CID 5711513.
- Soetedjo, R; Kaneko CR; Fuchs AF (2002). "Evidence against a moving hill in the superior colliculus during saccadic eye movements in the monkey". J. Neurophysiol. 87 (6): 2778–89. doi:10.1152/jn.2002.87.6.2778. PMID 12037180. S2CID 18294502.
- Sparks, DL (1999). "Conceptual issues related to the role of the superior colliculus in the control of gaze". Aktuální názor v neurobiologii. 9 (6): 698–707. doi:10.1016/S0959-4388(99)00039-2. PMID 10607648.
- Sparks, DL; Gandhi NJ (2003). Single-cell signals: an oculomotor perspective. Prog Brain Res. Pokrok ve výzkumu mozku. 142. str. 35–53. doi:10.1016/S0079-6123(03)42005-0. ISBN 9780444509772. PMID 12693253.
- Sprague, JM (1996). Neural mechanisms of visual orienting responses. Prog Brain Res. Pokrok ve výzkumu mozku. 112. s. 1–15. doi:10.1016/S0079-6123(08)63317-8. ISBN 9780444823472. PMID 8979817.
- Stein, BE; Clamman HP (1981). "Control of pinna movements and sensorimotor register in cat superior colliculus". Brain Behav Evol. 19 (3–4): 180–192. doi:10.1159/000121641. PMID 7326575.
- Ulanovsky, N; Moss CF (2008). "What the bat's voice tells the bat's brain". PNAS. 105 (25): 8491–98. Bibcode:2008PNAS..105.8491U. doi:10.1073/pnas.0703550105. PMC 2438418. PMID 18562301.
- Valentine, D; Moss CF (1997). "Spatially selective auditory responses in the superior colliculus of the echolocating bat". J. Neurosci. 17 (5): 1720–33. doi:10.1523/JNEUROSCI.17-05-01720.1997. PMC 6573370. PMID 9030631.
- Wallace, MT; Meredith MA; Stein BE (1998). "Multisensory integration in the superior colliculus of the alert cat". J. Neurophysiol. 80 (2): 1006–10. doi:10.1152/jn.1998.80.2.1006. PMID 9705489.
externí odkazy
| Kontrolní úřad |
|---|