Lidský herpesvirus 6 - Human herpesvirus 6
| Lidský herpesvirus 6 | |
|---|---|
 | |
| Elektronový mikrofotografie jednoho z druhů HHV6 | |
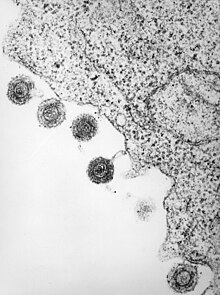 | |
| HHV-6 uvolňovaný z infikovaného lymfocytu | |
| Vědecká klasifikace | |
| (bez hodnocení): | Virus |
| Oblast: | Duplodnaviria |
| Království: | Heunggongvirae |
| Kmen: | Peploviricota |
| Třída: | Herviviricetes |
| Objednat: | Herpesvirales |
| Rodina: | Herpesviridae |
| Podčeleď: | Betaherpesvirinae |
| Rod: | Roseolovirus |
| Skupiny v ceně | |
| Kladisticky zahrnuty, ale tradičně vyloučeny taxony | |
Vše ostatní Roseolovirus spp .: | |
Lidský herpesvirus 6 (HHV-6) je společný souhrnný název pro lidský betaherpesvirus 6A (HHV-6A) a lidský betaherpesvirus 6B (HHV-6B). Tyto blízce příbuzné viry jsou dva z devíti herpesvirů, o nichž je známo, že jsou jejich primárním hostitelem lidé.[1]
HHV-6A a HHV-6B jsou dvouvláknové DNA viry uvnitř Betaherpesvirinae podčeleď a rodu Roseolovirus. HHV-6A a HHV-6B infikují téměř všechny lidské populace, které byly testovány.[2]
HHV-6A byl popsán jako další neurovirulentní,[3] a jako takový se častěji vyskytuje u pacientů s neurozánětlivými chorobami, jako je roztroušená skleróza.[4] Hladiny HHV-6 (a HHV-7) v mozku jsou také zvýšené u lidí s Alzheimerova choroba.[5]
Primární infekce HHV-6B je příčinou běžné dětské nemoci exanthema subitum (také známý jako roseola infantum nebo šestá nemoc). Přenáší se z dítěte na dítě. Je neobvyklé, že se dospělí nakazili touto chorobou, protože ji většina lidí měla ve školce a jakmile se nakazí, vytvoří se protilátka, která pomůže v budoucnu zabránit reinfekci. U příjemců transplantátu je navíc běžná reaktivace HHV-6B, která může způsobit několik klinických projevů, jako je encefalitida, potlačení kostní dřeně a pneumonitida.[6]
Při detekci HHV-6 se používá celá řada testů, z nichž některé tyto dva druhy nerozlišují.[7]
Dějiny
V průběhu roku 1986 Syed Zaki Salahuddin, Dharam Ablashi, a Robert Gallo kultivované mononukleární buňky periferní krve od pacientů s AIDS a lymfoproliferativní onemocnění. Byly dokumentovány krátkodobé velké refrakční buňky, které často obsahovaly intranukleární a / nebo intracytoplazmatická inkluzní tělíska. Elektronová mikroskopie odhalili nový virus, který nazvali Human B-Lymphotrophic Virus (HBLV).[8][9]
Krátce po svém objevu Ablashi a kol. popsáno pět buněčných linií, které mohou být infikovány nově objeveným HBLV. To zveřejnili HSB-2, konkrétní linie T-buněk, je vysoce náchylná k infekci. Průkopnický výzkum společnosti Ablashi byl zakončen návrhem na změnu názvu viru z HBLV na HHV-6 v souladu se zveřejněnou předběžnou klasifikací herpetických virů.[10][11]
O několik let později byl HHV-6 rozdělen na podtypy. Early research (1992) popsal dvě velmi podobné, přesto jedinečné varianty: HHV-6A a HHV-6B. Tento rozdíl byl zaručen díky jedinečným štěpením restrikční endonukleázy, reakcím monoklonálních protilátek,[12] a růstové vzorce.[13]
HHV-6A zahrnuje několik kmenů odvozených od dospělých a jeho spektrum onemocnění není dobře definováno, i když někteří si myslí, že je více neurovirulentní.[14][15] HHV-6B je běžně detekován u dětí s roseola infantum, protože se jedná o etiologický prostředek pro tento stav. V rámci těchto dvou virů je 95% sekvenční homologie.[16]
V roce 2012 byly HHV-6A a HHV-6B oficiálně uznány jako odlišné druhy.[1]
Taxonomie
HHV-6A a HHV-6B byly uznány organizací Mezinárodní výbor pro taxonomii virů (ICTV) jako odlišné druhy v roce 2012. Mezi lidské roseoloviry patří HHV-6A, HHV-6B a HHV-7.[1]
Herpesvirus byla založena jako rod v roce 1971 v první zprávě ICTV. Tento rod sestával z 23 virů ve 4 skupinách.[17] V roce 1976 byla vydána druhá zpráva ICTV, ve které byl tento rod povýšen na rodinnou úroveň - herpetoviridae. Kvůli možné záměně s viry pocházejícími z plazů bylo ve třetí zprávě (1979) změněno příjmení na herpesviridae. V této zprávě rodina Herpesviridae byl rozdělen do 3 podskupin (alphaherpesvirinae, betaherpesvirinae a gammaherpesvirinae) a 5 nejmenovaných rodů; Jako členové rodiny bylo rozpoznáno 21 virů.[18]
V roce 2009 objednávka Herpesvirales byl vytvořen. To bylo nutné díky objevu, že herpetické viry ryb a měkkýšů jsou jen vzdáleně příbuzné virům ptáků a savců. Objednat Herpesvirales obsahuje tři rodiny, Herpesviridae, který obsahuje dlouho uznávané herpesviry savců, ptáků a plazů, plus dvě nové rodiny - rodinu Alloherpesviridae který zahrnuje herpetické viry kostnatých ryb a žab a rodiny Malacoherpesviridae který obsahuje viry měkkýšů.[19]
Od roku 2012 má tento řád v současné době 3 rodiny, 4 podskupiny (1 nepřiřazeno), 18 rodů (4 nepřiřazeno) a 97 druhů.[1]
Struktura
Průměr virionu HHV-6 je asi 2 000 angstromů.[9] Vnější část virionu se skládá z lipidové dvouvrstvé membrány, která obsahuje viry glykoproteiny a je odvozen od hostitele. Pod touto membránovou obálkou je a tegument který obklopuje icosahedral kapsid, složený ze 162 kapsomery. Ochranná kapsida HHV-6 obsahuje dvouvláknovou lineární DNA.
Během zrání virionů HHV-6 se lidské buněčné membrány používají k vytvoření obalů virových lipidů (jak je charakteristické pro všechny obalené viry). Během tohoto procesu využívá HHV-6 lipidové rafty, což jsou membránové mikrodomény obohacené o cholesterol, sfingolipidy, a glykosylfosfatidylinositol - ukotvené proteiny.[20] První badatelé měli podezření, že viriony HHV-6 v jádru dozrávají; někteří to dokonce nesprávně publikovali, protože zobecnili a aplikovali na HHV-6 to, co bylo známo o jiných virech. Výzkum zveřejněný v roce 2009 však naznačuje, že virus HHV-6 využívá pro sestavování vezikuly odvozené od trans-Golgiho sítě.[20]
Genom
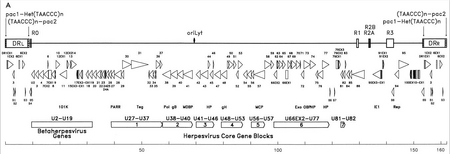
Genetický materiál HHV-6 se skládá z lineární (cirkulární během aktivní infekce), dvouvláknové DNA, která obsahuje původ replikace, dva 8–10 kb levé a pravé přímé opakované konce a jedinečný segment, který má 143–145 kB.[22]
Počátek replikace (v literatuře často označovaný jako „oriLyt“) je místo, kde začíná replikace DNA.[21] Přímé opakované konce (DRL a DRR) mají opakovanou sekvenci TTAGGG, identickou se sekvencí člověka telomery. Variabilita v počtu telomerických opakování je pozorována v rozmezí 15–180.[23][24] Tyto konce také obsahují štěpení pac-1 a pac-2 a obalové signály, které jsou mezi herpesviry zachovány.
Unikátní segment obsahuje sedm hlavních bloků jádrových genů (U27-U37, U38-U40, U41-U46, U48-U53, U56-U57, U66EX2-U77 a U81-U82),[21] což je také charakteristické pro herpesviry. Tyto konzervované geny kódují proteiny, které se účastní replikace, štěpení a balení virového genomu do zralého virionu.[23] Kromě toho kódují řadu imunomodulačních proteinů. Unikátní segment také obsahuje blok genů (U2-U19), které jsou konzervovány mezi HHV-6, HHV-7 a Cytomegaloviry (betaherpesviry). Řada jedinečných genů segmentů je spojena například s rodinou HCMV US22 (InterPro: IPR003360 ). V tabulce níže jsou uvedeny některé jejich známé vlastnosti.[21]
Geny
| Gen | Etapa | Vlastnosti |
|---|---|---|
| IE-A (IE1? U89?) | Okamžitě brzy | Část IE lokusu [25] - zhoršuje expresi genu interferonu, aby omezil vývoj buněčných antivirových opatření, upřednostňuje úspěšnou infekci - není v membráně - aktivuje virové DNA polymerázy, podílí se na replikaci klouzavého kruhu - exprese tohoto genu může být modulována mikro RNA [26] |
| IE-B | Okamžitě brzy | Část IE lokusu [25] Aktivuje virové DNA polymerázy, podílí se na replikaci klouzavého kruhu |
| DR1 | Genová rodina HCMV US22 | |
| DR6 | Genová rodina HCMV US22, transaktivátor, onkogen | |
| DR7 / U1 | SR doména, maligní transformační aktivita, se váže na p53 | |
| U2 | Genová rodina HCMV US22 - protein proteinu | |
| U3 | Homolog HCMV UL24, rodina genů HCMV US22, tegumentový protein - transaktivační aktivita [25] | |
| U4 | Rezistence na HCMV maribavir | |
| U7 | Genová rodina HCMV US22 | |
| U10 | rodina dUTPase | |
| U11 | Silně imunoreaktivní virionový protein [21] - antigenní tegumentový protein | |
| U12 | Receptor spojený s proteinem chemokinu G. | |
| U13 | CMV: Potlačí přepis US3 | |
| U14 | Váže a začleňuje p53 do virových částic - rodina genů HCMV UL25 - antigenní protein proteinu | |
| U15 | Genová rodina HCMV UL25 | |
| U17 | Rodina genů HCMV UL25 - tegumentový protein | |
| U18 | IE-B | Membránový glykoprotein |
| U19 | IE-B protein | Glykoprotein |
| U20 | Glykoprotein (specifický pro Roseolovirus) předpokládaná struktura imunoglobulinu | |
| U21 | Váže se na molekuly MHC-1 a brání buňkám prezentujícím antigen prezentovat peptidy HHV-6 - glykoprotein, snižuje hladinu HLA I (specifické pro Roseolovirus) | |
| U22 | Pozdní gen | Glykoprotein (chybí z HHV-7, specifické pro Roseolovirus) |
| U23 | Glykoprotein (specifický pro Roseolovirus ) | |
| U24 | Inhibuje správnou aktivaci T buněk, snižuje sekreci cytokinů v místě infekce - fosforylační cíl pro kinázy - glykoprotein M (gM) (specifický pro Roseolovirus ) | |
| U25 | Rodina genů HCMV UL22, tegumentový protein | |
| U26 | Předpokládaný mnohonásobný transmembránový protein | |
| U27 | Továrna na zpracování DNA polymerázy | |
| U28 | Velká podjednotka ribonukleotid reduktázy, tegumentový protein | |
| U29 | Sestavení kapsidy a zrání DNA | |
| U30 | Tegumentový protein | |
| U31 | Velký protein proteinu | |
| U32 | Kapsidový protein, hexon tipy | |
| U33 | Virionový protein | |
| U34 | Fosfoprotein asociovaný s membránou, primární obal | |
| U35 | Terminázová složka, obal DNA | |
| U36 | DNA balení | |
| U37 | Tegumentový protein, primární obal, fosfoprotein | |
| U38 | DNA polymeráza | |
| U39 (gB, gp116) | Glykoprotein | |
| U40 | Transport, montáž kapsidy | |
| U41 | Časný gen | Hlavní protein vázající DNA |
| U42 | Tegumentový protein, blok buněčného cyklu, transaktivátor | |
| U43 | DNA Komplex helikáza-primáza | |
| U44 | Tegumentový protein | |
| U45 | dUTPase | |
| U46 | Glykoprotein N, membránový protein | |
| U47 (gO, O) | Glykoprotein O, sdružený s lipidovými rafty, existuje ve dvou formách, gO-120K a gO-80K, a gO-80K obsahuje komplexní oligosacharidy vázané na N, které jsou inkorporovány do virových částic | |
| U48 (gH, gp100) | Glykoprotein gH, složka virionu, součást komplexu ligandu CD46 gQ1 / gQ2 / gL / gH, asociuje s lipidovými rafty | |
| U49 | Regulační protein spojený s viriony, fúzní protein | |
| U50 | DNA balení | |
| U51 | Časný gen | Chemokinový receptor spojený s G proteinem, který brání expresi, výrazně snižuje replikaci - zvyšuje intracelulární hladiny inositol fosfátu druhého posla, podporuje chemotaxi - časný gen, spolu s U41 a U69 [7] |
| U52 | ||
| U53 | Proteáza,[25] kapsidový montážní protein | |
| U54 | Tegumentový protein, transaktivátor virionu | |
| U55 | Role v syntéze RNA, dUTPáza | |
| U56 | Kapsidový protein | |
| U57 | Hlavní kapsidový protein | |
| U58 | ||
| U59 | Tegumentový protein | |
| U61 | ||
| U62 | ||
| U63 | ||
| U64 | DNA obal: tegumentový protein | |
| U65 | Tegumentový protein | |
| U66 | Terminázová složka | |
| U69 | Časný gen | Tegumentová proteinkináza (ganciklovirkináza) zapojená do replikace [25] |
| U70 | Alkalická exonukleáza | |
| U71 | Myristylovaný virionový protein | |
| U72 (gM) | Glykoprotein M. | |
| U73 | Protein vázající původ | |
| U74 | Komplex DNa helikáza-primáza | |
| U75 | Tegumentový protein | |
| U76 | DNA obal, virionový protein | |
| U77 | Komplex helikáza-primáza | |
| U79 | Aktivace transkripce | |
| U80 | Předpokládaná struktura imunoglobulinu | |
| U81 | Uracil-DNA glykosyláza | |
| U82 (gL, gp80) | Glykoprotein L, složka virionu, součást komplexu ligandu CD46 gQ1 / gQ2 / gL / gH, asociuje s lipidovými rafty | |
| U83 | Vylučovaný chemotaktický (chemoatraktant) glykoprotein, váže se na receptory chemokinů, rekrutuje hostitelské buňky, které vylučují chemokiny specifické pro U51 | |
| U85 | Glykoprotein (specifický pro Roseolovirus) | |
| U86 | IE-2 | Transaktivátor IE-2 |
| U88 | IE-A | |
| U90 | IE-A (IE 1) | Transaktivátor |
| U91 | IE-A, glykoprotein | |
| U94 | Latence (okamžitý časný nebo časný gen) | Podílí se na transkripční represi lytických genů - pomáhá při specifické integraci HHV-6A / HHV-6B do telomer - vysoce exprimován během latence - parvovirus rep homolog (chybí v HHV-7) |
| U95 | Rodina genů CMV US22 - kolokalizuje a interaguje s mitochondriálním proteinem GRIM-19, základní složkou systému oxidační fosforylace [7] - váže se na nukleární faktor-kappa B (NF-κB), jehož deregulace má přispívat k rakovině [14] | |
| U100 (Gp82-105) | Pozdní gen | Glykoprotein Q, složka virionu, se asociuje s lipidovými rafty |
| gQ1 | Glykoprotein, komplexy s gH a gL za vzniku virového ligandu k receptoru CD46 - modifikovaný N-glykosylací - exprimovaný ve dvou různých formách: forma 80-kDa (gQ1-80K) a forma 74-kDa (gQ1-74K) - pouze gQ1-80K, ale ne gQ1-74K, tvoří komplex ligandu CD46 s gQ2, gH a gL [27] Spojuje se s lipidovými rafty. | |
| gM1 | Gangliozid specifický pro lipidový vor, začleněný do virionu | |
| gQ2 | Glykoprotein, tvoří komplex gH / gL / gQ1 / gQ2, část receptorového ligandu - nezbytný pro růst virů, asociuje se s lipidovými rafty - existuje ve dvou formách: gQ2-34K a gQ2-37K | |
| Mikro RNA | hhv6b-miR-Ro6-1, -Ro6-2, -Ro6-3 a -Ro6-4. Může regulovat časný přepis | |
| P100 aka p101 | Imunogenní, složka tegumentu | |
| ORF-1 (DR7) | Váže a inhibuje transkripční aktivitu p53 - může transformovat lidské epidermální keratinocyty a buňky NIH 3T3 in vitro - buňky exprimující protein ORF-1 produkují fibrosarkomy po injekci do nahých myší |
Virální vstup
HHV-6 receptor
Když extracelulární virion HHV-6 narazí na lidské buňky, narazí na diferenciační klastr lidského receptorového proteinu 46 (CD46 ), který hraje roli při regulaci systému komplementu. Výsledkem je, že protein CD46 má jednu variabilní oblast alternativní sestřih. Jako takové existuje alespoň čtrnáct izoforem CD46, z nichž všechny vážou HHV-6a.[28]
Extracelulární oblast CD46 obsahuje čtyři krátké konsenzuální opakování přibližně 60 aminokyselin, které se skládají do kompaktní formy beta-barel doména obklopená pružnými smyčkami.[23] Jak bylo prokázáno pro CD46 s jinými ligandy, proteinová struktura CD46 se linearizuje po navázání HHV-6. Zatímco jejich přesná interakce ještě nebyla stanovena, byla prokázána druhá a třetí SCR doména, jak je požadováno pro vazbu na receptor HHV-6 a buněčný vstup.
Ligand receptoru HHV-6
Mori a kol. nejprve identifikoval genový produkt gQ1, glykoprotein jedinečný pro HHV-6, a zjistil, že tvoří komplex s glykoproteiny gH a gL.[12][29] Věřili, že tento heterotrimerový komplex slouží jako virový ligand pro CD46.[22] Brzy poté byl identifikován další glykoprotein s názvem gQ2 a bylo zjištěno, že je součástí ligandového komplexu gH / gL / gQ1, tvořící heterotetramer který byl pozitivně identifikován jako virový CD46 ligand.[29] Přesný proces vstupu není dosud dobře znám.
Slinné žlázy
Slinné žlázy byly popsány jako rezervoár in vivo pro infekci HHV-6.[23]
Leukocyty
Výzkumní pracovníci[30] prokázali, že T buňky jsou vysoce infikovatelné HHV-6.
Nervový systém
V průběhu roku 2011 vědci z Národní institut zdraví se pokusil objasnit tehdy neznámou metodu, při níž HHV-6a získává vstup do nervového systému. Jako takové pitvali mozky přibližně 150 subjektů. Když byly různé anatomické oblasti testovány na virovou zátěž, čichový Bylo zjištěno, že tkáně mají nejvyšší obsah HHV-6. Došli k závěru, že tyto tkáně jsou vstupním bodem pro HHV-6a.[16]
Výsledky výše jsou v souladu s výsledky předchozích studií, které zahrnovaly HSV-1 (a řada dalších virů), které se také šíří do CNS prostřednictvím čichové tkáně.[31]
Vědci to také předpokládali čichové obklopující buňky (OECs), skupina specializovaných gliové buňky nalezené v nosní dutině, mohou hrát roli v infekčnosti HHV-6.[16] Měli podezření na tuto asociaci v důsledku toho, že OEC mají vlastnosti podobné těm z astrocyty, jiný typ gliové buňky, který byl dříve identifikován jako náchylný k infekci HHV-6.[32] Výzkum pokračoval infikováním OEC in vitro oběma typy HHV-6. Nakonec pouze OEC, ve kterých byl použit HHV-6a, byly pozitivně testovány na známky de novo virové syntézy, jak je také charakteristické pro astrocyty.[32]
Buněčná aktivita
Jakmile jste uvnitř, byly popsány dva výsledky: aktivní a neaktivní infekce.
Aktivní infekce
Aktivní infekce zahrnují lineární genom dsDNA cirkulující od začátku do konce kovalentní vazby. Tento proces byl poprvé nahlášen pro virus herpes simplex.[24] Po cirkularizaci začne HHV-6 exprimovat geny známé jako „okamžité rané“. Předpokládá se, že tyto genové produkty jsou transkripční aktivátory[7] a mohou být regulovány expresí viru mikro RNA.[26] Následně dochází k následné expresi "časných genů" a aktivuje se například virová DNA polymerázy. Rané geny jsou také zapojeny do replikace postupného kruhu to následuje.[23]
Replikace HHV-6 vede k tvorbě dobyvatelé, což jsou dlouhé molekuly, které obsahují několik opakování sekvence DNA.[33] Tyto dlouhé concatemery jsou poté štěpeny mezi oblastmi pac-1 a pac-2 pomocí ribozymy pro balení genomu do jednotlivých virionů.[24]
Neaktivní infekce
Ne všechny nově infikované buňky začnou replikaci válcovaného kruhu. Ve skutečnosti, opar pochází z řeckého slova herpein, což znamená „plazit se“. Herpesviry se „plazí“ v tom, že mohou vstoupit do latentního stádia a nečinně infikovat svého lidského hostitele. Od svého objevu v roce 1993 byl tento fenomén nalezen mezi všemi betaherpesviry.[34]
Jiné betaherpesviry stanoví latenci jako nukleární epizom, což je kruhová molekula DNA (analogicky k plazmidy ). U HHV-6 se předpokládá, že latence nastává výlučně integrací virových telomerních repeticí do člověka subtelomerické oblasti.[15] Pouze jeden další virus, Marekova nemoc virus je známo, že tímto způsobem dosahuje latence.[7] Tento jev je možný v důsledku telomerických repeticí nalezených v přímých opakovaných koncích genomu HHV-6.
Správný přímý terminál opakování se integruje do 5 až 41 opakování lidských telomer a přednostně to dělá do proximálního konce[35] chromozomů 9, 17, 18, 19 a 22, ale také se příležitostně vyskytovala v chromozomech 10 a 11.[33] Je podezření, že téměř 70 milionů jedinců nese chromozomálně integrovaný HHV-6.[15][33]
Řada genů exprimovaných HHV-6 je jedinečná pro fázi neaktivní latence. Tyto geny zahrnují udržování genomu a zabránění destrukci hostitelské buňky.[35] Například se předpokládá, že protein U94 potlačuje geny, které se účastní buněčné lýzy (apoptózy), a také může pomáhat při telomerické integraci.[23] Jakmile je virus uložen v lidských telomerách, je přerušovaně reaktivován.[35]
Reaktivace
Konkrétní spouštěče reaktivace nejsou dobře známy. Někteří vědci naznačují, že by mohlo dojít ke zranění, fyzickému nebo emočnímu stresu a hormonální nerovnováze.[36]
Vědci v průběhu roku 2011 zjistili, že reaktivaci může pozitivně spustit in vitro histon deacetyláza inhibitory. Jakmile začne reaktivace, je zahájen proces válcování kruhu a jsou vytvořeny concatemery, jak je popsáno výše.[23]
Interakce
Lidský herpesvirus 6 žije primárně na lidech a zatímco varianty viru mohou způsobit mírná až smrtelná onemocnění, mohou žít komensálně na svém hostiteli.[13] Bylo prokázáno, že HHV-6 podporuje progresi HIV-1 při koinfekci v T buňkách.[37] HHV-6 zvyšuje expresi primárního HIV receptoru CD4, čímž rozšiřuje rozsah buněk citlivých na HIV. Několik studií také ukázalo, že infekce HHV-6 zvyšuje produkci zánětlivých cytokinů, které zvyšují expresi HIV-1 in vitro, jako je TNF-alfa,[38] IL-1 beta a IL-8.[39] Novější studie in vivo ukazuje, že koinfekce HHV-6A dramaticky urychluje postup od HIV k AIDS u makaků pigtailovaných.[40]
Bylo také prokázáno, že HHV-6 transaktivuje virus Epstein-Barr.[31]
Epidemiologie
Stáří
Lidé získávají virus v raném věku, někteří již ve věku méně než jeden měsíc. Primární infekce HHV-6 tvoří až 20% návštěv dětských pohotovostních sálů s horečkou ve Spojených státech[41][42] a jsou spojeny s několika závažnějšími komplikacemi, jako např encefalitida, lymfadenopatie, myokarditida a myelosuprese. Prevalence viru v těle se zvyšuje s věkem (míra infekce je nejvyšší u kojenců ve věku od 6 do 12 měsíců) a předpokládá se, že je to způsobeno ztrátou mateřských protilátek u dítěte, které ho chrání před infekcemi .[13]
Existují nesrovnalosti v korelacích mezi věkem a séropozitivitou: Podle některých zpráv došlo k poklesu o séropozitivita s nárůstem věku, zatímco některé nenaznačují žádný významný pokles a jiné uvádějí zvýšenou míru séropozivity u osob ve věku 62 let a starších. Po primární infekci se ve slinných žlázách stanoví latence, hematopoetické kmenové buňky a další buňky a existuje po celou dobu životnosti hostitele.
Zeměpisné rozdělení
Je známo, že virus je rozšířen po celém světě. Míra infekce HHV-6 64-83% ve věku 13 měsíců byla hlášena pro země včetně USA, Velké Británie, Japonska a Tchaj-wanu.[13][43] Studie zjistily, že séroprevalence se pohybuje „od přibližně 39 do 80% u etnicky různorodých dospělých populací z Tanzanie, Malajsie, Thajska a Brazílie“.[13] Neexistují žádné významné rozdíly mezi etnickými skupinami žijícími ve stejné zeměpisné poloze nebo mezi pohlavími. Zatímco HHV-6B je přítomen téměř ve všech světových populacích, zdá se, že HHV-6A je méně častý v Japonsku, Severní Americe a Evropě.[13]
Přenos
Předpokládá se, že k přenosu dochází nejčastěji vylučováním virových částic do slin. Jak HHV-6B, tak HHV-7 se nacházejí v lidských slinách, přičemž první z nich je na nižší frekvenci. Studie uvádějí různé míry prevalence HHV-6 ve slinách (mezi 3–90%),[13] a také popsali slinné žlázy jako zásobník in vivo pro HHV-6. Virus infikuje slinné žlázy, stanoví latenci a pravidelně se znovu aktivuje, aby se infekce rozšířila na další hostitele.[23]
Vertikální přenos byl také popsán a vyskytuje se u přibližně 1% porodů ve Spojených státech.[7][44] Tato forma je snadno identifikovatelná, protože virový genom je obsažen v každé buňce infikovaného jedince.
Diagnóza
Diagnóza infekce HHV-6 se provádí jak sérologickými, tak přímými metodami. Nejvýznamnější technikou je kvantifikace virové DNA v krvi, jiných tělních tekutinách a orgánech pomocí real-time PCR.[45]
Klinický význam
Klasická prezentace primární infekce HHV-6b je jako exanthema subitum (ES) nebo „roseola“ s vysokou teplotou následovanou vyrážkou. Jedna studie (1997) však naznačila, že vyrážka není charakteristickým rysem infekce HHV-6, s výskytem podobným infekcím jiným než HHV-6 (10–20% febrilních dětí v obou skupinách). Infekce HHV-6 se častěji vyskytují při vysokých teplotách (nad 40 ° C), rychlostí přibližně dvou třetin ve srovnání s méně než polovinou u pacientů bez HHV-6. Podobně významné rozdíly byly pozorovány v malátnosti, podrážděnosti a zánětu bubínku.[13]
Primární infekce u dospělých bývá závažnější.[13]
Diagnóza viru, zejména HHV-6B, je pro pacienta zásadní kvůli nepříznivým účinkům infekce. Příznaky, které poukazují na tuto infekci, jako jsou vyrážky, zůstávají bez povšimnutí u pacientů, kteří dostávají antibiotika, protože mohou být nesprávně interpretovány jako vedlejší účinek léku.[13] Je známo, že HHV-6B je spojován s dětským onemocněním roseola infantum, stejně jako s jinými nemocemi způsobenými infekcí. Patří mezi ně hepatitida, febrilní křeče a encefalitida. Děti, které trpí exanthema subitum, způsobené infekcí HHV-6B, pociťují horečky trvající 3 až 5 dní; vyrážky na trupu, krku a obličeji; a někdy febrilní křeče, příznaky však nejsou vždy přítomny společně. Primární infekce u dospělých jsou vzácné, protože většina případů je u dětí. Pokud k infekci dojde poprvé u dospělého, mohou být příznaky závažné.
Virus se pravidelně znovu aktivuje ze svého latentního stavu, přičemž DNA HHV-6 je detekovatelná u 20–25% zdravých dospělých ve Spojených státech. V imunokompetentním prostředí jsou tyto reaktivace často asymptomatické, ale u jedinců s potlačenou imunitou mohou nastat závažné komplikace. Reaktivace HHV-6 způsobuje závažné onemocnění u transplantace příjemci a může vést k odmítnutí štěpu, často ve spojení s jinými betaherpesviridae. Stejně tak v HIV / AIDS, HHV-6 reaktivace způsobují diseminované infekce vedoucí k onemocnění koncových orgánů a smrti. I když je až 100% populace vystaveno (séropozitivně) HHV-6, nejvíce ve věku 3 let, existují vzácné případy primárních infekcí u dospělých. Ve Spojených státech byly tyto spojeny spíše s HHV-6a, o kterém se předpokládá, že je více patogenní a neurotropní a byla spojena s několika centrální nervový systém související poruchy.
HHV-6 byl hlášen u roztroušená skleróza pacientů[46] a je implikován jako kofaktor u několika dalších onemocnění, včetně syndrom chronické únavy,[47] fibromyalgie, AIDS,[48] optická neuritida, rakovina, a epilepsie spánkového laloku.[49]
Roztroušená skleróza
Roztroušená skleróza (MS) je autoimunitní a zánětlivá porucha nervového systému, která vede k demyelinaci axonů v mozku a míše. Historie MS v kontextu HHV-6 začala v roce 1995, kdy Peter Challoner, vědec v PathoGenesis Corporation z Seattle, začali hledat nehumánní genetické sekvence v mozku pacientů s MS. Našel uvnitř neobvykle vysokou expresi HHV-6 DNA oligodendrocyty. Také si všiml vyšší koncentrace infikovaných buněk v oblastech, kde došlo k demyelinaci.[50] Jeho výzkum byl pravděpodobně první publikovanou studií naznačující souvislost mezi HHV-6 a MS.
Epidemiologické údaje
Prevalence MS se zvyšuje v populacích, protože jsou dále od EU Rovník.[51][52] Incidence je u narozených třikrát vyšší 42 stupňů severní šířky a vyšší než u narozených 37 stupňů na sever a níže. U osob je také méně pravděpodobné, že se u nich MS objeví jako dospělí, pokud jejich dětství bylo stráveno v oblasti s nízkým výskytem. Možnost vzniku původce infekce ve spojení s MS byla hodnocena optikou těchto epidemiologických nálezů.
K vysvětlení výše uvedených údajů byly navrženy dvě hypotézy.[53] První je známá jako hypotéza poliomyelitidy a naznačuje, že infekce v mladém věku poskytuje imunitu, ale infekce dospělých zvyšuje riziko RS. Druhá je známá jako hypotéza Prevalence a naznačuje, že MS je způsobena patogenem, který je častější v oblastech s vysokou mírou MS. Tento patogen by byl rozšířený a způsoboval u většiny jedinců asymptomatickou (latentní) infekci. Pouze zřídka a roky po primární infekci způsobuje toto hypotetické činidlo neurologické příznaky MS. Třetí hypotéza v podstatě kombinuje tyto dvě a také navrhuje zapojení více patogenů. Třetí může nejlépe platit pro epidemiologická data.[53][54]
Možné virové postižení
Za zmínku stojí také paradox viru Epstein – Barr (EBV), protože se uvádí, že HHV-6 transaktivuje EBV.[31] Jednotlivci mají 10krát menší riziko MS, pokud jsou séronegativní na EBV. Avšak mezi jedinci, kteří jsou pozitivní, jsou ti, kteří získají infekci EBV později v životě, trojnásobně větší riziko MS.
Výzkum naznačuje, že virové infekce mohou být vázány ještě blíže k MS. Protilátky EBV u zdravých jedinců zůstávají konstantní, zatímco hladiny protilátek u jedinců, u kterých se později rozvine MS, se začínají zvyšovat a plató mezi 20. a 30. rokem věku, bez ohledu na věk nástupu.
Specifičtěji pro HHV-6 vědci v roce 2004 zjistili, že počáteční stadia MS jsou spojena s vysokými hladinami aktivního viru.[55] Brzy poté vědci zjistili, že hladiny aktivního HHV-6 jsou také zvýšené během relapsů / exacerbací MS.[4]
Vědci prokázali, že hladiny HHV-6 IgG1 a IgM protilátek jsou u pacientů s MS ve srovnání s kontrolami zvýšené.[23] Výzkum publikovaný v roce 2014 ve skutečnosti zjistil, že zvýšení titrů anti-HHV-6A / B IgG a IgM predikují relaps MS.[56]
Výše uvedená analýza epidemiologických, sérologických a imunologických údajů podporuje asociaci mezi infekčním agens a MS. Přesný mechanismus možného virového ovlivnění projevu RS je však méně jasný. Přesto bylo navrženo několik mechanismů: molekulární mimikry, fosforylační dráhy a cytokiny.[16][57][58][59][60]
Molekulární mimikry
První studie, která specificky zkoumala demyelinaci související s HHV-6, se objevila v literatuře v průběhu roku 1996, kdy se u dříve zdravého 19měsíčního dítěte vyvinula akutní encefalopatie. Úrovně základní protein myelinu v jeho mozkomíšní mok, což naznačuje, že došlo k demyelinaci.[57] Tento odkaz byl téměř zapomenut, až o čtyři roky později, kdy byla zveřejněna studie související s MS, ukazující prevalenci HHV-6 90% mezi demyelinizovanými mozkovými tkáněmi. Ve srovnání s tím virus obsahovalo pouze 13% mozkových tkání bez onemocnění.[61]
Hypotéza molekulární mimikry, ve které T buňky si v podstatě pletou virový protein HHV-6 s myelinovým bazickým proteinem, poprvé se objevily kolem tohoto času. Na počátku vývoje této hypotézy (2002) italští vědci použili variantu HHV-6a spolu s bovinním myelinovým bazickým proteinem ke generování zkříženě reaktivních T buněčných linií. Ty byly porovnány s T buňkami jedinců s MS, stejně jako s kontrolami, a nebyl zjištěn žádný významný rozdíl mezi těmito dvěma. Jejich rané výzkumy naznačovaly, že molekulární mimikry nemusí být mechanismem, který je součástí MS.[58]
O několik měsíců později vědci ve Spojených státech vytvořili syntetický peptid se sekvencí identickou se sekvencí peptidu HHV-6. Byli schopni ukázat, že T buňky byly aktivovány tímto peptidem. Tyto aktivované T buňky také rozpoznaly a zahájily imunitní reakci proti synteticky vytvořené peptidové sekvenci, která je identická s částí lidského základního proteinu myelinu. Během svého výzkumu zjistili, že hladiny těchto zkříženě reagujících T buněk jsou významně zvýšené u pacientů s MS.[59] Jejich výzkum byl zakončen tvrzením, že HHV-6 může být skutečně původcem RS.
Následovalo několik podobných studií. Studie z října 2014 podpořila úlohu dlouhodobé infekce HHV-6 demyelinizací u progresivních neurologických onemocnění.[62]
Fosforylační dráhy
Myelinový základní protein (MBP) pravidelně vyměňuje fosfátové skupiny s prostředím a jeho schopnost tak činit má důsledky pro správné myelinová vrstva integrita. Přesněji, dva threoninové zbytky na MBP byly identifikovány jako fosforylační cíle glykogen syntáza kináza a mitogenem aktivovaná protein kináza. Jejich působení na MBP se říká, že pomáhá v jeho schopnosti polymerovat a svazovat myelin. Fosforylovaný MBP je také odolnější vůči několika proteázám.[60]
U jedinců s MS bylo zjištěno, že tyto cílové threoniny jsou fosforylovány méně často. Ve skutečnosti HHV-6 produkuje transmembránový protein, známý jako U24, který je také fosforylačním cílem výše zmíněných kináz. Naše kinázy působí na protein HHV-6 díky sdílené sekvenci sedmi aminokyselin (MBP92–104= IVTPRTPPPSQGK; U241–13= MDPPRTPPPSYSE). Ve výsledku nemusí u jedinců s aktivní infekcí HHV-6 docházet k zásadním posttranslačním úpravám MBP.[60]
- Roztroušená skleróza - přímé poškození buněk a změněné cytokiny
Bylo prokázáno, že HHV-6 infikuje čichové obklopující buňky (OEC). OEC byly důkladně vyšetřovány ve vztahu k poranění míchy, Amyotrofní laterální skleróza a další neurodegenerativní onemocnění. Vědci naznačují, že tyto buňky mají jedinečnou schopnost remyelinizovat poškozené neurony.[16]
Některé z genů exprimovaných HHV-6 manipulují s různými úrovněmi hostitele cytokiny (viz část o genových produktech). Například infikované buňky mají zvýšenou hladinu interleukin-8, o kterém se předpokládá, že indukuje MMP-9 represe. U jedinců s MS byly zjištěny zvýšené hladiny MMP-9.[63]
Reaktivace HHV-6 se také podílí na exacerbaci MS prostřednictvím posunu v podskupinách Th lymfocytů.[64]
Syndrom chronické únavy
Syndrom chronické únavy (CFS) je vysilující nemoc,[65] jehož příčina není známa. Pacienti s CFS mají abnormální neurologické, imunologické a metabolické nálezy.
U mnoha, ale ne u všech pacientů, kteří splňují kritéria pro CFS, onemocnění začíná akutním, infekčním syndromem. Případy CFS mohou sledovat dobře zdokumentované infekce několika infekčními agens.[66] Studie 259 pacientů s „onemocněním podobným CFS“ publikovaná krátce po objevení HHV-6 použila primární kultury lymfocytů k identifikaci osob s aktivní replikací HHV-6. Taková aktivní replikace byla nalezena u 70% pacientů oproti 20% kontrolních subjektů ().[67] Otázkou, kterou tato studie vznesla, ale na kterou nebyla zodpovězena, bylo, zda nemoc způsobovala jemnou imunitní nedostatečnost, která vedla k reaktivaci HHV-6, nebo zda reaktivace HHV-6 vedla k příznakům nemoci.

Následné studie využívající pouze sérologické techniky, které nerozlišují aktivní a latentní infekci, přinesly smíšené výsledky: většina, ale ne všechny, našla souvislost mezi CFS a HHV-6 infekcí.[66][68][69]
V dalších studiích byly použity testy, které dokážou detekovat aktivní infekci: primární buněčná kultura, PCR séra nebo plazmy nebo testy IgM časných antigenů. Většina těchto studií prokázala souvislost mezi CFS a aktivní infekcí HHV-6,[68][70][71][72][73][74] ačkoli několik ne.[69][75]
Stručně řečeno, aktivní infekce HHV-6 je přítomna u podstatné části pacientů s CFS. Navíc je známo, že HHV-6 infikuje buňky nervového systému a imunitního systému, orgánové systémy s prokazatelnými abnormalitami v CFS. Despite this association, it remains unproven that reactivated HHV-6 infection is a cause of CFS.
Hashimotova tyroiditida
Hashimotova tyroiditida je nejčastější Štítná žláza disease and is characterized by abundant lymfocyt infiltrate and thyroid impairment. Recent research suggests a potential role for HHV-6 (possibly variant A) in the development or triggering of Hashimoto's thyroiditis.[76]
Těhotenství
The role of HHV-6 during pregnancy leading to inflammation in the plodová dutina byl studován.[77]
Neplodnost
HHV-6A DNA was found in the endometrium of almost half of a group of infertile women, but in none of the fertile control group. Přirozené zabijácké buňky specific for HHV-6A, and high uterine levels of certain cytokiny, were also found in the endometrium of the infertile women positive for HHV-6A. The authors suggest that HHV-6A may prove to be an important factor in ženská neplodnost.[78]
Rakovina
Many human oncogenic viruses have been identified. Například, HHV-8 je propojen s Kaposiho sarkom,[79] the Virus Epstein – Barr na Burkittův lymfom, a HPV na rakovina děložního hrdla. Ve skutečnosti Světová zdravotnická organizace estimated (2002) that 17.8% of human cancers were caused by infection.[80] The typical methods whereby viruses initiate oncogenesis involve suppressing the host's immune system, causing inflammation, or altering genes.
HHV-6 has been detected in lymfomy, leukémie, cervical cancers, a mozkové nádory.[14] Rozličný meduloblastom cell lines as well as the cells of other brain tumors have been demonstrated to express the CD46 receptor. Viral DNA has also been identified in many other non-pathological brain tissues, but the levels are lower.[14]
Člověk P53 protein functions as a tumor suppressor. Individuals who do not properly produce this protein experience a higher incidence of cancer, a phenomenon known as Li-Fraumeniho syndrom. One of HHV-6's gene products, the U14 protein, binds P53 and incorporates it into viriony. Another gene product, the ORF-1 protein, can also bind and inactivate P53. Cells expressing the ORF-1 gene have even been shown to produce fibrosarcomas when injected into mice.[14]
Another product of HHV-6, the immediate early protein U95, has been shown to bind nukleární faktor-kappa B. Deregulation of this factor is associated with cancer.[14]
Optická neuritida
HHV-6 induced ocular inflammation has been reported three times. All three were reported in elderly individuals, two during 2007 and one during 2011. The first two were reported in Japan and France, the most recent one in Japan.[81][82][83]
These were believed to have occurred as a result of a reactivation, as anti-HHV-6 IgM antibody levels were low.[83]
Epilepsie spánkového laloku
Epilepsy of the mesial temporal lobe is associated with HHV-6 infection. Within this region of the brain exists three structures: the amygdala, hipokampus, a parahippocampální gyrus. Mesial temporal lobe epilepsy (MTLE) is the most common form of chronic epilepsy and its underlying mechanism is not fully understood.[84]
Researchers consistently report having found HHV-6 DNA in tissues that were removed from patients with MTLE. Studies have demonstrated a tendency for HHV-6 to aggregate in the temporal lobe,[85] with the highest concentrations in astrocytes of the hippocampus.[84]
However, one group of researchers ultimately concluded that HHV-6 may not be involved in MTLE related to Mesial Temporal Sclerosis.[86]
Selhání jater
The virus is a common cause of liver dysfunction and acute liver failure, and has recently been linked to periportal confluent necrosis. Furthermore, HHV-6 DNA is often detectable only in the biopsy tissues as DNA levels fall below the level of detection in blood in persistent cases.[87]
Léčba
There are no pharmaceuticals approved specifically for treating HHV-6 infection, although the usage of Cytomegalovirus treatments (valganciklovir, ganciklovir,[88] cidofovir, a foscarnet ) have shown some success.[7] These drugs are given with the intent of inhibiting proper DNA polymerization by competing with deoxy triphosphate nucleotides[88] or specifically inactivating viral DNA polymerases.[2]
Finding a treatment can be difficult when HHV-6 reactivation occurs following transplant surgery because transplant medications include immunosuppressants.[89]
Reference
- ^ A b C d Adams, M. J.; Carstens, E. B. (2012). "Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2012)". Archivy virologie. 157 (7): 1411–1422. doi:10.1007/s00705-012-1299-6. PMID 22481600.
- ^ A b Jaworska, J.; Gravel, A.; Flamand, L. (2010). "Divergent susceptibilities of human herpesvirus 6 variants to type I interferons". Sborník Národní akademie věd. 107 (18): 8369–74. Bibcode:2010PNAS..107.8369J. doi:10.1073/pnas.0909951107. PMC 2889514. PMID 20404187.
- ^ De Bolle, L.; Van Loon, J.; De Clercq, E.; Naesens, L. (2005). "Quantitative analysis of human herpesvirus 6 cell tropism". Journal of Medical Virology. 75 (1): 76–85. doi:10.1002/jmv.20240. PMID 15543581.
- ^ A b Álvarez-Lafuente, Roberto; García-Montojo, Marta; De Las Heras, Virginia; Bartolomé, Manuel; Arroyo, Rafael (2006). "Clinical parameters and HHV-6 active replication in relapsing—remitting multiple sclerosis patients". Journal of Clinical Virology. 37: S24–6. doi:10.1016/S1386-6532(06)70007-5. PMID 17276363.
- ^ Readhead, Ben; Haure-Mirande, Jean-Vianney; Funk, Cory C.; Richards, Matthew A.; Shannon, Paul; Haroutunian, Vahram; Sano, Mary; Liang, Winnie S.; Beckmann, Noam D.; Price, Nathan D .; Reiman, Eric M .; Schadt, Eric E .; Ehrlich, Michelle E.; Gandy, Sam; Dudley, Joel T. (June 2018). "Multiscale Analysis of Independent Alzheimer's Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks by Human Herpesvirus". Neuron. 99 (1): 64–82.e7. doi:10.1016/j.neuron.2018.05.023. PMC 6551233. PMID 29937276.
- ^ Yoshikawa, Tetsushi (2004). "Human herpesvirus 6 infection in hematopoietic stem cell transplant patients". British Journal of Hematology. 124 (4): 421–32. doi:10.1046/j.1365-2141.2003.04788.x. PMID 14984492.
- ^ A b C d E F G Flamand, Louis; Komaroff, Anthony L.; Arbuckle, Jesse H.; Medveczky, Peter G.; Ablashi, Dharam V. (2010). "Review, part 1: Human herpesvirus-6-basic biology, diagnostic testing, and antiviral efficacy". Journal of Medical Virology. 82 (9): 1560–8. doi:10.1002/jmv.21839. PMID 20648610.
- ^ Anderson, L. (1988). "Human Retrovirus Family: Cancer, Central Nervous System Disease, and AIDS". JNCI Journal of the National Cancer Institute. 80 (13): 987–9. doi:10.1093/jnci/80.13.987. PMID 2842514.
- ^ A b Salahuddin, S.; Ablashi, D.; Markham, P.; Josephs, S.; Sturzenegger, S; Kaplan, M; Halligan, G; Biberfeld, P; et al. (1986). "Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders". Věda. 234 (4776): 596–601. Bibcode:1986Sci...234..596Z. doi:10.1126/science.2876520. PMID 2876520.
- ^ Ablashi, DV; Salahuddin, SZ; Josephs, SF; Imam, F; Lusso, P; Gallo, RC; Hung, C; Lemp, J; Markham, PD (1987). "HBLV (or HHV-6) in human cell lines". Příroda. 329 (6136): 207. Bibcode:1987Natur.329..207A. doi:10.1038/329207a0. PMID 3627265.
- ^ Ablashi, Dharam; Krueger, Gerhard (2006). Human Herpesvirus-6 General Virology, Epidemiology and Clinical Pathology (2. vyd.). Burlington: Elsevier. p.7. ISBN 9780080461281.
- ^ A b Kawabata, A.; Oyaizu, H.; Maeki, T.; Tang, H .; Yamanishi, K.; Mori, Y. (2011). "Analysis of a Neutralizing Antibody for Human Herpesvirus 6B Reveals a Role for Glycoprotein Q1 in Viral Entry". Journal of Virology. 85 (24): 12962–71. doi:10.1128/JVI.05622-11. PMC 3233151. PMID 21957287.
- ^ A b C d E F G h i j Braun, DK; Dominguez, G; Pellett, PE (1997). "Human herpesvirus 6". Recenze klinické mikrobiologie. 10 (3): 521–67. doi:10.1128/CMR.10.3.521. PMC 172933. PMID 9227865.
- ^ A b C d E F Kofman, Alexander; Marcinkiewicz, Lucasz; Dupart, Evan; Lyshchev, Anton; Martynov, Boris; Ryndin, Anatolii; Kotelevskaya, Elena; Brown, Jay; et al. (2011). "The roles of viruses in brain tumor initiation and oncomodulation". Journal of Neuro-Oncology. 105 (3): 451–66. doi:10.1007/s11060-011-0658-6. PMC 3278219. PMID 21720806.
- ^ A b C Arbuckle, J. H.; Medveczky, M. M.; Luka, J.; Hadley, S. H.; Luegmayr, A.; Ablashi, D.; Lund, T. C.; Tolar, J.; et al. (2010). "The latent human herpesvirus-6A genome specifically integrates in telomeres of human chromosomes in vivo and in vitro". Sborník Národní akademie věd. 107 (12): 5563–5568. Bibcode:2010PNAS..107.5563A. doi:10.1073/pnas.0913586107. PMC 2851814. PMID 20212114.
- ^ A b C d E Harberts, E.; Yao, K.; Wohler, J. E.; Maric, D.; Ohayon, J.; Henkin, R.; Jacobson, S. (2011). "Human herpesvirus-6 entry into the central nervous system through the olfactory pathway". Sborník Národní akademie věd. 108 (33): 13734–9. Bibcode:2011PNAS..10813734H. doi:10.1073/pnas.1105143108. PMC 3158203. PMID 21825120.
- ^ Fenner, F. (1971). "The nomenclature and classification of viruses the International Committee on Nomenclature of Viruses". Virologie. 46 (3): 979–980. doi:10.1016/0042-6822(71)90102-4. PMID 18619371.
- ^ Fenner, F. (1976). "Classification and nomenclature of viruses. Second report of the International Committee on Taxonomy of Viruses". Intervirologie. 7 (1–2): 1–115. doi:10.1159/000149938. PMID 826499.
- ^ Davison AJ (2010) Herpesvirus systematics. Veterinář Microbiol. 143(1–2): 52–69
- ^ A b Kawabata, Akiko; Tang, Huamin; Huang, Honglan; Yamanishi, Koichi; Mori, Yasuko (2009). "Y Human herpesvirus 6 envelope components enriched in lipid rafts: Evidence for virion-associated lipid rafts". Virology Journal. 6: 127. doi:10.1186/1743-422X-6-127. PMC 2743664. PMID 19689819.
- ^ A b C d E Dominguez, G.; Dambaugh, T. R.; Stamey, F. R.; Dewhurst, S.; Inoue, N.; Pellett, P. E. (1999). "Human herpesvirus 6B genome sequence: Coding content and comparison with human herpesvirus 6A". Journal of Virology. 73 (10): 8040–8052. doi:10.1128/JVI.73.10.8040-8052.1999. PMC 112820. PMID 10482553.
- ^ A b Tang, Huamin; Kawabata, Akiko; Yoshida, Mayumi; Oyaizu, Hiroko; Maeki, Takahiro; Yamanishi, Koichi; Mori, Yasuko (2010). "Human herpesvirus 6 encoded glycoprotein Q1 gene is essential for virus growth". Virologie. 407 (2): 360–7. doi:10.1016/j.virol.2010.08.018. PMID 20863544.
- ^ A b C d E F G h i Arbuckle, Jesse H.; Medveczky, Peter G. (2011). "Molekulární biologie latence lidského herpesviru-6 a integrace telomer". Mikroby a infekce. 13 (8–9): 731–41. doi:10.1016 / j.micinf.2011.03.006. PMC 3130849. PMID 21458587.
- ^ A b C Borenstein, R.; Frenkel, N. (2009). „Klonování genomu lidského herpes viru 6A do bakteriálních umělých chromozomů a studium meziproduktů replikace DNA“. Sborník Národní akademie věd. 106 (45): 19138–19143. Bibcode:2009PNAS..10619138B. doi:10.1073 / pnas.0908504106. PMC 2767366. PMID 19858479.
- ^ A b C d E Yoshikawa, Tetsushi; Asano, Yoshizo; Akimoto, Shiho; Ozaki, Takao; Iwasaki, Takuya; Kurata, Takeshi; Goshima, Fumi; Nishiyama, Yukihiro (2002). "Latent infection of human herpesvirus 6 in astrocytoma cell line and alteration of cytokine synthesis". Journal of Medical Virology. 66 (4): 497–505. doi:10.1002/jmv.2172. PMID 11857528.
- ^ A b Tuddenham, L.; Jung, J. S.; Chane-Woon-Ming, B.; Dolken, L.; Pfeffer, S. (2011). "Small RNA Deep Sequencing Identifies MicroRNAs and Other Small Noncoding RNAs from Human Herpesvirus 6B". Journal of Virology. 86 (3): 1638–49. doi:10.1128/JVI.05911-11. PMC 3264354. PMID 22114334.
- ^ Tang, H .; Hayashi, M.; Maeki, T.; Yamanishi, K.; Mori, Y. (2011). "Human Herpesvirus 6 Glycoprotein Complex Formation is Required for Folding and Trafficking of the gH/gL/gQ1/gQ2 Complex and Its Cellular Receptor Binding". Journal of Virology. 85 (21): 11121–30. doi:10.1128/JVI.05251-11. PMC 3194968. PMID 21849437.
- ^ Greenstone, H. L.; Santoro, F; Lusso, P; Berger, EA (2002). "Human Herpesvirus 6 and Measles Virus Employ Distinct CD46 Domains for Receptor Function". Journal of Biological Chemistry. 277 (42): 39112–8. doi:10.1074/jbc.M206488200. PMID 12171934.
- ^ A b Mori, Yasuko (2009). "Recent topics related to human herpesvirus 6 cell tropism". Buněčná mikrobiologie. 11 (7): 1001–6. doi:10.1111/j.1462-5822.2009.01312.x. PMID 19290911.
- ^ J Exp Med. 1995 Apr 1;181(4):1303–10. Infection of gamma/delta T lymphocytes by human herpesvirus 6: transcriptional induction of CD4 and susceptibility to HIV infection. Lusso P, Garzino-Demo A, Crowley RW, Malnati MS.
- ^ A b C Liedtke, W.; Opalka, B.; Zimmermann, C.W.; Lignitz, E. (1993). "Age distribution of latent herpes simplex virus 1 and varicella-zoster virus genome in human nervous tissue". Časopis neurologických věd. 116 (1): 6–11. doi:10.1016/0022-510X(93)90082-A. PMID 8389816.
- ^ A b Cassiani-Ingoni, Riccardo; Greenstone, Heather L.; Donati, Donatella; Fogdell-Hahn, Anna; Martinelli, Elena; Refai, Daniel; Martin, Roland; Berger, Edward A.; Jacobson, Steven (2005). "CD46 on glial cells can function as a receptor for viral glycoprotein-mediated cell-cell fusion". Glia. 52 (3): 252–8. doi:10.1002/glia.20219. PMID 15920733.
- ^ A b C Morissette, G.; Flamand, L. (2010). "Herpesviruses and Chromosomal Integration". Journal of Virology. 84 (23): 12100–9. doi:10.1128/JVI.01169-10. PMC 2976420. PMID 20844040.
- ^ Potenza, Leonardo; Barozzi, Patrizia; Torelli, Giuseppe; Luppi, Mario (2010). "Translational challenges of human herpesvirus 6 chromosomal integration". Budoucí mikrobiologie. 5 (7): 993–5. doi:10.2217/fmb.10.74. hdl:11380/704738. PMID 20632798.
- ^ A b C Kaufer, B. B.; Jarosinski, K. W.; Osterrieder, N. (2011). "Herpesvirus telomeric repeats facilitate genomic integration into host telomeres and mobilization of viral DNA during reactivation". Journal of Experimental Medicine. 208 (3): 605–15. doi:10.1084/jem.20101402. PMC 3058580. PMID 21383055.
- ^ Isegawa, Yuji; Matsumoto, Chisa; Nishinaka, Kazuko; Nakano, Kazushi; Tanaka, Tatsuya; Sugimoto, Nakaba; Ohshima, Atsushi (2010). "PCR with quenching probes enables the rapid detection and identification of ganciclovir-resistance-causing U69 gene mutations in human herpesvirus 6". Molekulární a buněčné sondy. 24 (4): 167–77. doi:10.1016/j.mcp.2010.01.002. PMID 20083192.
- ^ Lusso, Paolo; De Maria, Andrea; Malnati, Mauro; Lori, Franco; Derocco, Susan E.; Baseler, Michael; Gallo, Robert C. (1991). "Induction of CD4 and susceptibility to HIV-1 infection in human CD8+ T lymphocytes by human herpesvirus 6". Příroda. 349 (6309): 533–5. Bibcode:1991Natur.349..533L. doi:10.1038/349533a0. PMID 1846951.
- ^ Arena, A; Liberto, MC; Capozza, AB; Focà, A (1997). "Productive HHV-6 infection in differentiated U937 cells: Role of TNF alpha in regulation of HHV-6". Nová mikrobiologie. 20 (1): 13–20. PMID 9037664.
- ^ Inagi, Reiko; Guntapong, Ratigorn; Nakao, Masayuki; Ishino, Yoshizumi; Kawanishi, Kazunobu; Isegawa, Yuji; Yamanishi, Koichi (1996). "Human herpesvirus 6 induces IL-8 gene expression in human hepatoma cell line, Hep G2". Journal of Medical Virology. 49 (1): 34–40. doi:10.1002/(SICI)1096-9071(199605)49:1<34::AID-JMV6>3.0.CO;2-L. PMID 8732869.
- ^ Lusso, P.; Crowley, R. W.; Malnati, M. S.; Di Serio, C.; Ponzoni, M.; Biancotto, A.; Markham, P. D.; Gallo, R. C. (2007). "Human herpesvirus 6A accelerates AIDS progression in macaques". Sborník Národní akademie věd. 104 (12): 5067–72. Bibcode:2007PNAS..104.5067L. doi:10.1073/pnas.0700929104. JSTOR 25427145. PMC 1829265. PMID 17360322.
- ^ Hall, Caroline Breese; Long, Christine E.; Schnabel, Kenneth C.; Caserta, Mary T.; McIntyre, Kim M.; Costanzo, Maria A.; Knott, Anne; Dewhurst, Stephen; et al. (1994). "Human Herpesvirus-6 Infection in Children -- A Prospective Study of Complications and Reactivation". New England Journal of Medicine. 331 (7): 432–8. doi:10.1056/NEJM199408183310703. PMID 8035839.
- ^ Newly Found Herpes Virus Is Called Major Cause of Illness in Young, New York Times
- ^ Okuno, T; Takahashi, K; Balachandra, K; Shiraki, K; Yamanishi, K; Takahashi, M; Baba, K (1989). "Seroepidemiology of human herpesvirus 6 infection in normal children and adults". Journal of Clinical Microbiology. 27 (4): 651–3. doi:10.1128/JCM.27.4.651-653.1989. PMC 267390. PMID 2542358.
- ^ Araujo, A.; Pagnier, A.; Frange, P.; Wroblewski, I.; Stasia, M.-J.; Morand, P.; Plantaz, D. (2011). "Syndrome d'activation lymphohistiocytaire associé à une infection à Burkholderia cepacia complex chez un nourrisson révélant une granulomatose septique et une intégration génomique du virus HHV-6" [Lymphohistiocytic activation syndrome and Burkholderia cepacia complex infection in a child revealing chronic granulomatous disease and chromosomal integration of the HHV-6 genome]. Archives de Pédiatrie (francouzsky). 18 (4): 416–9. doi:10.1016/j.arcped.2011.01.006. PMID 21397473.
- ^ Agut Henri, Bonnafous Pascale, Gautheret-Dejean Agnès (2015). "Laboratory and Clinical Aspects of Human Herpesvirus 6 Infections". Recenze klinické mikrobiologie. 28: 313–335. doi:10.1128/CMR.00122-14.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Alvarez-Lafuente, R.; Martin-Estefania, C.; De Las Heras, V.; Castrillo, C.; Cour, I.; Picazo, J.J.; Varela De Seijas, E.; Arroyo, R. (2002). "Prevalence of herpesvirus DNA in MS patients and healthy blood donors". Acta Neurologica Scandinavica. 105 (2): 95–9. doi:10.1034/j.1600-0404.2002.1o050.x. PMID 11903118.
- ^ Komaroff, Anthony L. (2006). "Is human herpesvirus-6 a trigger for chronic fatigue syndrome?". Journal of Clinical Virology. 37: S39–46. doi:10.1016/S1386-6532(06)70010-5. PMID 17276367.
- ^ HHV-6 and AIDS Archivováno 8 února 2007 na Wayback Machine, Wisconsin Viral Research Group
- ^ Fotheringham, Julie; Donati, Donatella; Akhyani, Nahid; Fogdell-Hahn, Anna; Vortmeyer, Alexander; Heiss, John D.; Williams, Elizabeth; Weinstein, Steven; et al. (2007). "Association of Human Herpesvirus-6B with Mesial Temporal Lobe Epilepsy". PLOS Medicine. 4 (5): e180. doi:10.1371/journal.pmed.0040180. PMC 1880851. PMID 17535102.[nespolehlivý lékařský zdroj? ]
- ^ Challoner, Peter B.; Smith, Kirsten T.; Parker, Jay D.; MacLeod, David L.; Coulter, Silvija N.; Rose, Timothy M.; Schultz, Emily R.; Lindsley Bennett, J.; et al. (1995). "Plaque-Associated Expression of Human Herpesvirus 6 in Multiple Sclerosis". Sborník Národní akademie věd Spojených států amerických. 92 (16): 7440–4. Bibcode:1995PNAS...92.7440C. doi:10.1073/pnas.92.16.7440. PMC 41355. PMID 7638210.
- ^ Compston, Alastair; Coles, Alasdair (2008). "Multiple sclerosis". Lancet. 372 (9648): 1502–17. doi:10.1016/S0140-6736(08)61620-7. PMID 18970977.
- ^ Compston, Alastair; Coles, Alasdair (2002). "Multiple sclerosis". Lancet. 359 (9313): 1221–31. doi:10.1016/S0140-6736(02)08220-X. PMID 11955556.
- ^ A b Hernán, Miguel A.; Zhang, Shumin M.; Lipworth, Loren; Olek, Michael J.; Ascherio, Alberto (2001). "Multiple Sclerosis and Age at Infection with Common Viruses". Epidemiologie. 12 (3): 301–6. doi:10.1097/00001648-200105000-00009. PMID 11337603.
- ^ Delbue, Serena; Carluccio, Silvia; Ferrante, Pasquale (2012). "The long and evolving relationship between viruses and multiple sclerosis". Budoucí virologie. 7 (9): 871–83. doi:10.2217/fvl.12.78.
- ^ Rotola, A; Merlotti, I; Caniatti, L; Caselli, E; Granieri, E; Tola, MR; Di Luca, D; Cassai, E (2004). "Human herpesvirus 6 infects the central nervous system of multiple sclerosis patients in the early stages of the disease". Roztroušená skleróza. 10 (4): 348–54. doi:10.1191/1352458504ms1045oa. PMID 15327028.
- ^ Ortega-Madueno, I.; Garcia-Montojo, M.; Dominguez-Mozo, M.; Arias-Leal, A.; Casanova, I. (2014). "Anti-human herpesvirus 6A/B IgG correlates with relapses and progression in multiple sclerosis". PLOS One. 9 (8): e104836. Bibcode:2014PLoSO...9j4836O. doi:10.1371/journal.pone.0104836. PMC 4128748. PMID 25110949.
- ^ A b Kamei, A.; Ichinohe, S.; Onuma, R.; Hiraga, S.; Fujiwara, T. (1997). "Acute disseminated demyelination due to primary human herpesvirus-6 infection". European Journal of Pediatrics. 156 (9): 709–12. doi:10.1007/s004310050695. PMID 9296535.
- ^ A b Cirone, Mara; Cuomo, Laura; Zompetta, Claudia; Ruggieri, Stefano; Frati, Luigi; Faggioni, Alberto; Ragona, Giuseppe (2002). "Human herpesvirus 6 and multiple sclerosis: A study of t cell cross-reactivity to viral and myelin basic protein antigens". Journal of Medical Virology. 68 (2): 268–72. doi:10.1002/jmv.10190. PMC 7167112. PMID 12210418.
- ^ A b Tejada-Simon, Maria V.; Zang, Ying C. Q.; Hong, Jian; Rivera, Victor M.; Zhang, Jingwu Z. (2003). "Cross-reactivity with myelin basic protein and human herpesvirus-6 in multiple sclerosis". Annals of Neurology. 53 (2): 189–97. doi:10.1002/ana.10425. PMID 12557285.
- ^ A b C Tait, A; Straus, S (2008). "Phosphorylation of U24 from Human Herpes Virus type 6 (HHV-6) and its potential role in mimicking myelin basic protein (MBP) in multiple sclerosis". FEBS Dopisy. 582 (18): 2685–8. doi:10.1016/j.febslet.2008.06.050. PMID 18616943.
- ^ Knox, Konstance K.; Brewer, Joseph H.; Henry, James M.; Harrington, Daniel J.; Carrigan, Donald R. (2000). "Human Herpesvirus 6 and Multiple Sclerosis: Systemic Active Infections in Patients with Early Disease". Klinické infekční nemoci. 31 (4): 894–903. doi:10.1086/318141. PMID 11049767.
- ^ Pietilainen-Nicklen, J.; Virtanen, J (2014). "HHV-6-positivity in diseases with demyelination". Journal of Clinical Virology. 61 (2): 216–9. doi:10.1016/j.jcv.2014.07.006. PMID 25088617.
- ^ Kawamura, Yoshiki; Sugata, Ken; Ihira, Masaru; Mihara, Takateru; Mutoh, Tatsuro; Asano, Yoshizo; Yoshikawa, Tetsushi (2011). "Different characteristics of human herpesvirus 6 encephalitis between primary infection and viral reactivation". Journal of Clinical Virology. 51 (1): 12–9. doi:10.1016/j.jcv.2011.02.002. PMID 21376661.
- ^ Nora-Krukle, Z; Chapenko, S; Logina, I; Millers, A; Platkajis, A; Murovska, M (2011). "Human herpesvirus 6 and 7 reactivation and disease activity in multiple sclerosis". Medicina. 47 (10): 527–31. doi:10.3390/medicina47100075. PMID 22186115.
- ^ Fukuda, K; Straus, SE; Hickie, I; Sharpe, MC; Dobbins, JG; Komaroff, A (1994). "The chronic fatigue syndrome: A comprehensive approach to its definition and study. International Chronic Fatigue Syndrome Study Group". Annals of Internal Medicine. 121 (12): 953–9. doi:10.7326/0003-4819-121-12-199412150-00009. PMID 7978722.
- ^ A b Hickie, I .; Davenport, T; Wakefield, D; Vollmer-Conna, U; Cameron, B; Vernon, SD; Reeves, WC; Lloyd, A; Dubbo Infection Outcomes Study Group (2006). "Post-infective and chronic fatigue syndromes precipitated by viral and non-viral pathogens: Prospective cohort study". BMJ. 333 (7568): 575. doi:10.1136/bmj.38933.585764.AE. PMC 1569956. PMID 16950834.
- ^ Buchwald, D; Cheney, PR; Peterson, DL; Henry, B; Wormsley, SB; Geiger, A; Ablashi, DV; Salahuddin, SZ; et al. (1992). "A chronic illness characterized by fatigue, neurologic and immunologic disorders, and active human herpesvirus type 6 infection". Annals of Internal Medicine. 116 (2): 103–13. doi:10.7326/0003-4819-116-2-103. PMID 1309285.
- ^ A b Ablashi, D.V; Eastman, H.B; Owen, C.B; Roman, M.M; Friedman, J; Zabriskie, J.B; Peterson, D.L; Pearson, G.R; Whitman, J.E (2000). "Frequent HHV-6 reactivation in multiple sclerosis (MS) and chronic fatigue syndrome (CFS) patients". Journal of Clinical Virology. 16 (3): 179–91. doi:10.1016/S1386-6532(99)00079-7. PMID 10738137.
- ^ A b Reeves, W. C.; Stamey, F. R.; Black, J. B.; Mawle, A. C.; Stewart, J. A.; Pellett, P. E. (2000). "Human Herpesviruses 6 and 7 in Chronic Fatigue Syndrome: A Case-Control Study". Klinické infekční nemoci. 31 (1): 48–52. doi:10.1086/313908. PMID 10913395.
- ^ Nicolson, G. L.; Gan, R.; Haier, J. (2003). "Multiple co-infections (Mycoplasma, Chlamydia, human herpes virus-6) in blood of chronic fatigue syndrome patients: Association with signs and symptoms". APMIS. 111 (5): 557–66. doi:10.1034/j.1600-0463.2003.1110504.x. PMID 12887507.
- ^ Patnaik, M.; Komaroff, A. L.; Conley, E.; Ojo-Amaize, E. A.; Peter, J. B. (1995). "Prevalence of IgM Antibodies to Human Herpesvirus 6 Early Antigen (p41/38) in Patients with Chronic Fatigue Syndrome". Journal of Infection Diseases. 172 (5): 1364–7. doi:10.1093/infdis/172.5.1364. PMID 7594679.
- ^ Secchiero, P.; Carrigan, D. R.; Asano, Y.; Benedetti, L.; Crowley, R. W.; Komaroff, A. L.; Gallo, R. C.; Lusso, P. (1995). "Detection of Human Herpesvirus 6 in Plasma of Children with Primary Infection and Immunosuppressed Patients by Polymerase Chain Reaction". Journal of Infection Diseases. 171 (2): 273–80. doi:10.1093/infdis/171.2.273. PMID 7844362.
- ^ Wagner, Mathias; Krueger, Gerhard; Ablashi, Dharam; Whitman, James (1996). "Chronic Fatigue Syndrome (CFS): A Critical Evaluation of Testing for Active Human Herpesvirus-6 (HHV-6) Infection". Journal of Chronic Fatigue Syndrome. 2 (4): 3–16. doi:10.1300/J092v02n04_02.
- ^ Zorzenon, Marcella; Rukh, Gull; Botta, Giuseppe; Colle, Roberto; Barsanti, Laura; Ceccherini-Nelli, Luca (1996). "Active HHV-6 Infection in Chronic Fatigue Syndrome Patients from Italy". Journal of Chronic Fatigue Syndrome. 2: 3–12. doi:10.1300/J092v02n01_02.
- ^ Koelle, David M .; Barcy, Serge; Huang, Meei‐Li; Ashley, Rhoda L.; Corey, Lawrence; Zeh, Judy; Ashton, Suzanne; Buchwald, Dedra (2002). "Markers of Viral Infection in Monozygotic Twins Discordant for Chronic Fatigue Syndrome". Klinické infekční nemoci. 35 (5): 518–25. doi:10.1086/341774. PMID 12173124.
- ^ Caselli, Elisabetta; Zatelli, Maria Chiara; Rizzo, Roberta; Benedetti, Sabrina; Martorelli, Debora; Trasforini, Giorgio; Cassai, Enzo; Degli Uberti, Ettore C.; et al. (Říjen 2012). Moore, Patrick S (ed.). "Virologic and immunologic evidence supporting an association between HHV-6 and Hashimoto's thyroiditis". PLOS patogeny. 8 (10): e1002951. doi:10.1371/journal.ppat.1002951. PMC 3464215. PMID 23055929.
- ^ Silasi, Michelle (March 2013). "Viral invasion of the Amniotic Cavity (VIAC) in the Midtrimester of Pregnancy". American Journal of Reproductive Immunology. 69 (3): 195–196. doi:10.1111/aji.12073. PMID 23384234.
- ^ Marci Roberto, Gentili Valentina, Bortolotti Daria, Lo Monte Giuseppe, Caselli Elisabetta, Bolzani Silvia, Rotola Antonella, Di Luca Dario, Rizzo Roberta (2016). "Presence of HHV-6A in Endometrial Epithelial Cells from Women with Primary Unexplained Infertility". PLOS One. 11 (7): e0158304. Bibcode:2016PLoSO..1158304M. doi:10.1371/journal.pone.0158304. PMC 4930213. PMID 27367597.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Tamburro, Kristen; Yang, Dongmei; Poisson, Jessica; Fedoriw, Yuri; Roy, Debasmita; Lucas, Amy; Sin, Sang-Hoon; Malouf, Nadia; et al. (10 November 2012). "Vironome of Kaposi sarcoma associated herpesvirus-inflammatory cytokine syndrome in an AIDS patient reveals co-infection of human herpesvirus 8 and human herpesvirus 6A". Virologie. 433 (1): 220–225. doi:10.1016/j.virol.2012.08.014. PMC 3505605. PMID 22925337.
- ^ Parkin, Donald Maxwell (2006). „Globální zdravotní zátěž způsobená rakovinou související s infekcí v roce 2002“. International Journal of Cancer. 118 (12): 3030–44. doi:10.1002 / ijc.21731. PMID 16404738.
- ^ Oberacher-Velten, Isabel M.; Jonas, Jost B.; Jünemann, Anselm; Schmidt, Barbara (2004). "Bilateral optic neuropathy and unilateral tonic pupil associated with acute human herpesvirus 6 infection: A case report". Graefe's Archive for Clinical and Experimental Ophthalmology. 243 (2): 175–7. doi:10.1007/s00417-004-0986-8. PMID 15742213.
- ^ Maslin, Jérôme; Bigaillon, Christine; Froussard, Françoise; Dost, Vincent; Nicand, Elisabeth (2007). "Acute bilateral uveitis associated with an active human herpesvirus-6 infection". Journal of Infection. 54 (4): e237–40. doi:10.1016/j.jinf.2006.12.012. PMID 17303245.
- ^ A b Ogata, Nahoko; Koike, Naoko; Yoshikawa, Tadanobu; Takahashi, Kanji (2011). "Human herpesvirus 6-associated uveitis with optic neuritis diagnosed by multiplex PCR". Japanese Journal of Ophthalmology. 55 (5): 502–5. doi:10.1007/s10384-011-0069-4. PMID 21814813.
- ^ A b Li, Jin-Mei; Lei, Ding; Peng, Fang; Zeng, Yi-Jun; Li, Liang; Xia, Zeng-Liang; Xia, Xiao-Qiang; Zhou, Dong (2011). "Detection of human herpes virus 6B in patients with mesial temporal lobe epilepsy in West China and the possible association with elevated NF-κB expression". Výzkum epilepsie. 94 (1–2): 1–9. doi:10.1016/j.eplepsyres.2010.11.001. PMID 21256714.
- ^ Noguchi, T.; Yoshiura, T.; Hiwatashi, A.; Togao, O.; Yamashita, K.; Nagao, E.; Uchino, A.; Hasuo, K.; et al. (2010). "CT and MRI Findings of Human Herpesvirus 6-Associated Encephalopathy: Comparison with Findings of Herpes Simplex Virus Encephalitis". American Journal of Roentgenology. 194 (3): 754–60. doi:10.2214/AJR.09.2548. PMID 20173155.
- ^ Niehusmann, Pitt; Mittelstaedt, Tobias; Bien, Christian G.; Drexler, Jan F.; Grote, Alexander; Schoch, Susanne; Becker, Albert J. (2010). "Presence of human herpes virus 6 DNA exclusively in temporal lobe epilepsy brain tissue of patients with history of encephalitis". Epilepsie. 51 (12): 2478–83. doi:10.1111/j.1528-1167.2010.02741.x. PMID 21204811.
- ^ Buyse, S.; Roque-Afonso, A.-M.; Vaghefi, P.; Gigou, M.; Dussaix, E.; Duclos-Vallee, J.-C.; Samuel, D.; Guettier, C. (2013). "Acute Hepatitis With Periportal Confluent Necrosis Associated With Human Herpesvirus 6 Infection in Liver Transplant Patients". American Journal of Clinical Pathology. 140 (3): 403–9. doi:10.1309/AJCP0FWI2XAHECBJ. PMID 23955460.
- ^ A b Nakano, Kazushi; Nishinaka, Kazuko; Tanaka, Tatsuya; Ohshima, Atsushi; Sugimoto, Nakaba; Isegawa, Yuji (2009). "Detection and identification of U69 gene mutations encoded by ganciclovir-resistant human herpesvirus 6 using denaturing high-performance liquid chromatography". Journal of Virological Methods. 161 (2): 223–30. doi:10.1016/j.jviromet.2009.06.016. PMID 19559728.
- ^ Shiley, Kevin; Blumberg, Emily (2010). "Herpes Viruses in Transplant Recipients: HSV, VZV, Human Herpes Viruses, and EBV". Kliniky infekčních nemocí v Severní Americe. 24 (2): 373–93. doi:10.1016/j.idc.2010.01.003. PMID 20466275.
externí odkazy
 Média související s Lidský herpesvirus 6 na Wikimedia Commons
Média související s Lidský herpesvirus 6 na Wikimedia Commons Údaje týkající se Lidský herpesvirus 6 na Wikispecies
Údaje týkající se Lidský herpesvirus 6 na Wikispecies