Polyomaviridae - Polyomaviridae
| Polyomaviridae | |
|---|---|
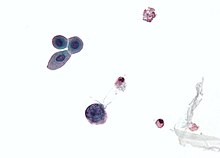 | |
| Mikrograf zobrazeno a polyomavirus infikovaná buňka - velká (modrá) buňka dole uprostřed vlevo. Cytologický vzorek moči. | |
| Klasifikace virů | |
| (bez hodnocení): | Virus |
| Oblast: | Monodnaviria |
| Království: | Shotokuvirae |
| Kmen: | Cossaviricota |
| Třída: | Papovaviricetes |
| Objednat: | Sepolyvirales |
| Rodina: | Polyomaviridae |
| Rody | |
Polyomaviridae je rodina viry jehož přirozené hostitelé jsou to především savci a ptáci.[1][2] Od roku 2019 existují čtyři uznané rody a 102 druhů, z nichž devět není přiřazeno rodu.[3] Je známo, že 14 druhů infikuje člověka, zatímco jiné, jako např Virus opičí 40, byly u lidí identifikovány v menší míře.[4][5] Většina z těchto virů je velmi běžná a typicky asymptomatická u většiny zkoumaných lidských populací.[6][7] Virus BK je spojován s nefropatie v transplantace ledvin a nerenální pacienti po transplantaci pevných orgánů,[8][9] Virus JC s progresivní multifokální leukoencefalopatie,[10] a Virus Merkelových buněk s Rakovina Merkelových buněk.[11]
Struktura a genom

Polyomaviry jsou neobalený dvouvláknová DNA viry s oběžníkem genomy kolem 5000 základní páry. Genom je zabalen v a virová kapsida asi 40-50 nanometry v průměru, což je icosahedral ve tvaru (T = 7 symetrie).[2][12] Kapsida se skládá ze 72 pentamerik kapsomery bílkoviny zvané VP1, který je schopen se shromáždit do uzavřeného dvacetistěnu;[13] každý pentamer VP1 je spojen s jednou molekulou jednoho z dalších dvou kapsidových proteinů, VP2 nebo VP3.[5]

Genom typického polyomaviru kóduje mezi 5 a 9 bílkoviny, rozdělena na dvě části transkripční regiony nazývané časné a pozdní regiony kvůli době během infekce, ve které jsou přepsány. Každá oblast je přepsána hostitelskou buňkou RNA polymeráza II jako singl pre-poselská RNA obsahující více genů. Časná oblast obvykle kóduje dva proteiny, malé a velké nádorové antigeny produkované alternativní sestřih. Pozdní oblast obsahuje tři kapsidové strukturní proteiny VP1, VP2 a VP3, produkované alternativně překladový začít stránky. U některých virů jsou přítomny další geny a další variace na toto téma: například polyomaviry hlodavců mají třetí protein zvaný antigen středního nádoru v rané oblasti, což je při navádění extrémně efektivní buněčná transformace; SV40 má další kapsidový protein VP4; některé příklady obsahují další regulační protein agnoprotein vyjádřeno z pozdní oblasti. Genom také obsahuje a nekódování kontrolní nebo regulační oblast obsahující časnou a pozdní oblast ' promotéři, počáteční stránky transkripce a původ replikace.[2][12][5][15]
| Rod | Struktura | Symetrie | Capsid | Genomické uspořádání | Genomická segmentace |
|---|---|---|---|---|---|
| Polyomavirus | Icosahedral | T = 7 | Neobalené | Oběžník | Monopartitní |
Replikace a životní cyklus
Životní cyklus polyomaviru začíná vstupem do a hostitelská buňka. Buněčné receptory pro polyomaviry jsou kyselina sialová zbytky glykany, běžně gangliosidy. Připojení polyomavirů k hostitelským buňkám je zprostředkováno vazbou VP1 na sialylované glykany na povrchu buněk.[2][12][15][16] U některých konkrétních virů dochází k dalším interakcím na povrchu buněk; například Virus JC se předpokládá, že vyžaduje interakci s 5HT2A receptor a Virus Merkelových buněk s heparan sulfát.[15][17] Interakce virus-buňka jsou však obecně zprostředkovány běžně se vyskytujícími molekulami na buněčném povrchu, a proto pravděpodobně nepůsobí významným způsobem k pozorování buněčného typu jednotlivých virů tropismus.[15] Po navázání na molekuly na povrchu buňky je to virion endocytovaný a vstoupí do endoplazmatické retikulum - chování jedinečné mezi známými neobalenými viry[18] - kde je pravděpodobné, že struktura virové kapsidy bude narušena působením hostitelské buňky disulfid izomeráza enzymy.[2][12][19]
Podrobnosti o přenosu do jádra nejsou jasné a mohou se u jednotlivých polyomavirů lišit. Často se uvádí, že intaktní, i když zkreslená virionová částice se uvolňuje z endoplazmatického retikula do buněčné cytoplazmy, kde se genom uvolňuje z kapsidy, pravděpodobně kvůli nízkému vápník koncentrace v cytoplazmě.[18] Exprese virových genů i replikace virového genomu se vyskytují v jádro pomocí strojního vybavení hostitelské buňky. Rané geny - zahrnující minimálně malý nádorový antigen (Vydržet velký nádorový antigen (LT) - jsou vyjádřeny jako první, z jediného alternativně sestříhané messenger RNA pramen. Tyto proteiny slouží k manipulaci s hostitelem buněčný cyklus - dysregulovat přechod z Fáze G1 na S fáze, když se replikuje genom hostitelské buňky - protože pro replikaci virového genomu je zapotřebí replikační mechanismus DNA hostitelské buňky.[2][12][15] Přesný mechanismus této dysregulace závisí na viru; například, SV40 LT může přímo vázat hostitelskou buňku p53, ale myší polyomavirus LT ne.[20] LT indukuje replikaci DNA z nekódující kontrolní oblasti virového genomu (NCCR), po které je snížena exprese časné mRNA a začíná exprese pozdní mRNA, která kóduje proteiny virového kapsidu.[19] Jak tyto interakce začínají, LT patřící k několika polyomavirům, včetně Polyomavirus z Merkelových buněk, představují onkogenní potenciál.[21]Bylo popsáno několik mechanismů pro regulaci přechodu z časné na pozdní genovou expresi, včetně zapojení proteinu LT do potlačování časného promotoru,[19] exprese nedokončených pozdních mRNA s prodloužením komplementárním k časné mRNA,[15] a vyjádření regulace mikroRNA.[15]Exprese pozdních genů vede k akumulaci virových kapsidových proteinů v cytoplazmě hostitelské buňky. Složky kapsidy vstupují do jádra za účelem enkapsidace nové virové genomové DNA. Mohou být smontovány nové virióny virové továrny.[2][12] Mechanismus uvolňování viru z hostitelské buňky se u polyomavirů liší; některé exprimují proteiny, které usnadňují odchod z buňky, například agnoprotein nebo VP4.[19] V některých případech vede vysoká hladina enkapsidovaného viru k buňce lýza, uvolňující viriony.[15]
| Rod | Podrobnosti o hostiteli | Tkáňový tropismus | Vstupní údaje | Podrobnosti o vydání | Replikační web | Místo montáže | Přenos |
|---|---|---|---|---|---|---|---|
| Polyomavirus | Savci; ptactvo | Dýchací systém; ledviny, mozek | Endocytóza buněčných receptorů | Lyza | Jádro | Jádro | Orálně-fekální |
Virové proteiny
Nádorové antigeny
The velký nádorový antigen hraje klíčovou roli v regulaci životního cyklu viru vazbou na virový původ replikace DNA, kde podporuje syntézu DNA. Protože polyomavirus spoléhá na to, že hostitelská buňka replikuje, hostitelská buňka musí být v s-fázi, aby to mohlo začít. Díky tomu velký T-antigen také moduluje buněčné signální dráhy, aby stimuloval progresi buněčného cyklu vazbou na řadu buněčných kontrolních proteinů.[22] Toho je dosaženo útokem dvou hrotů, kterým je inhibice tumor supresních genů p53 a členů retinoblastom (pRB) rodina,[23] a stimulace buněčných růstových drah vazbou buněčné DNA, ATPázy-helikázy, asociace DNA polymerázy a a vazbou transkripčních preinitačních komplexních faktorů.[24] Tato abnormální stimulace buněčného cyklu je silnou silou pro onkogenní transformaci.
The malý nádorový antigen protein je také schopen aktivovat několik buněčných drah, které stimulují buněčnou proliferaci. Polyomavirové malé T antigeny se běžně zaměřují na proteinovou fosfatázu 2A (PP2A ),[25] klíčový regulátor více podjednotek více cest včetně Akt, mitogenem aktivovaná proteinová kináza (MAPK) a stresem aktivovaná proteinová kináza (SAPK).[26][27] Polyomavirus z Merkelových buněk malý T antigen kóduje jedinečnou doménu, nazývanou LT-stabilizační doména (LSD), která se váže na a inhibuje FBXW7 E3 ligáza regulující jak buněčné, tak virové onkoproteiny.[28] Na rozdíl od SV40 MCV malý T antigen přímo transformuje buňky hlodavců in vitro.[29]
The antigen středního nádoru se používá v modelové organismy vyvinuté ke studiu rakoviny, jako je MMTV-PyMT systém, kde je střední T spojen s MMTV promotér. Tam funguje jako onkogen, zatímco tkáň, kde se nádor vyvíjí, je určena promotorem MMTV.
Kapsidové proteiny
Polyomavirová kapsida se skládá z jedné hlavní složky, hlavní kapsidový protein VP1 a jedna nebo dvě vedlejší součásti, minoritní kapsidové proteiny VP2 a VP3. VP1 pentamery tvoří uzavřený icosahedral virová kapsida, a uvnitř kapsidy je každý pentamer spojen s jednou molekulou buď VP2 nebo VP3.[5][30] Některé polyomaviry, jako např Polyomavirus z Merkelových buněk, nekódujte ani nevyjadřujte VP3.[31] Kapsidové proteiny jsou exprimovány z pozdní oblasti genomu.[5]
Agnoprotein
The agnoprotein je malý multifunkční fosfo-protein nacházející se v pozdní kódující části genomu některých polyomavirů, zejména BK virus, Virus JC, a SV40. Je nezbytný pro proliferaci virů, které jej exprimují, a předpokládá se, že se účastní regulace životního cyklu viru, zejména replikace a výstupu viru z hostitelské buňky, ale přesné mechanismy jsou nejasné.[32][33]
Taxonomie
Polyomaviry jsou členy skupiny I (dsDNA viry). Klasifikace polyomavirů byla předmětem několika navrhovaných revizí, když jsou objeveni noví členové skupiny. Dříve polyomaviry a papilomaviry, které sdílejí mnoho strukturálních rysů, ale mají velmi odlišné genomové organizace, byly společně zařazeny do dnes již zastaralé rodiny Papovaviridae.[34] (Název Papovaviridae odvozeno ze tří zkratek: Pa pro Papilomavirus, Po pro Polyomavirusa Va pro „vakuování“.)[35] Polyomaviry byly rozděleny do tří hlavních klady (tj. geneticky příbuzné skupiny): kladu SV40, ptačího kladu a myšího polyomavirového kladu.[36] Následná navrhovaná reklasifikace ze strany Mezinárodní výbor pro taxonomii virů (ICTV) doporučil rozdělit rodinu Polyomaviridae do tří rodů:[37]
- Rod Orthopolyomavirus (druh druhu SV40 )
- Rod Wukipolyomavirus (druh druhu KI polyomavirus )
- Rod Avipolyomavirus (druh druhu Ptačí polyomavirus )
Současný klasifikační systém ICTV rozeznává čtyři rody a 102 druhů, z nichž devíti nelze přiřadit rod. Tento systém zachovává rozdíl mezi ptačími a savčími viry a seskupuje ptačí podskupinu do rodu Gamapolyomavirus.[3]
- Rod Alphapolyomavirus, zadejte druh Mus musculus polyomavirus 1 (myší polyomavirus)
- Rod Betapolyomavirus, zadejte druh Polyomavirus Macaca mulatta 1 (SV40)
- Rod Deltapolyomavirus, zadejte druh Lidský polyomavirus 6
- Rod Gamapolyomavirus, zadejte druh Aves polyomavirus 1
Následující druhy nejsou přiřazeny k rodu:[3]
- Polymavirus Ailuropoda melanoleuca 1
- Bos taurus polyomavirus 1
- Centropristis striata polyomavirus 1
- Polyphavirus Delphinus delphis 1
- Procyon lotor polyomavirus 2
- Rhynchobatus djiddensis polyomavirus 1
- Polyaravirus Sparus aurata 1
- Trematomus bernacchii polyomavirus 1
- Trematomus pennellii polyomavirus 1
Popis dalších virů probíhá. Patří mezi ně polyomavirus mořské vydry 1[38] a alpakový polyomavirus[39] Dalším virem je polyomavirus obrovská panda 1.[40] Další virus byl popsán u hlodavců sigmodontinu.[41] Další - polyomavirus rejska stromová 1 - byl popsán v rejska stromu.[42]
Lidské polyomaviry
Většina polyomavirů neinfikuje člověka. Z polyomavirů katalogizovaných od roku 2017 bylo u lidských hostitelů známo celkem 14.[4] Některé polyomaviry jsou však spojeny s lidskými chorobami, zejména v imunokompromitovaný Jednotlivci. MCV je vysoce odlišný od ostatních lidských polyomavirů a nejvíce se vztahuje k myšímu polyomaviru. Polychodavirus spojený s Trichodysplasia spinulosa (TSV) je vzdáleně příbuzný MCV. Dva viry - HPyV6 a HPyV7 - se nejvíce podobají virům KI a WU, zatímco HPyV9 se nejvíce podobá lymfotropnímu polyomaviru (LPV) odvozenému z afrických zelených opic.
Byl popsán čtrnáctý virus.[43] Lyon IARC polyomavirus souvisí s mývalím polyomavirem.
Seznam lidských polyomavirů
Bylo identifikováno a mělo jejich 14 polyomavirů s lidskými hostiteli genomy seřazeno od roku 2017:[4]
| Druh | Navrhovaný rod | Název viru | Zkratka | NCBI RefSeq | Rok objevu | Klinická korelace (pokud existuje) | Reference |
|---|---|---|---|---|---|---|---|
| Lidský polyomavirus 5 | Alfa | Polyomavirus z Merkelových buněk | MCPyV | NC_010277 | 2008 | Rakovina Merkelových buněk[5] | [44][11][45] |
| Lidský polyomavirus 8 | Alfa | Trichodysplasia spinulosa polyomavirus | TSPyV | NC_014361 | 2010 | Trichodysplasia spinulosa[5] | [46][47] |
| Lidský polyomavirus 9 | Alfa | Lidský polyomavirus 9 | HPyV9 | NC_015150 | 2011 | Nejsou známy | [48] |
| Lidský polyomavirus 12 | Alfa | Lidský polyomavirus 12 | HPyV12 | NC_020890 | 2013 | Nejsou známy | [49] |
| Lidský polyomavirus 13 | Alfa | New Jersey polyomavirus | NJPyV | NC_024118 | 2014 | Nejsou známy | [50] |
| Lidský polyomavirus 1 | Beta | BK polyomavirus | BKPyV | NC_001538 | 1971 | Související s polyomavirem nefropatie; hemoragická cystitida[5] | [51] |
| Lidský polyomavirus 2 | Beta | JC polyomavirus | JCPyV | NC_001699 | 1971 | Progresivní multifokální leukoencefalopatie[5] | [52] |
| Lidský polyomavirus 3 | Beta | KI polyomavirus | KIPyV | NC_009238 | 2007 | Nejsou známy | [53] |
| Lidský polyomavirus 4 | Beta | WU polyomavirus | WUPyV | NC_009539 | 2007 | Nejsou známy | [14] |
| Lidský polyomavirus 6 | Delta | Lidský polyomavirus 6 | HPyV6 | NC_014406 | 2010 | Pruritická a dyskeratotická dermatóza spojená s HPyV6 (H6PD)[54] | [31] |
| Lidský polyomavirus 7 | Delta | Lidský polyomavirus 7 | HPyV7 | NC_014407 | 2010 | Hyperplázie epitelu související s HPyV7[54][55][56] | [31] |
| Lidský polyomavirus 10 | Delta | MW polyomavirus | MWPyV | NC_018102 | 2012 | Nejsou známy | [57][58][59] |
| Lidský polyomavirus 11 | Delta | STL polyomavirus | STLPyV | NC_020106 | 2013 | Nejsou známy | [60] |
| Lidský polyomavirus 14 | Alfa | Lyon IARC polyomavirus | LIPyV | NC_034253.1 | 2017 | Nejsou známy | [61][62] |
Deltapolyomavirus obsahuje pouze čtyři lidské viry uvedené v tabulce výše, přičemž jako typový typ obsahuje lidský polyomavirus 6. Skupiny Alpha a Beta obsahují viry, které infikují různé savce. Skupina Gamma obsahuje ptačí viry.[4] Klinicky významné asociace nemocí jsou zobrazeny pouze tam, kde se očekává kauzalita.[5][63]
U lidí byly detekovány protilátky proti opičímu lymfotropnímu polyomaviru, což naznačuje, že tento virus - nebo blízce příbuzný virus - může infikovat člověka.[64]
Klinický význam
Všechny polyomaviry jsou velmi časté infekce u dětí a mladých dospělých.[65] Zdá se, že většina z těchto infekcí způsobuje malé nebo žádné příznaky. Tyto viry jsou pravděpodobně celoživotně přetrvávající téměř u všech dospělých. Nejčastěji se vyskytují nemoci způsobené lidskými polyomavirovými infekcemi imunokompromitovaný lidé; sdružení nemocí zahrnují BK virus s nefropatie v transplantace ledvin a nerenální pacienti po transplantaci pevných orgánů,[8][9] Virus JC s progresivní multifokální leukoencefalopatie,[10] a Virus Merkelových buněk (MCV) s Rakovina Merkelových buněk.[11]
SV40
SV40 se replikuje v ledvinách opice aniž by způsoboval onemocnění, ale za laboratorních podmínek může u hlodavců způsobit rakovinu. V 50. a na počátku 60. let mohlo být SV40 vystaveno více než 100 milionů lidí kvůli dříve nezjištěné kontaminaci SV40 vakcína proti obrně, což vyvolalo obavy z možnosti, že by virus mohl způsobit onemocnění u lidí.[66][67] Ačkoli to bylo hlášeno jako přítomné u některých lidských rakovin, včetně mozkové nádory, kostní nádory, mezoteliomy, a non-Hodgkinovy lymfomy,[68] přesná detekce je často zmatena vysokou úrovní zkřížené reaktivity pro SV40 s rozšířenými lidskými polyomaviry.[67] Většina virologů odmítá SV40 jako příčinu lidských rakovin.[66][69][70]
Diagnóza
Diagnóza polyomaviru se téměř vždy vyskytuje po primární infekci, protože je asymptomatická nebo subklinická. Protilátkové testy se běžně používají k detekci přítomnosti protilátek proti jednotlivým virům.[71] Pro rozlišení mezi vysoce podobnými polyomaviry jsou často zapotřebí kompetitivní testy.[72]
V případech progresivní multifokální leukoencefalopatie (PML) se k přímému barvení tkání na přítomnost T antigenu viru JC používá zkříženě reaktivní protilátka proti SV40 T antigenu (obvykle Pab419). PCR lze použít na biopsii tkáně nebo mozkomíšní mok k amplifikaci polyomavirové DNA. To umožňuje nejen detekci polyomaviru, ale také o jaký podtyp jde.[73]
K diagnostice reaktivace polyomaviru u polyomavirové nefropatie (PVN) se používají tři hlavní diagnostické techniky: cytologie moči, kvantifikace virové zátěže v moči i v krvi a renální biopsie.[71]Reaktivace polyomaviru v ledvinách a močových cestách způsobuje vylučování infikovaných buněk, virionů a / nebo virových proteinů v moči. To umožňuje cytologii moči vyšetřit tyto buňky, což v případě polyomavirové inkluze jádra diagnostikuje infekci.[74] Protože moč infikovaného jedince bude obsahovat viriony a / nebo virovou DNA, lze kvantifikaci virové zátěže provést pomocí PCR.[75] To platí také pro krev.
Renální biopsii lze také použít, pokud jsou dvě právě popsané metody neprůkazné nebo pokud je požadována specifická virová zátěž pro renální tkáň. Podobně jako u cytologie moči se ledvinové buňky zkoumají světelnou mikroskopií na polyomavirové začlenění jádra, jakož i na buněčnou lýzu a virové částice v extracelulární tekutině. Virová zátěž jako dříve se také měří pomocí PCR.[Citace je zapotřebí ]
Tkáňové barvení pomocí monoklonální protilátky proti MCV T antigenu ukazuje užitečnost při diferenciaci karcinomu Merkelových buněk od jiných malých, kulatých buněčných nádorů.[76] Byly vyvinuty krevní testy k detekci protilátek proti MCV, které ukazují, že infekce virem je velmi rozšířená, ačkoli pacienti s karcinomem Merkelových buněk mají výjimečně vyšší protilátkové odpovědi než asymptomaticky infikovaní lidé.[7][77][78][79]
Použití při sledování lidské migrace
Virus JC nabízí slibný genetický marker pro lidskou evoluci a migraci.[80] Nosí jej 70–90 procent lidí a obvykle se přenáší z rodičů na potomky. Tato metoda se nejeví jako spolehlivá pro sledování nedávný africký původ moderních lidí.
Dějiny
Myší polyomavirus byl první objevený polyomavirus, který ohlásil Ludwik Gross v roce 1953 jako extrakt z myši leukémie schopné vyvolat příušní žláza nádory.[81] Původce byl identifikován jako virus Sarah Stewart a Bernice Eddy, po kterém se kdysi nazýval „SE polyoma“.[82][83][84] Termín „polyom“ označuje schopnost virů produkovat za určitých podmínek mnohočetné (poly-) nádory (-oma). Název byl kritizován jako „bezmasý jazykový sendvič“ („bez masa“, protože obojí morfémy v „polyomu“ jsou přípony) poskytující malý vhled do biologie virů; ve skutečnosti následný výzkum zjistil, že většina polyomavirů zřídka způsobuje klinicky významné onemocnění v jejich hostitelských organismech za přirozených podmínek.[85]
Od roku 2017 byly identifikovány a sekvenovány desítky polyomavirů, které infikovaly hlavně ptáky a savce. Je známo, že dva polyomaviry infikují ryby, černý mořský okoun[86] a pražma mořská.[87] Je známo, že celkem čtrnáct polyomavirů infikuje člověka.[4]
Reference
- ^ Moens U, Calvignac-Spencer S, Lauber C, Ramqvist T, Feltkamp MC, Daugherty MD, Verschoor EJ, Ehlers B (červen 2017). „Profil taxonomie viru ICTV: Polyomaviridae“. The Journal of General Virology. 98 (6): 1159–1160. doi:10.1099 / jgv.0.000839. PMC 5656788. PMID 28640744.
- ^ A b C d E F G "Zpráva ICTV Polyomaviridae".
- ^ A b C „Virus Taxonomy: 2019 Release“. talk.ictvonline.org. Mezinárodní výbor pro taxonomii virů. Citováno 26. dubna 2020.
- ^ A b C d E Calvignac-Spencer S, Feltkamp MC, Daugherty MD, Moens U, Ramqvist T, Johne R, Ehlers B (červen 2016). „Aktualizace taxonomie pro rodinu Polyomaviridae“. Archivy virologie. 161 (6): 1739–50. doi:10.1007 / s00705-016-2794-r. PMID 26923930.
- ^ A b C d E F G h i j DeCaprio JA, Garcea RL (duben 2013). „Roh hojnosti lidských polyomavirů“. Recenze přírody. Mikrobiologie. 11 (4): 264–76. doi:10.1038 / nrmicro2992. PMC 3928796. PMID 23474680.
- ^ Gossai A, Waterboer T, Nelson HH, Michel A, Willhauck-Fleckenstein M, Farzan SF, Hoen AG, Christensen BC, Kelsey KT, Marsit CJ, Pawlita M, Karagas MR (leden 2016). "Seroepidemiologie lidských polyomavirů v populaci USA". American Journal of Epidemiology. 183 (1): 61–9. doi:10.1093 / aje / kwv155. PMC 5006224. PMID 26667254.
- ^ A b Kean JM, Rao S, Wang M, Garcea RL (březen 2009). "Seroepidemiologie lidských polyomavirů". PLOS patogeny. 5 (3): e1000363. doi:10.1371 / journal.ppat.1000363. PMC 2655709. PMID 19325891.
- ^ A b Jamboti JS (srpen 2016). „Nefropatie u viru BK u příjemců transplantovaných ledvin“. Nefrologie. 21 (8): 647–54. doi:10.1111 / nep.12728. PMID 26780694.
- ^ A b Kuppachi S, Kaur D, Holanda DG, Thomas CP (duben 2016). „Infekce virem BK polyomů a onemocnění ledvin při nerenální transplantaci pevných orgánů“. Klinický ledvinový deník. 9 (2): 310–8. doi:10.1093 / ckj / sfv143. PMC 4792618. PMID 26985385.
- ^ A b Adang L, Berger J (2015). „Progresivní multifokální leukoencefalopatie“. F1000Výzkum. 4: 1424. doi:10.12688 / F1000Research.7071.1. PMC 4754031. PMID 26918152.
- ^ A b C Feng H, Shuda M, Chang Y, Moore PS (únor 2008). „Klonální integrace polyomaviru do lidského karcinomu Merkelových buněk“. Věda. 319 (5866): 1096–100. Bibcode:2008Sci ... 319.1096F. doi:10.1126 / science.1152586. PMC 2740911. PMID 18202256.
- ^ Salunke DM, Caspar DL, Garcea RL (září 1986). "Vlastní sestavení purifikovaného polyomavirového kapsidového proteinu VP1". Buňka. 46 (6): 895–904. doi:10.1016/0092-8674(86)90071-1. PMID 3019556.
- ^ A b Gaynor AM, Nissen MD, Whiley DM, Mackay IM, Lambert SB, Wu G, Brennan DC, Storch GA, Sloots TP, Wang D (květen 2007). „Identifikace nového polyomaviru od pacientů s akutními infekcemi dýchacích cest“. PLOS patogeny. 3 (5): e64. doi:10.1371 / journal.ppat.0030064. PMC 1864993. PMID 17480120.
- ^ A b C d E F G h Mezinárodní agentura pro výzkum rakoviny (2013). "Úvod do polyomavirů" (PDF). Monografie IARC o hodnocení karcinogenních rizik pro člověka. 104: 121–131.
- ^ A b Buch MH, Liaci AM, O'Hara SD, Garcea RL, Neu U, Stehle T (říjen 2015). „Strukturální a funkční analýza proteinů kapsidy myšího polyomaviru potvrzuje determinanty rozpoznávání a patogenity ligandu“. PLOS patogeny. 11 (10): e1005104. doi:10.1371 / journal.ppat.1005104. PMC 4608799. PMID 26474293.
- ^ Schowalter RM, Pastrana DV, Buck CB (červenec 2011). „Glykosaminoglykany a sialylované glykany postupně usnadňují infekční vstup polyomaviru z Merkelových buněk“. PLOS patogeny. 7 (7): e1002161. doi:10.1371 / journal.ppat.1002161. PMC 3145800. PMID 21829355.
- ^ A b Inoue T, Tsai B (leden 2013). „Jak viry používají endoplazmatické retikulum pro vstup, replikaci a sestavení“. Perspektivy Cold Spring Harbor v biologii. 5 (1): a013250. doi:10.1101 / cshperspect.a013250. PMC 3579393. PMID 23284050.
- ^ A b C d Gjoerup O, Chang Y (2010). "Aktualizace lidských polyomavirů a rakoviny". Pokroky ve výzkumu rakoviny. 106: 1–51. doi:10.1016 / S0065-230X (10) 06001-X. ISBN 9780123747716. PMID 20399955.
- ^ Andrabi S, Hwang JH, Choe JK, Roberts TM, Schaffhausen BS (2011). „Srovnání mezi myším polyomavirem a opičím virem 40 vykazuje významné rozdíly ve funkci malého antigenu T“. Journal of Virology. 85 (20): 10649–10658. doi:10.1128 / JVI.05034-11. PMC 3187521. PMID 21835797.
- ^ Rotondo JC, Bononi I, Puozzo A, Govoni M, Foschi V, Lanza G, Gafà R, Gaboriaud P, Touzé FA, Selvatici R, Martini F, Tognon M (červenec 2017). „Karcinomy z Merkelových buněk vznikající u pacientů s postižením autoimunitních onemocnění léčených biologickými léky, včetně anti-TNF“. Klinický výzkum rakoviny. 23 (14): 3929–3934. doi:10.1158 / 1078-0432.CCR-16-2899. PMID 28174236.
- ^ White MK, Gordon J, Reiss K, Del Valle L, Croul S, Giordano A, Darbinyan A, Khalili K (prosinec 2005). "Lidské polyomaviry a mozkové nádory". Výzkum mozku. Recenze výzkumu mozku. 50 (1): 69–85. doi:10.1016 / j.brainresrev.2005.04.007. PMID 15982744.
- ^ Kazem S, van der Meijden E, Wang RC, Rosenberg AS, Pope E, Benoit T, Fleckman P, Feltkamp MC (2014). „Trichodysplasia spinulosa spojená s polyomavirem zahrnuje hyperproliferaci, fosforylaci pRB a upregulaci p16 a p21“. PLOS One. 9 (10): e108947. Bibcode:2014PLoSO ... 9j8947K. doi:10.1371 / journal.pone.0108947. PMC 4188587. PMID 25291363.
- ^ Kelley WL, Georgopoulos C (duben 1997). „T / t běžný exon opičích virů 40, JC a BK polyomavirových T antigenů může funkčně nahradit J-doménu molekulárního chaperonu Escherichia coli DnaJ“. Sborník Národní akademie věd Spojených států amerických. 94 (8): 3679–84. Bibcode:1997PNAS ... 94,3679 tis. doi:10.1073 / pnas.94.8.3679. PMC 20500. PMID 9108037.
- ^ Pallas DC, Shahrik LK, Martin BL, Jaspers S, Miller TB, Brautigan DL, Roberts TM (leden 1990). „Polyoma malé a střední T antigeny a SV40 malý t antigen tvoří stabilní komplexy s proteinovou fosfatázou 2A“. Buňka. 60 (1): 167–76. doi:10.1016 / 0092-8674 (90) 90726-u. PMID 2153055.
- ^ Sontag E, Fedorov S, Kamibayashi C, Robbins D, Cobb M, Mumby M (prosinec 1993). „Interakce malého nádoru antigenu SV40 s proteinovou fosfatázou 2A stimuluje dráhu kinázy mapy a indukuje buněčnou proliferaci“. Buňka. 75 (5): 887–97. doi:10.1016 / 0092-8674 (93) 90533-V. PMID 8252625.
- ^ Watanabe G, Howe A, Lee RJ, Albanese C, Shu IW, Karnezis AN, Zon L, Kyriakis J, Rundell K, Pestell RG (listopad 1996). "Indukce cyklinu D1 opičím virem 40 malým nádorovým antigenem". Sborník Národní akademie věd Spojených států amerických. 93 (23): 12861–6. Bibcode:1996PNAS ... 9312861W. doi:10.1073 / pnas.93.23.12861. PMC 24011. PMID 8917510.
- ^ Kwun HJ, Shuda M, Feng H, Camacho CJ, Moore PS, Chang Y (srpen 2013). „Malý T antigen polyomaviru z Merkelových buněk kontroluje replikaci viru a expresi onkoproteinu zaměřením na buněčnou ubikvitin ligázu SCFFbw7“. Mobilní hostitel a mikrob. 14 (2): 125–35. doi:10.1016 / j.chom.2013.06.008. PMC 3764649. PMID 23954152.
- ^ Shuda M, Kwun HJ, Feng H, Chang Y, Moore PS (září 2011). „Malý T antigen lidské buňky Merkel z polyomaviru je onkoprotein zaměřený na regulátor translace 4E-BP1“. The Journal of Clinical Investigation. 121 (9): 3623–34. doi:10,1172 / JCI46323. PMC 3163959. PMID 21841310.
- ^ Chen XS, Stehle T, Harrison SC (červen 1998). „Interakce interního proteinu polyomaviru VP2 s hlavním kapsidovým proteinem VP1 a důsledky pro účast VP2 na vstupu viru“. Časopis EMBO. 17 (12): 3233–40. doi:10.1093 / emboj / 17.12.3233. PMC 1170661. PMID 9628860.
- ^ A b C Schowalter RM, Pastrana DV, Pumphrey KA, Moyer AL, Buck CB (červen 2010). „Polyomavirus z Merkelových buněk a dva dříve neznámé polyomaviry jsou chronicky vylučovány z lidské kůže“. Mobilní hostitel a mikrob. 7 (6): 509–15. doi:10.1016 / j.chom.2010.05.006. PMC 2919322. PMID 20542254.
- ^ Sariyer IK, Saribas AS, White MK, Safak M (květen 2011). „Infekce agomprotein-negativními mutanty polyomaviru JC a SV40 vede k uvolňování virionů, které mají většinou nedostatek obsahu DNA“. Virology Journal. 8: 255. doi:10.1186 / 1743-422X-8-255. PMC 3127838. PMID 21609431.
- ^ Saribas AS, Coric P, Hamazaspyan A, Davis W, Axman R, White MK, Abou-Gharbia M, Childers W, Condra JH, Bouaziz S, Safak M (říjen 2016). „Vznikající z neznáma: strukturní a funkční vlastnosti agnoproteinu polyomavirů“. Journal of Cellular Physiology. 231 (10): 2115–27. doi:10,1002 / jcp.25329. PMC 5217748. PMID 26831433.
- ^ „Webové stránky taxonomie ICTV“.
- ^ Mezinárodní agentura pro výzkum rakoviny (2013). „Pracovní skupina IARC pro hodnocení karcinogenních rizik pro člověka. Malárie a některé polyomaviry (viry SV40, BK, JC a Merkelovy buňky)“. Monografie IARC o hodnocení karcinogenních rizik pro člověka. 104.
- ^ Pérez-Losada M, Christensen RG, McClellan DA, Adams BJ, Viscidi RP, Demma JC, Crandall KA (červen 2006). „Srovnání fylogenetické společné divergence mezi polyomaviry a jejich hostiteli“. Journal of Virology. 80 (12): 5663–9. doi:10.1128 / JVI.00056-06. PMC 1472594. PMID 16731904.
- ^ Johne R, Buck CB, Allander T, Atwood WJ, Garcea RL, Imperiale MJ, Major EO, Ramqvist T, Norkin LC (září 2011). "Taxonomický vývoj v rodině Polyomaviridae". Archivy virologie. 156 (9): 1627–34. doi:10.1007 / s00705-011-1008-x. PMC 3815707. PMID 21562881.
- ^ Siqueira JD, Ng TF, Miller M, Li L, Deng X, Dodd E, Batac F, Delwart E (červenec 2017). "Endemická infekce uvízlých vydry jižní (Enhydra lutris nereis) novým parvovirem, polyomavirem a adenovirem". Journal of Wildlife Diseases. 53 (3): 532–542. doi:10.7589/2016-04-082. PMID 28192039.
- ^ Dela Cruz FN, Li L, Delwart E, Pesavento PA (2017). "Nový plicní polyomavirus v alpakách (Vicugna pacos)". Veterinární mikrobiologie. 201: 49–55. doi:10.1016 / j.vetmic.2017.01.005. PMID 28284622.
- ^ Qi D, Shan T, Liu Z, Deng X, Zhang Z, Bi W, Owens JR, Feng F, Zheng L, Huang F, Delwart E, Hou R, Zhang W (2017). „Nový polyomavirus z nosní dutiny pandy obrovské (Ailuropoda melanoleuca)“. Virology Journal. 14 (1): 207. doi:10.1186 / s12985-017-0867-5. PMC 5658932. PMID 29078783.
- ^ Gonçalves Motta Maia F, Marciel de Souza W, Sabino-Santos G, Jorge Fumagalli M, Modha S, Ramiro Murcia P, Tadeu Moraes Figueiredo L (2018). "Nový polyomavirus u sigmodontinových hlodavců ze státu São Paulo v Brazílii". Archivy virologie. 163 (10): 2913–2915. doi:10.1007 / s00705-018-3913-8. PMID 29931397.
- ^ Liu P, Qiu Y, Xing C, Zhou JH, Yang WH, Wang Q, Li JY, Han X, Zhang YZ, Ge XY (2019). "Detekce a genomová charakterizace dvou nových papilomavirů a nového polyomaviru v rejska stromu (Tupaia belangeri chinensis) v Číně". Virol J. 16 (1): 35. doi:10.1186 / s12985-019-1141-9. PMC 6423848. PMID 30885224.
- ^ Gheit T, Dutta S, Oliver J, Robitaille A, Hampras S, Combes JD, McKay-Chopin S, Le Calvez-Kelm F, Fenske N, Cherpelis B, Giuliano AR, Franceschi S, McKay J, Rollison DE, Tommasino M ( 2017). „Izolace a charakterizace nového domnělého lidského polyomaviru“. Virologie. 506: 45–54. doi:10.1016 / j.virol.2017.03.007. PMID 28342387.
- ^ Altman LK (18. ledna 2008). „Virus souvisí s výkonným karcinomem kůže“. The New York Times. Citováno 18. ledna 2008.
- ^ Shuda M, Arora R, Kwun HJ, Feng H, Sarid R, Fernández-Figueras MT, Tolstov Y, Gjoerup O, Mansukhani MM, Swerdlow SH, Chaudhary PM, Kirkwood JM, Nalesnik MA, Kant JA, Weiss LM, Moore PS, Chang Y (září 2009). „Infekce polyomaviry lidských Merkelových buněk I. Exprese antigenu MCV T v karcinomu Merkelových buněk, lymfoidních tkáních a lymfoidních nádorech“. International Journal of Cancer. 125 (6): 1243–9. doi:10.1002 / ijc.24510. PMC 6388400. PMID 19499546.
- ^ van der Meijden E, Janssens RW, Lauber C, Bouwes Bavinck JN, Gorbalenya AE, Feltkamp MC (červenec 2010). „Objev nového lidského polyomaviru spojeného s trichodysplasia spinulosa u imunokompromitovaného pacienta“. PLOS patogeny. 6 (7): e1001024. doi:10.1371 / journal.ppat.1001024. PMC 2912394. PMID 20686659.
- ^ Kazem S, van der Meijden E, Feltkamp MC (srpen 2013). „Polyomavirus související s trichodysplasia spinulosa: virologické pozadí a klinické důsledky“. APMIS. 121 (8): 770–82. doi:10,1111 / přibližně 12092. PMID 23593936.
- ^ Scuda N, Hofmann J, Calvignac-Spencer S, Ruprecht K, Liman P, Kühn J, Hengel H, Ehlers B (květen 2011). „Nový lidský polyomavirus, který úzce souvisí s lymfotropním polyomavirem pocházejícím z africké zelené opice“. Journal of Virology. 85 (9): 4586–90. doi:10.1128 / jvi.02602-10. PMC 3126223. PMID 21307194.
- ^ Korup S, Rietscher J, Calvignac-Spencer S, Trusch F, Hofmann J, Moens U, Sauer I, Voigt S, Schmuck R, Ehlers B (2013). „Identifikace nového lidského polyomaviru v orgánech gastrointestinálního traktu“. PLOS One. 8 (3): e58021. Bibcode:2013PLoSO ... 858021K. doi:10.1371 / journal.pone.0058021. PMC 3596337. PMID 23516426.
- ^ Mishra N, Pereira M, Rhodes RH, An P, Pipas JM „Jain K, Kapoor A, Briese T, Faust PL, Lipkin WI (listopad 2014). „Identifikace nového polyomaviru u příjemce transplantace pankreatu s retinální slepotou a vaskulitickou myopatií“. The Journal of Infectious Diseases. 210 (10): 1595–9. doi:10.1093 / infdis / jiu250. PMC 4334791. PMID 24795478.
- ^ Gardner SD, Field AM, Coleman DV, Hulme B (červen 1971). „Nový lidský papovavirus (B.K.) izolovaný z moči po transplantaci ledvin“. Lanceta. 1 (7712): 1253–7. doi:10.1016 / s0140-6736 (71) 91776-4. PMID 4104714.
- ^ Padgett BL, Walker DL, ZuRhein GM, Eckroade RJ, Dessel BH (červen 1971). "Pěstování viru podobného papově z lidského mozku s progresivní multifokální leukoencefalopatií". Lanceta. 1 (7712): 1257–60. doi:10.1016 / S0140-6736 (71) 91777-6. PMID 4104715.
- ^ Allander T, Andreasson K, Gupta S, Bjerkner A, Bogdanovic G, Persson MA, Dalianis T, Ramqvist T, Andersson B (duben 2007). „Identifikace třetího lidského polyomaviru“. Journal of Virology. 81 (8): 4130–6. doi:10.1128 / JVI.00028-07. PMC 1866148. PMID 17287263.
- ^ A b Nguyen KD, Lee EE, Yue Y, Stork J, Pock L, North JP, Vandergriff T, Cockerell C, Hosler GA, Pastrana DV, Buck CB, Wang RC (květen 2017). „Lidský polyomavirus 6 a 7 je spojen s pruritickými a dyskeratotickými dermatózami“. Journal of the American Academy of Dermatology. 76 (5): 932–940.e3. doi:10.1016 / j.jaad.2016.11.035. PMC 5392424. PMID 28040372.
- ^ Ho J, Jedrych JJ, Feng H, Natalie AA, Grandinetti L, Mirvish E, Crespo MM, Yadav D, Fasanella KE, Proksell S, Kuan SF, Pastrana DV, Buck CB, Shuda Y, Moore PS, Chang Y (květen 2015 ). „Pruritická vyrážka spojená s lidským polyomavirem 7 a virémie u příjemců transplantátu“. The Journal of Infectious Diseases. 211 (10): 1560–5. doi:10.1093 / infdis / jiu524. PMC 4425822. PMID 25231015.
- ^ Toptan T, Yousem SA, Ho J, Matsushima Y, Stabile LP, Fernández-Figueras MT, Bhargava R, Ryo A, Moore PS, Chang Y (únor 2016). „Průzkum lidských polyomavirů u rakoviny“. JCI Insight. 1 (2). doi:10.1172 / jci.insight.85562. PMC 4811373. PMID 27034991.
- ^ Siebrasse EA, Reyes A, Lim ES, Zhao G, Mkakosya RS, Manary MJ, Gordon JI, Wang D (říjen 2012). „Identifikace MW polyomaviru, nového polyomaviru v lidské stolici“. Journal of Virology. 86 (19): 10321–6. doi:10.1128 / JVI.01210-12. PMC 3457274. PMID 22740408.
- ^ Buck CB, Phan GQ, Raiji MT, Murphy PM, McDermott DH, McBride AA (říjen 2012). „Kompletní genomová sekvence desátého lidského polyomaviru“. Journal of Virology. 86 (19): 10887. doi:10.1128 / JVI.01690-12. PMC 3457262. PMID 22966183.
- ^ Yu G, Greninger AL, Isa P, Phan TG, Martínez MA, de la Luz Sanchez M, Contreras JF, Santos-Preciado JI, Parsonnet J, Miller S, DeRisi JL, Delwart E, Arias CF, Chiu CY (2012). „Objev nového polyomaviru v akutních průjmových vzorcích od dětí“. PLOS One. 7 (11): e49449. Bibcode:2012PLoSO ... 749449Y. doi:10.1371 / journal.pone.0049449. PMC 3498111. PMID 23166671.
- ^ Lim ES, Reyes A, Antonio M, Saha D, Ikumapayi UN, Adeyemi M, Stine OC, Skelton R, Brennan DC, Mkakosya RS, Manary MJ, Gordon JI, Wang D (únor 2013). „Objev STL polyomaviru, polyomaviru původního rekombinantního původu, který kóduje jedinečný T antigen alternativním sestřihem“. Virologie. 436 (2): 295–303. doi:10.1016 / j.virol.2012.12.005. PMC 3693558. PMID 23276405.
- ^ Gheit, Tarik; Dutta, Sankhadeep; Oliver, Javier; Robitaille, Alexis; Hampras, Shalaka; Combes, Jean-Damien; McKay-Chopin, Sandrine; Calvez-Kelm, Florence Le; Fenske, Neil (2017). „Izolace a charakterizace nového domnělého lidského polyomaviru“. Virologie. 506: 45–54. doi:10.1016 / j.virol.2017.03.007. PMID 28342387.
- ^ Prado JC, Monezi TA, Amorim AT, Lino V, Paladino A, Boccardo E (2018). „Lidské polyomaviry a rakovina: přehled“. Kliniky (Sao Paulo). 73 (doplněk 1): e558s. doi:10,6061 / kliniky / 2018 / e558s. PMC 6157077. PMID 30328951.
- ^ Dalianis T, Hirsch HH (březen 2013). „Lidské polyomaviry v nemoci a rakovině“. Virologie. 437 (2): 63–72. doi:10.1016 / j.virol.2012.12.015. PMID 23357733.
- ^ Van Ghelue M, Khan MT, Ehlers B, Moens U (listopad 2012). "Analýza genomu nových lidských polyomavirů". Recenze v lékařské virologii. 22 (6): 354–77. doi:10,1002 / rmv.1711. PMID 22461085.
- ^ Egli A, Infanti L, Dumoulin A, Buser A, Samaridis J, Stebler C, Gosert R, Hirsch HH (březen 2009). „Prevalence infekce a replikace polyomaviru BK a JC u 400 zdravých dárců krve“. The Journal of Infectious Diseases. 199 (6): 837–46. doi:10.1086/597126. PMID 19434930.
- ^ A b Poulin DL, DeCaprio JA (září 2006). "Existuje role pro SV40 u lidské rakoviny?". Journal of Clinical Oncology. 24 (26): 4356–65. doi:10.1200 / JCO.2005.03.7101. PMID 16963733.
- ^ A b zur Hausen H (prosinec 2003). „SV40 u lidských rakovin - nekonečný příběh?“. International Journal of Cancer. 107 (5): 687. doi:10.1002 / ijc.11517. PMID 14566815.
- ^ Gazdar AF, Butel JS, Carbone M (prosinec 2002). „SV40 a lidské nádory: mýtus, asociace nebo kauzalita?“. Recenze přírody. Rakovina. 2 (12): 957–64. doi:10.1038 / nrc947. PMID 12459734.
- ^ Carroll-Pankhurst C, Engels EA, Strickler HD, Goedert JJ, Wagner J, Mortimer EA (listopad 2001). „Třicet pětiletá úmrtnost po obdržení vakcíny proti dětské obrně kontaminované SV40 během novorozeneckého období“. British Journal of Cancer. 85 (9): 1295–7. doi:10.1054 / bjoc.2001.2065. PMC 2375249. PMID 11720463.
- ^ Shah KV (leden 2007). „SV40 a lidská rakovina: přehled nedávných údajů“. International Journal of Cancer. 120 (2): 215–23. doi:10.1002 / ijc.22425. PMID 17131333.
- ^ A b Drachenberg CB, Hirsch HH, Ramos E, Papadimitriou JC (prosinec 2005). „Polyomavirové onemocnění při transplantaci ledvin: přehled patologických nálezů a diagnostických metod“. Lidská patologie. 36 (12): 1245–55. doi:10.1016 / j.humpath.2005.08.009. PMID 16311117.
- ^ Viscidi RP, Clayman B (2006). "Sérologická zkřížená reaktivita mezi kapsulemi polyomaviru". V Ahsan N (ed.). Polyomaviry a lidské nemoci. Pokroky v experimentální medicíně a biologii. 577. str. 73–84. doi:10.1007/0-387-32957-9_5. ISBN 978-0-387-29233-5. PMID 16626028.
- ^ Drews K, Bashir T, Dörries K (leden 2000). "Kvantifikace lidského polyomaviru JC v mozkové tkáni a mozkomíšním moku pacientů s progresivní multifokální leukoencefalopatií kompetitivní PCR". Journal of Virological Methods. 84 (1): 23–36. doi:10.1016 / S0166-0934 (99) 00128-7. PMID 10644084.
- ^ Nickeleit V, Hirsch HH, Binet IF, Gudat F, Prince O, Dalquen P, Thiel G, Mihatsch MJ (květen 1999). „Polyomavirová infekce u příjemců ledvinových aloštěpů: od latentní infekce po manifestní onemocnění“. Časopis Americké nefrologické společnosti. 10 (5): 1080–9. PMID 10232695.
- ^ Randhawa PS, Vats A, Zygmunt D, Swalsky P, Scantlebury V, Shapiro R, Finkelstein S (srpen 2002). "Kvantifikace virové DNA v tkáni ledvinového aloštěpu od pacientů s nefropatií viru BK". Transplantace. 74 (4): 485–8. doi:10.1097/00007890-200208270-00009. PMID 12352906.
- ^ Busam KJ, Jungbluth AA, Rekthman N, Coit D, Pulitzer M, Bini J, Arora R, Hanson NC, Tassello JA, Frosina D, Moore P, Chang Y (září 2009). „Exprese polyomaviru z Merkelových buněk v karcinomech z Merkelových buněk a jeho absence v kombinovaných nádorech a plicních neuroendokrinních karcinomech“. American Journal of Surgical Pathology. 33 (9): 1378–85. doi:10.1097 / PAS.0b013e3181aa30a5. PMC 2932664. PMID 19609205.
- ^ Tolstov YL, Pastrana DV, Feng H, Becker JC, Jenkins FJ, Moschos S, Chang Y, Buck CB, Moore PS (září 2009). „Infekce polyomavirem z lidských Merkelových buněk II. MCV je běžná infekce člověka, kterou lze detekovat konformačními kapsidovými epitopovými imunotesty“. International Journal of Cancer. 125 (6): 1250–6. doi:10.1002 / ijc.24509. PMC 2747737. PMID 19499548.
- ^ Pastrana DV, Tolstov YL, Becker JC, Moore PS, Chang Y, Buck CB (září 2009). „Kvantifikace lidské seroresponzivity na polyomavirus z Merkelových buněk“. PLOS patogeny. 5 (9): e1000578. doi:10.1371 / journal.ppat.1000578. PMC 2734180. PMID 19750217.
- ^ Carter JJ, Paulson KG, Wipf GC, Miranda D, Madeleine MM, Johnson LG, Lemos BD, Lee S, Warcola AH, Iyer JG, Nghiem P, Galloway DA (listopad 2009). „Sdružení protilátek specifických pro polyomaviry z Merkelových buněk s karcinomem z Merkelových buněk“. Journal of the National Cancer Institute. 101 (21): 1510–22. doi:10.1093 / jnci / djp332. PMC 2773184. PMID 19776382.
- ^ Elizabeth Matisoo-Smith; K. Ann Horsburgh (2012). DNA pro archeology. Routledge. ISBN 978-1598746815.
- ^ Gross L (červen 1953). „Filtrovatelné činidlo získané z leukemických extraktů Ak, které způsobuje karcinomy slinných žláz u myší C3H“. Sborník Společnosti pro experimentální biologii a medicínu. 83 (2): 414–21. doi:10.3181/00379727-83-20376. PMID 13064287.
- ^ Stewart SE, Eddy BE, Borgese N (červen 1958). "Novotvary u myší inokulovaných nádorovým činidlem prováděným v tkáňové kultuře". Journal of the National Cancer Institute. 20 (6): 1223–43. doi:10.1093 / jnci / 20.6.1223. PMID 13549981.
- ^ Eddy BE, Stewart SE (listopad 1959). "Vlastnosti viru SE polyomu". American Journal of Public Health and the Nation's Health. 49 (11): 1486–92. doi:10,2105 / AJPH.49.11.1486. PMC 1373056. PMID 13819251.
- ^ Schowalter RM, Pastrana DV, Buck CB (červenec 2011). „Glykosaminoglykany a sialylované glykany postupně usnadňují infekční vstup polyomaviru z Merkelových buněk“. PLOS patogeny. 7 (7): e1002161. doi:10.1371 / journal.ppat.1002161. PMC 3145800. PMID 21829355.
- ^ Gottlieb KA, Villarreal LP (červen 2001). "Přírodní biologie polyomavirového středního T antigenu". Recenze mikrobiologie a molekulární biologie. 65 (2): 288–318, druhá a třetí strana, obsah. doi:10.1128 / mmbr. 65.2.288-318.2001. PMC 99028. PMID 11381103.
- ^ Peretti A, FitzGerald PC, Bliskovsky V, Pastrana DV, Buck CB (leden 2015). "Genome Sequence of a Fish-Associated Polyomavirus, Black Sea Bass (Centropristis striata) Polyomavirus 1". Oznámení o genomu. 3 (1): e01476-14. doi:10.1128 / genomA.01476-14. PMC 4319505. PMID 25635011.
- ^ López-Bueno A, Mavian C, Labella AM, Castro D, Borrego JJ, Alcami A, Alejo A (říjen 2016). „Souběh iridoviru, polyomaviru a jedinečného člena nové skupiny rybích papilomavirů v lykožrutu postiženém mořskou vlnou“. Journal of Virology. 90 (19): 8768–79. doi:10.1128 / JVI.01369-16. PMC 5021401. PMID 27440877.