Stimulátor genů interferonu - Stimulator of interferon genes





Stimulátor genů interferonu (BODNUTÍ), také známý jako transmembránový protein 173 (TMEM173) a MPYS/MITA/ERIS je protein že u lidí je kódováno STING1 gen.[5]
STING hraje důležitou roli v imunita. STING indukuje interferon typu I. produkce, když jsou buňky infikovány intracelulárními patogeny, jako je např viry, mykobakterie a intracelulární paraziti.[6] Interferon typu I., zprostředkovaný STING, chrání infikované buňky a blízké buňky před lokální infekcí vazbou na stejnou buňku, která ji vylučuje (autokrinní signalizace) a blízké buňky (parakrin signalizace.)
STING funguje jako přímý cytosolický DNA senzor (CDS) a an adaptační protein v Interferon typu I. signalizace prostřednictvím různých molekulárních mechanismů. Ukázalo se, že aktivuje následné transkripční faktory STAT6 a IRF3 přes TBK1, které jsou odpovědné za antivirovou reakci a vrozenou imunitní reakci proti intracelulární patogen.[7]
Struktura

Mezi aminokyseliny 1–379 lidského STING patří 4 transmembránové oblasti (TM) a a C-koncová doména. The C-koncová doména (CTD: aminokyseliny 138–379) obsahuje dimerizační doménu (DD) a karboxy-koncový ocas (CTT: aminokyseliny 340–379).[7]
STING tvoří v buňce symetrický dimer. Dimer STING připomíná motýla s hlubokou rozštěpem mezi dvěma protomery. The hydrofobní zbytky z každé formy STING protomeru hydrofobní interakce mezi sebou na rozhraní.[7][8]
Výraz
STING je vyjádřeno v krvetvorné buňky v periferní lymfoidní tkáně, počítaje v to T lymfocyty, NK buňky, myeloidní buňky a monocyty. Bylo také prokázáno, že STING je vysoce exprimován v plicích, vaječník, srdce, hladký sval, sítnice, kostní dřeň a pochvy.[9][10]
Lokalizace
Subcelulární lokalizace STING byla objasněna jako endoplazmatické retikulum protein. Je také pravděpodobné, že se STING sdružuje v těsné blízkosti s ER membrána asociovaná s mitochondrií (MAM) - rozhraní mezi mitochondrií a ER.[11] Během intracelulární infekce je STING schopen přemístit se z endoplazmatické retikulum na perinukleární vezikuly potenciálně zapojen do exocyst zprostředkovaný transport.[11] Bylo také prokázáno, že STING se kolokalizuje autofagickými proteiny, protein spojený s mikrotubuly 1 lehký řetězec 3 (LC3) a protein 9A související s autofagií, po stimulaci dvouvláknovou DNA, což naznačuje její přítomnost v autofagozom.[12]
Funkce
STING zprostředkovává interferon typu I. produkce v reakci na intracelulární DNA a různé intracelulární patogeny, včetně viry, intracelulární bakterie a intracelulární paraziti.[13] Po infekci může STING z infikovaných buněk cítit přítomnost nukleové kyseliny z intracelulárních patogenů a poté vyvolat interferon β a více než 10 forem interferon α Výroba. Interferon typu I. produkované infikovanými buňkami lze najít a vázat se na ně Receptor interferonu alfa / beta blízkých buněk k ochraně buněk před lokální infekcí.
Antivirová imunita
STING vyvolává silné imunita vůči interferonu typu I. proti virové infekci. Po virový vstup virové nukleové kyseliny bude přítomen v cytosolu infikovaných buněk. Několik senzorů DNA, jako např DAI, RNA polymeráza III, IFI16, DDX41 a cGAS, může detekovat cizí nukleové kyseliny. Po rozpoznání virové DNA iniciují senzory DNA následné signální dráhy aktivací interferonové reakce zprostředkované STING.[14]
Adenovirus, virus herpes simplex, HSV-1 a HSV-2, jakož i negativní RNA virus -virus vezikulární stomatitidy (VSV) Bylo prokázáno, že jsou schopné aktivovat STING-závislé vrozená imunitní odpověď.[13]
Nedostatek STING u myší vedl k letální náchylnosti k infekci HSV-1 v důsledku absence úspěšné reakce na interferon typu I.[15]
Bodová mutace serinu S358 tlumí aktivaci STING-IFN u netopýrů a navrhuje se, aby netopýrům poskytl jejich schopnost hostitele rezervoáru.[16]
Proti intracelulárním bakteriím
Intracelulární bakterie, Listeria monocytogenes Bylo prokázáno, že stimulují imunitní odpověď hostitele pomocí STING.[17] STING může hrát důležitou roli při výrobě MCP-1 a CCL7 chemokiny. STING deficitní monocyty jsou vnitřně defektní při migraci do jater během Listeria monocytogenes infekce. Tímto způsobem chrání STING hostitele před Listeria monocytogenes infekce regulací monocyt migrace. Aktivaci STING pravděpodobně zprostředkuje cyklický-di-AMP vylučované intracelulárními bakteriemi.[17][18]
jiný
STING může být důležitou molekulou pro ochranu imunity proti infekčním organismům. Například zvířata, která nemohou vyjádřit STING, jsou náchylnější k infekci VSV, HSV-1 a Listeria monocytogenes, což naznačuje jeho potenciální korelaci s lidskými infekčními chorobami.[19]
Role v imunitě hostitele
Ačkoli IFN typu I je naprosto zásadní pro odolnost vůči virům, roste literatura o negativní roli viru interferon typu I. v imunitě hostitele zprostředkované STING. Motiv DNA kmenové smyčky bohaté na AT v Plasmodium falciparum a Plasmodium berghei genom a extracelulární DNA z Mycobacterium tuberculosis byly aktivovány interferon typu I. přes STING.[20][21] Perforace fagosomové membrány zprostředkovaná ESX1 sekreční systém umožňuje extracelulární mykobakteriální DNA přístup k hostitelským senzorům cytosolické DNA, čímž indukuje produkci interferon typu I. v makrofágech. Vysoký interferon typu I. podpis vede k M. tuberculosis patogeneze a prodloužená infekce.[21] Zprostředkováno STING-TBK1-IRF interferon typu I. odpověď je ústřední pro patogenezi experimentální mozkové malárie u laboratorních zvířat infikovaných Plasmodium berghei. Laboratorní myši s nedostatkem interferon typu I. reakce jsou rezistentní na experimentální mozkovou malárii.[20]
Signální mechanismy STING
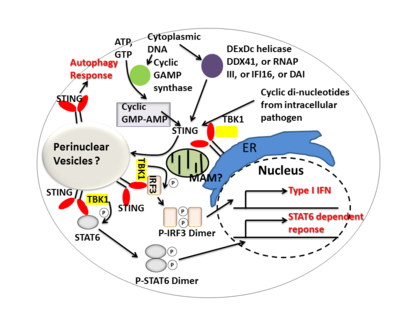
STING zprostředkovává interferon typu I. imunitní odpověď fungováním jak přímého senzoru DNA, tak a signální adaptér protein. Po aktivaci stimuluje STING TBK1 aktivita fosforylovat IRF3 nebo STAT6. Fosforylované IRF3 a STAT6 dimerizují a poté vstupují do jádra, aby stimulovaly expresi genů zapojených do imunitní odpovědi hostitele, jako je například IFNB, CCL2, CCL20, atd.[7][22]
Několik zpráv naznačuje, že STING je spojen s aktivací selektivní autofagie.[12] Mycobacterium tuberculosis Bylo prokázáno, že produkuje ligandy cytosolické DNA, které aktivují STING, což vede k ubikvitinace bakterií a následný nábor autofagie příbuzné proteiny, které jsou všechny potřebné pro „selektivní“ autofagické cílení a vrozenou obranu proti M. tuberculosis.[23]
Stručně řečeno, STING koordinuje více imunitních odpovědí na infekci, včetně indukce interferonů a reakce závislé na STAT6 a selektivní autofagické odpovědi.[7]
Jako senzor cytosolické DNA
Cyklické dinukleotidy - signální molekuly druhého posla produkované různými bakteriálními druhy byly detekovány v cytosolu savčích buněk během intracelulární patogenní infekce; to vede k aktivaci TBK1 -IRF3 a navazující výroba interferon typu I..[7][24]Ukázalo se, že STING se váže přímo na cyklický di-GMP a toto uznání vede k výrobě cytokiny, jako interferon typu I., které jsou nezbytné pro úspěšnou eliminaci patogenů.[25]
Jako signalizační adaptér
DDX41, člen rodiny helikáz DEXDc, v myeloidních dendritických buňkách rozpoznává intracelulární DNA a zprostředkovává vrozenou imunitní odpověď prostřednictvím přímé asociace s STING.[26] Další senzory DNA DAI, RNA polymeráza III, IFI16 Bylo také prokázáno, že aktivují STING přímými nebo nepřímými interakcemi.[14]
Cyklická GMP-AMP syntáza (cGAS), který patří do rodiny nukleotidyltransferáz, je schopen rozpoznat obsah cytosolické DNA a vyvolat STING-dependentní interferonovou reakci produkcí sekundárního posla cyklický guanosinmonofosfát – adenosinmonofosfát (cyklický GMP-AMP nebo cGAMP). Po cyklický GMP-AMP vázaný STING je aktivován, zvyšuje se TBK1 aktivita fosforylovat IRF3 a STAT6 pro navazující interferon typu I. Odezva.[27][28]
Bylo navrženo, že intracelulární vápník hraje důležitou roli v reakci dráhy STING.[29]
Reference
- ^ A b C ENSG00000288243 GRCh38: Vydání souboru 89: ENSG00000184584, ENSG00000288243 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000024349 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ "STING1 stimulátor interferonové odezvy cGAMP interakce 1 [Homo sapiens (člověk)]" ".
- ^ Nakhaei P, Hiscott J, Lin R (červen 2010). „ZAVÍRÁNÍ antivirové cesty“. Journal of Molecular Cell Biology. 2 (3): 110–2. doi:10.1093 / jmcb / mjp048. PMID 20022884.
- ^ A b C d E F Burdette DL, Vance RE (leden 2013). "STING a vrozená imunitní odpověď na nukleové kyseliny v cytosolu". Přírodní imunologie. 14 (1): 19–26. doi:10.1038 / ni.2491. PMID 23238760. S2CID 7968532.
- ^ Shu C, Yi G, Watts T, Kao CC, Li P (červenec 2012). „Struktura STING vázaného na cyklický di-GMP odhaluje mechanismus rozpoznávání cyklických dinukleotidů imunitním systémem“. Přírodní strukturní a molekulární biologie. 19 (7): 722–4. doi:10.1038 / nsmb.2331. PMC 3392545. PMID 22728658.
- ^ "EST expresní profil TMEM173". biogps org. biogps.org.
- ^ „NCBI TMEM173 expression GEOprofile“. NCBI. www.ncbi.nlm.nih.gov/geoprofiles.
- ^ A b Ishikawa H, Barber GN (říjen 2008). „STING je adaptér endoplazmatického retikula, který usnadňuje vrozenou imunitní signalizaci“. Příroda. 455 (7213): 674–8. doi:10.1038 / nature07317. PMC 2804933. PMID 18724357.
- ^ A b Saitoh T, Fujita N, Hayashi T, Takahara K, Satoh T, Lee H, Matsunaga K, Kageyama S, Omori H, Noda T, Yamamoto N, Kawai T, Ishii K, Takeuchi O, Yoshimori T, Akira S (prosinec 2009 ). „Atg9a řídí dsDNA řízenou dynamickou translokaci STING a vrozenou imunitní odpověď“. Sborník Národní akademie věd Spojených států amerických. 106 (49): 20842–6. doi:10.1073 / pnas.0911267106. PMC 2791563. PMID 19926846.
- ^ A b Barber GN (únor 2011). „Vrozené dráhy snímání imunitní DNA: STING, AIMII a regulace produkce interferonu a zánětlivých reakcí“. Aktuální názor na imunologii. 23 (1): 10–20. doi:10.1016 / j.coi.2010.12.015. PMC 3881186. PMID 21239155.
- ^ A b Keating SE, Baran M, Bowie AG (prosinec 2011). „Senzory cytosolické DNA regulující indukci interferonu typu I“ (PDF). Trendy v imunologii. 32 (12): 574–81. doi:10.1016 / j.it.2011.08.004. hdl:2262/68041. PMID 21940216.
- ^ Ma Z, Damania B (únor 2016). „Cesta obrany cGAS-STING a její potírání viry“. Mobilní hostitel a mikrob. 19 (2): 150–8. doi:10.1016 / j.chom.2016.01.010. PMC 4755325. PMID 26867174.
- ^ Xie J, Li Y, Shen X, Got G, Zhu Y, Cui J, Wang L, Shi Z, Zhou P (březen 2018). "Tlumená aktivace interferonu závislá na STING u netopýrů". Mobilní hostitel a mikrob. 23 (3): 297–301.e4. doi:10.1016 / j.chom.2018.01.006. PMC 7104992. PMID 29478775.
- ^ A b Jin L, Getahun A, Knowles HM, Mogan J, Akerlund LJ, Packard TA, Perraud AL, Cambier JC (březen 2013). „STING / MPYS zprostředkovává obranu hostitele proti infekci Listeria monocytogenes regulací migrace monocytů Ly6C (hi)“. Journal of Immunology. 190 (6): 2835–43. doi:10,4049 / jimmunol.1201788. PMC 3593745. PMID 23378430.
- ^ Woodward JJ, Iavarone AT, Portnoy DA (červen 2010). „c-di-AMP vylučovaný intracelulárně Listeria monocytogenes aktivuje interferonovou reakci hostitele typu I“. Věda. 328 (5986): 1703–5. doi:10.1126 / science.1189801. PMC 3156580. PMID 20508090.
- ^ Ishikawa H, Ma Z, Barber GN (říjen 2009). „STING reguluje intracelulární DNA zprostředkovanou vrozenou imunitu závislou na interferonu typu I“. Příroda. 461 (7265): 788–92. doi:10.1038 / nature08476. PMC 4664154. PMID 19776740.
- ^ A b Sharma S, DeOliveira RB, Kalantari P, Parroche P, Goutagny N, Jiang Z, Chan J, Bartholomeu DC, Lauw F, Hall JP, Barber GN, Gazzinelli RT, Fitzgerald KA, Golenbock DT (srpen 2011). „Vrozené imunitní rozpoznání motivu kmenové smyčky bohaté na AT v genomu Plasmodium falciparum“. Imunita. 35 (2): 194–207. doi:10.1016 / j.immuni.2011.05.016. PMC 3162998. PMID 21820332.
- ^ A b Manzanillo PS, Shiloh MU, Portnoy DA, Cox JS (květen 2012). „Mycobacterium tuberculosis aktivuje DNA závislou cytosolickou sledovací cestu v makrofázích“. Mobilní hostitel a mikrob. 11 (5): 469–80. doi:10.1016 / j.chom.2012.03.007. PMC 3662372. PMID 22607800.
- ^ Chen H, Sun H, You F, Sun W, Zhou X, Chen L, Yang J, Wang Y, Tang H, Guan Y, Xia W, Gu J, Ishikawa H, Gutman D, Barber G, Qin Z, Jiang Z (Říjen 2011). „Aktivace STAT6 metodou STING je pro antivirovou vrozenou imunitu zásadní“. Buňka. 147 (2): 436–46. doi:10.1016 / j.cell.2011.09.022. PMID 22000020.
- ^ Watson RO, Manzanillo PS, Cox JS (srpen 2012). „Extracelulární M. tuberculosis DNA se zaměřuje na bakterie pro autofagii aktivací hostitelské dráhy pro snímání DNA“. Buňka. 150 (4): 803–15. doi:10.1016 / j.cell.2012.06.040. PMC 3708656. PMID 22901810.
- ^ McWhirter SM, Barbalat R, Monroe KM, Fontana MF, Hyodo M, Joncker NT, Ishii KJ, Akira S, Colonna M, Chen ZJ, Fitzgerald KA, Hayakawa Y, Vance RE (srpen 2009). „Reakce hostitele typu I na interferon je vyvolána cytosolickým snímáním bakteriálního druhého posla cyklického di-GMP“. The Journal of Experimental Medicine. 206 (9): 1899–911. doi:10.1084 / jem.20082874. PMC 2737161. PMID 19652017.
- ^ Burdette DL, Monroe KM, Sotelo-Troha K, Iwig JS, Eckert B, Hyodo M, Hayakawa Y, Vance RE (říjen 2011). „STING je přímý vrozený imunitní senzor cyklického di-GMP“. Příroda. 478 (7370): 515–8. doi:10.1038 / příroda10429. PMC 3203314. PMID 21947006.
- ^ Zhang Z, Yuan B, Bao M, Lu N, Kim T, Liu YJ (říjen 2011). „Helikáza DDX41 snímá intracelulární DNA zprostředkovanou adaptérem STING v dendritických buňkách“. Přírodní imunologie. 12 (10): 959–65. doi:10.1038 / ni.2091. PMC 3671854. PMID 21892174.
- ^ Wu J, Sun L, Chen X, Du F, Shi H, Chen C, Chen ZJ (únor 2013). „Cyclic GMP-AMP je endogenní druhý posel vrozené imunitní signalizace cytosolickou DNA“. Věda. 339 (6121): 826–30. doi:10.1126 / science.1229963. PMC 3855410. PMID 23258412.
- ^ Sun L, Wu J, Du F, Chen X, Chen ZJ (únor 2013). „Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway“. Věda. 339 (6121): 786–91. doi:10.1126 / science.1232458. PMC 3863629. PMID 23258413.
- ^ Kim S, Koch P, Li L, Peshkin L, Mitchison TJ (4. června 2017). "Důkazy o roli vápníku v signalizaci STING". bioRxiv 10.1101/145854.
Další čtení
- Wang Y, Tong X, Omoregie ES, Liu W, Meng S, Ye X (říjen 2012). „Tetraspanin 6 (TSPAN6) negativně reguluje imunitní signalizaci zprostředkovanou kyselinou retinovou pomocí genu I podobného receptoru způsobem závislým na ubikvitinaci“. The Journal of Biological Chemistry. 287 (41): 34626–34. doi:10.1074 / jbc.M112.390401. PMC 3464568. PMID 22908223.
- Yin Q, Tian Y, Kabaleeswaran V, Jiang X, Tu D, Eck MJ, Chen ZJ, Wu H (červen 2012). „Cyklické snímání di-GMP prostřednictvím vrozeného imunitního signálního proteinu STING“. Molekulární buňka. 46 (6): 735–45. doi:10.1016 / j.molcel.2012.05.029. PMC 3697849. PMID 22705373.
- Aguirre S, Maestre AM, Pagni S, Patel JR, Savage T, Gutman D, Maringer K, Bernal-Rubio D, Shabman RS, Simon V, Rodriguez-Madoz JR, Mulder LC, Barber GN, Fernandez-Sesma A (2012) . "DENV inhibuje produkci IFN typu I v infikovaných buňkách štěpením lidského STINGu". PLOS patogeny. 8 (10): e1002934. doi:10.1371 / journal.ppat.1002934. PMC 3464218. PMID 23055924.
- Li Y, Li C, Xue P, Zhong B, Mao AP, Ran Y, Chen H, Wang YY, Yang F, Shu HB (květen 2009). „ISG56 je regulátor negativní zpětné vazby virem spouštěné signalizace a buněčné antivirové reakce“. Sborník Národní akademie věd Spojených států amerických. 106 (19): 7945–50. doi:10.1073 / pnas.0900818106. PMC 2683125. PMID 19416887.
- Conlon J, Burdette DL, Sharma S, Bhat N, Thompson M, Jiang Z, Rathinam VA, Monks B, Jin T, Xiao TS, Vogel SN, Vance RE, Fitzgerald KA (květen 2013). „Myš, ale nikoli lidská STING, se váže a signalizuje v reakci na látku narušující cévy 5,6-dimethylxanthenon-4-octovou kyselinu“. Journal of Immunology. 190 (10): 5216–25. doi:10,4049 / jimmunol. 1300097. PMC 3647383. PMID 23585680.
- Abe T, Harashima A, Xia T, Konno H, Konno K, Morales A, Ahn J, Gutman D, Barber GN (duben 2013). „STING rozpoznávání cytoplazmatické DNA podněcuje buněčnou obranu“. Molekulární buňka. 50 (1): 5–15. doi:10.1016 / j.molcel.2013.01.039. PMC 3881179. PMID 23478444.
- Nazmi A, Mukhopadhyay R, Dutta K, Basu A (2012). „STING zprostředkovává vrozenou imunitu neuronů po infekci virem japonské encefalitidy“. Vědecké zprávy. 2: 347. doi:10.1038 / srep00347. PMC 3317237. PMID 22470840.
- Zhang J, Hu MM, Wang YY, Shu HB (srpen 2012). „Protein TRIM32 moduluje indukci interferonu typu I a buněčnou antivirovou reakci zaměřením proteinu MITA / STING na ubikvitinaci spojenou s K63“. The Journal of Biological Chemistry. 287 (34): 28646–55. doi:10.1074 / jbc.M112.362608. PMC 3436586. PMID 22745133.
- Ishikawa H, Barber GN (říjen 2008). „STING je adaptér endoplazmatického retikula, který usnadňuje vrozenou imunitní signalizaci“. Příroda. 455 (7213): 674–8. doi:10.1038 / nature07317. PMC 2804933. PMID 18724357.