Glukosamin-fosfát N-acetyltransferáza - Glucosamine-phosphate N-acetyltransferase
| glukosamin 6-fosfát N-acetyltransferáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||
| EC číslo | 2.3.1.4 | ||||||||
| Číslo CAS | 9031-91-8 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
v enzymologie, glukosamin-fosfát N-acetyltransferáza (GNA) (ES 2.3.1.4 ) je enzym že katalyzuje přenos acetylové skupiny z acetyl-CoA na primární amin v glukosamid-6-fosfát, generování bezplatného CoA a N-acetyl-D-glukosamin-6-fosfát.[1]
Tento enzym patří do rodiny transferázy, skupina enzymů, která přenáší velmi specifickou funkční skupinu, v tomto případě acetyl, z dárce na receptor. Konkrétně lze tento enzym charakterizovat jako součást acyltransferázy rodiny, protože jde o převod generála acylová skupina s methylem jako substituentem.
Nomenklatura
The systematické jméno z této třídy enzymů je acetyl-CoA: D-glukosamin-6-fosfát N-acetyltransferáza. Mezi další běžně používané názvy patří fosfoglukosamin-transacetyláza, fosfoglukosamin-acetyláza, glukosamin-6-fosfát-acetyláza, D-glukosamin-6-P-N-acetyltransferáza, aminodeoxyglukóza-fosfát-acetyltransferáza, glukosamin-6-fosfát-acetyláza, glukosamin-6-fosfát-N-acetyláza -6-fosfát syntáza, fosfoglukosamin N-acetyláza, glukosamin-fosfát N-acetyltransferáza a glukosamin-6-fosfát N-acetyltransferáza.
Funkce
Tento enzym je součástí dráhy biosyntézy hexosaminu[2] (HBP), který je jedním z glukóza procesy zpracování v obecném metabolismu. Tato cesta sdílí počáteční dva kroky glykolýza a odchyluje se pouze malá část tok glukózy z této tradičnější glykolytické dráhy. Proto je výhodné, když existuje negativní regulace zpětné vazby na glykolýzu, jako v případě velkého množství volného mastné kyseliny. Konečným produktem této cesty je UDP-N-acetylglukosamin, který se podílí na modifikaci komplexních molekul, jako je glykolipidy, proteoglykany[3] a glykoproteiny. Tento konečný produkt funguje jako nosič N-acetylglukosamin, což je monomerní jednotka chitinu,[4] strukturní polymer, který tvoří skořápky korýši a hmyz, stejně jako buněčná stěna z houby. Dále N-acetylglukosamin je také jednotkou peptidoglykan polymer, který tvoří bakterie buněčná stěna[5] spolu s N-acetylmuramovým disacharidem.

Přesněji řečeno, enzym GNA katalyzuje čtvrtý krok dráhy HBP v eukaryoty, podpora přenosu uhlíku z Acetyl-CoA na druhý substrát, D-glukosamin-6-fosfát který nakonec přinese UDP-N-acetylglukosamin. Jedná se o malý, ale důležitý chemický krok, který je zásadní pro vlastnosti vedlejších produktů této metabolické dráhy. The acetylace se provádí až do samého konečného produktu hexaminové dráhy a je velmi charakteristický pro polymery vytvořen s N-acetylglukosamin. Například představuje jeden z hlavních rozdílů v molekulární struktuře chitin a celulóza,[7] a vysvětluje mnoho fyzikálních a chemických vlastností těchto polymerů. V případě chitin například výpočetní studie zjistily, že acylace přispívá k tvorbě Vodíkové vazby které stabilizují krystalickou strukturu tohoto polymeru a poskytují větší odolnost proti lomu.[8]
V prokaryotickém metabolismu však biosyntetická cesta hexosaminu sleduje jiný reakční krok, ve kterém jiný enzym působí na stejné charakteristické substráty[6] (Obrázek 1). v prokaryoty, k přenosu fosfátů ze 6-uhlíku na 1-uhlík dochází před acylací, takže substrát reakce přidávající uhlík je Glukosamin-1-fosfát spíše než D-glukosamin-6-fosfát. Tentokrát je enzymem zodpovědným za acetylaci bifunkční protein glmU (N-acetylglukosamin-1-fosfát uridyltransferáza),[9] který také katalyzuje přidání UDP k fosfátové skupině na N-acetyl-D-glukosamin-1-fosfátu.
U lidí je glukosamin-fosfát N-acetyltransferáza a dimer se dvěma identickými podjednotky,[10] a je kódován v genu GNPNAT[11] (Symbol HGNC). Přesněji řečeno, enzym je silně exprimován v játra, žaludek a tkáně gastrointestinálního traktu a v buňce se nachází v endozomy a v Golgiho aparát (manuální anotací).[11]
Mechanismus
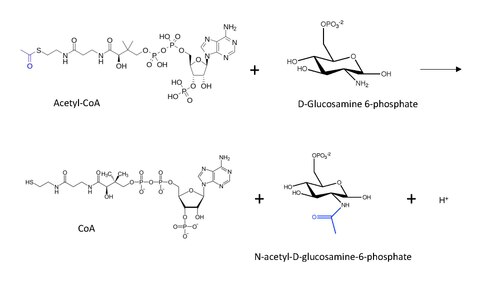
Molekulární struktura reakce katalyzované GNA je uvedena níže, s přenesenou acetylovou skupinou modře.
Obecný reakční mechanismus předpokládaný pro acetylaci N-konce proteinu (inspirovaný mechanismem acetylace lysinu) s Acetyl-CoA zahrnuje nukleofilní útok aminoskupiny (v tomto případě z D-glukosamin-6-fosfátu) na konci karbonyl v přenosu uhlíku, což vede k tvorbě uhlíkového tetrahedrálního meziproduktu.[13] Reakce pokračuje obnovou karbonyl odstraněním CoA jako odstupující skupina, takže nyní je acetylová skupina připojena k amino skupina v druhém substrátu.
Konkrétně pro tento katalyzátor N-acetyltransferázy, studie s S. cerevisiae Enzym GNA ukázal, že některé specifické aminokyseliny přispívají k vazbě substrátu, zvýšené nukleofilita aminoskupiny a nakonec katalýza, která podporuje výše popsaný postulovaný mechanismus.[14] Glu98, Asp99 a Ile100 polarizují karbonyl vazba v Acetyl-CoA, zvyšující uhlík elektrofilita stejně jako stabilizace uhlíkového čtyřbokého meziproduktu. Tyr143 je zodpovědný za stabilizaci thiolát anion, upřednostňující S-CoA jako odstupující skupinu z čtyřbokého uhlíku. Nakonec Asp134 vylepšuje nukleofilita z amino skupina v D-glukosamid-6-fosfátu darováním elektronové hustoty do dusík atom. V jiném organismu C. albicans, bylo zjištěno, že podobná sada aminokyselin je nezbytná pro katalytickou aktivitu,[15] respektive systém Glu88-Asp-89-Ile90, Asp125 a Tyr133.
Struktura

Ke konci roku 2019, 13 struktur byly pro tuto třídu enzymů u různých druhů vyřešeny pomocí PDB přístupové kódy 1I12 (Saccharomyces cerevisiae), 1I1D (Saccharomyces cerevisiae), 1I21 (Saccharomyces cerevisiae), 2HUZ (Homo sapiens), 2O28 (Homo sapiens), 4AG7 (Caenorhabditis elegans), mezi ostatními.
Obrázek 3 ukazuje navrhovanou krystalovou strukturu GNA v lidé,[17] s každou katalytickou podjednotkou v jiné barvě. The Acetyl-CoA navázaný na enzym je zobrazen světle růžově a produkt stále navázaný na katalytické místo je zobrazen fialově. Převedeno acetyl skupina v N-acetyl-D-glukosamin-6-fosfát produkt ve fialové barvě je zobrazen žlutě. Tato navrhovaná 3d struktura protein ukazuje, že specifické části substrátů zapojených do této reakce - terminální konec lineární části Acetyl-CoA a dusík skupina připojená k glukosaminovému kruhu - jsou ve velké blízkosti.
Reference
- ^ Kato N, Mueller CR, Wessely V, Lan Q, Christensen BM (červen 2005). „Mosquito glukosamin-6-fosfát N-acetyltransferáza: cDNA, genová struktura a kinetika enzymů“. Hmyzí biochemie a molekulární biologie. 35 (6): 637–46. doi:10.1016 / j.ibmb.2005.02.005. PMID 15857769.
- ^ Schwarzer M, Doenst T (2016). Vědecký průvodce srdečním metabolizmem. Akademický tisk. 39–55. ISBN 9780128023945.
- ^ Kim YH, Nakayama T, Nayak J (leden 2018). „Glykolýza a biosyntetická cesta hexosaminu jako nové cíle pro zánět horních a dolních dýchacích cest“. Výzkum alergie, astmatu a imunologie. 10 (1): 6–11. doi:10.4168 / aair.2018.10.1.6. PMC 5705485. PMID 29178672.
- ^ Cohen E (říjen 2001). "Syntéza a inhibice chitinu: revize". Věda o ochraně proti škůdcům. 57 (10): 946–50. doi:10.1002 / ps.363. PMID 11695188.
- ^ Meroueh SO, Bencze KZ, Hesek D, Lee M, Fisher JF, Stemmler TL, Mobashery S (březen 2006). "Trojrozměrná struktura peptidoglykanu bakteriální buněčné stěny". Sborník Národní akademie věd Spojených států amerických. 103 (12): 4404–9. Bibcode:2006PNAS..103,4404M. doi:10.1073 / pnas.0510182103. PMC 1450184. PMID 16537437.
- ^ A b Riegler H, Herter T, Grishkovskaya I, Lude A, Ryngajllo M, Bolger ME, Essigmann B, Usadel B (duben 2012). "Krystalová struktura a funkční charakterizace glukosamin-6-fosfát N-acetyltransferázy z Arabidopsis thaliana". The Biochemical Journal. 443 (2): 427–37. doi:10.1042 / BJ20112071. PMID 22329777.
- ^ „Obr. 1 Chemická struktura celulózy a chitinu“. ResearchGate. Citováno 2019-03-15.
- ^ Cui J, Yu Z, Lau D (leden 2016). „Vliv acetylové skupiny na mechanické vlastnosti nanokrystalů chitin / chitosan: studie molekulární dynamiky“. International Journal of Molecular Sciences. 17 (1): 61. doi:10,3390 / ijms17010061. PMC 4730306. PMID 26742033.
- ^ Vithani N, Bais V, Prakash B (červen 2014). „GlmU (N-acetylglukosamin-1-fosfát uridyltransferáza) vázaný na tři ionty hořčíku a ATP v aktivním místě“. Acta Crystallographica oddíl F. 70 (Pt 6): 703–8. doi:10.1107 / S2053230X14008279. PMC 4051520. PMID 24915076.
- ^ Wang J, Liu X, Liang YH, Li LF, Su XD (září 2008). „Vazba na substrát akceptoru odhalena krystalovou strukturou lidské glukosamin-6-fosfát N-acetyltransferázy 1“. FEBS Dopisy. 582 (20): 2973–8. doi:10.1016 / j.febslet.2008.07.040. PMID 18675810. S2CID 5131540.
- ^ A b „GNPNAT1 - Glukosamin 6-fosfát N-acetyltransferáza - Homo sapiens (člověk) - GNPNAT1 gen a protein“. www.uniprot.org. Citováno 2019-03-15.
- ^ "FlyBase - aktivita glukosamin 6-fosfát N-acetyltransferázy".
- ^ Lim S, Smith KR, Lim ST, Tian R, Lu J, Tan M (2016-04-14). „Regulace mitochondriálních funkcí fosforylací a defosforylací proteinů“. Cell & Bioscience. 6: 25. doi:10.1186 / s13578-016-0089-3. PMC 4832502. PMID 27087918.
- ^ Mio T, Yamada-Okabe T, Arisawa M, Yamada-Okabe H (leden 1999). „Saccharomyces cerevisiae GNA1, základní gen kódující novou acetyltransferázu zapojenou do syntézy UDP-N-acetylglukosaminu“. The Journal of Biological Chemistry. 274 (1): 424–9. doi:10.1074 / jbc.274.1.424. PMID 9867860.
- ^ Milewski S, Gabriel I, Olchowy J (leden 2006). "Enzymy biosyntézy UDP-GlcNAc v kvasinkách". Droždí. 23 (1): 1–14. doi:10,1002 / ano 1337. PMID 16408321. S2CID 39940329.
- ^ PDB: 2O28; Plotnikov AN, Bochkarev A, Edwards AM, Arrowsmith CH, Sundstrom M, Weigelt J a kol. "Krystalová struktura glukosamin-fosfát N-acetyltransferázy 1". Bude zveřejněno. doi:10,2210 / pdb2o28 / pdb.
- ^ Peneff C, Mengin-Lecreulx D, Bourne Y (květen 2001). „Krystalové struktury Apo a komplexu Saccharomyces cerevisiae GNA1 osvětlují katalytický mechanismus amino-cukr N-acetyltransferázy“. The Journal of Biological Chemistry. 276 (19): 16328–34. doi:10,1074 / jbc.M009988200. PMID 11278591.