Enantiornithes - Enantiornithes
| Enantiornithes | |
|---|---|
 | |
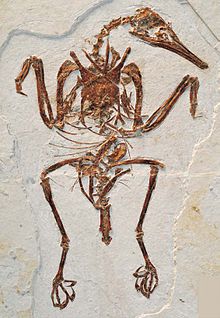
| Fosilní exemplář a bohaiornithid (Zhouornis hani ) | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Clade: | Dinosaurie |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Ornithothoraces |
| Clade: | †Enantiornithes chodec, 1981 |
| Podskupiny | |
a zobrazit text | |
Enantiornithes je skupina vyhynulých avialans („ptáci“ v širším slova smyslu), nejhojnější a nejrozmanitější skupina známá z Mezozoická éra.[3][4][5] Téměř všechny si ponechaly zuby a drápané prsty na každém křídle, ale jinak vypadaly navenek podobně jako moderní ptáci. Bylo pojmenováno více než 80 druhů enantiornitheans, ale některá jména představují pouze jednotlivé kosti, takže je pravděpodobné, že ne všechny jsou platné. Enantiornitheans vyhynuli na Křída – paleogenní hranice, spolu s hesperornithids a všechny ostatní jiné než ptačí dinosauři.
Objev a pojmenování
První objevení enantiornitheans byli nesprávně odkazoval se na moderní skupiny ptáků. Například první známý enantiornithean, Gobipteryx minuta, byl původně považován za paleognath související s pštrosi a tinamou.[6] Enantiornitheans byl nejprve rozpoznán jako zřetelná linie nebo „podtřída“ ptáků Cyril A. Walker v roce 1981. Walker učinil tento objev na základě několika částečných pozůstatků z pozdní doby Křídový doba toho, co je teď Argentina, kterou přiřadil novému rodu, Enantiornis, pojmenování celé skupiny. Od 90. let 20. století bylo objeveno mnohem více úplných enantiornitů a bylo zjištěno, že několik dříve popsaných „ptáků“ (např. Iberomesornis, Cathayornis, a Sinornis ) byli také enantiornitheans.
Jméno "Enantiornithes" znamená "protější ptáci" od Starořečtina enantios (ἐνάντιος) „naproti“ + ornithy (ὄρνιθες) „ptáci“. Název vytvořil Cyril Alexander Walker ve svém orientačním článku, který skupinu založil.[7] Walker ve svém příspěvku vysvětlil, co měl na mysli „protikladem“:
Snad nejzásadnějším a nejcharakterističtějším rozdílem mezi Enantiornithy a všemi ostatními ptáky je povaha artikulace mezi lopatka [...] a coracoid, kde je „normální“ stav zcela obrácen.[7]
To se týká anatomického znaku - artikulace rameno kosti - který má konkávně konvexní kloubový kloub, který je opakem moderních ptáků. Konkrétně v enantiornitheans, aspekt, kde lopatka (lopatka) splňuje coracoid (primární kost ramenního pletence u jiných obratlovců než savců) je a konvexní knoflík a odpovídající bod na lopatce je konkávní a ve tvaru misky. U moderních ptáků je způsob artikulace kloubu obrácen.[8]
Walker neměl jasno ve svých důvodech pro uvedení tohoto jména v části etymologie své práce a tato nejednoznačnost vedla k určitému zmatku mezi pozdějšími badateli. Například, Alan Feduccia uvedeno v roce 1996:
Ptáci jsou tak pojmenováni, protože mezi mnoha charakteristickými rysy existuje jedinečná formace trioseálního kanálu a metatarzály jsou spojeny proximálně až distálně, což je opak u moderních ptáků[9]
Názor Feduccie o tarsometatarsus (kombinovaná horní noha a kotníková kost) jsou správné, ale Walker tuto úvahu ve svém původním článku nepoužil. Walker nikdy nepopisoval fúzi tarsometatarsusu jako opačnou, ale spíše jako „Pouze částečnou“. Rovněž není jisté, že enantiornitheans měli trioseální kanály, protože žádná vlastnost nezachovává tuto vlastnost.[3]
Jako skupina se Enantiornithy často označují jako „enantiornithiny“. Několik vědců však poznamenalo, že to není správné, protože podle standardních pravidel pro vytváření názvů skupin zvířat to znamená odkaz pouze na podčeleď Enantiornithinae. V návaznosti na konvence pojmenování používané pro moderní ptáky i vyhynulé skupiny bylo zdůrazněno, že správný termín je „enantiornithean“.[10]
Původ a rozsah
Enantiornitheans byly nalezeny na všech kontinentech kromě Antarktida. Fosílie, které lze připsat této skupině, jsou výlučně Křídový ve věku a předpokládá se, že Enantiornitheans vyhynuli ve stejnou dobu jako jejich non-avialan dinosaurus příbuzní. Nejdříve známé enantiornitheans jsou z Raná křída z Španělsko (např. Noguerornis ) a Čína (např. Protopteryx ) a nejnovější z Pozdní křída Severní a Jižní Ameriky (např. Avisaurus a Enantiornis ). Široký výskyt této skupiny naznačuje, že přinejmenším někteří enantiornithové byli schopni překročit oceány vlastní silou; jsou první známou avialanskou linií s globální distribucí.
Popis

Mnoho enantiornithean fosílie jsou velmi fragmentární, a některé druhy jsou známy pouze z kusu jedné kosti. Jsou známy téměř všechny vzorky, které jsou úplné, v plné artikulaci a se zachováním měkkých tkání Las Hoyas v Cuenca, Španělsko a Jehol skupina v Liaoning (Čína ). Enantiornithské fosílie byly nalezeny ve vnitrozemských i mořských sedimentech, což naznačuje, že se jednalo o ekologicky rozmanitou skupinu. Zdá se, že Enantiornitheans zahrnovali brodivé ptáky, plavce, granivory, hmyzožravce, rybáře a dravce. Drtivá většina druhů enantiornithean byla malá, mezi velikostí a Vrabec a a špaček,[11] zatímco největší členové tohoto kladu jsou Pengornis houi,[12] Xiangornis shenmi[13] a Zhouornis hani.[11] Mohlo existovat alespoň několik mnohem větších druhů, včetně potenciálně jeřábovitých druhů známých pouze ze stop v Eumeralla formace (a případně také zastoupena v EU) Wonthaggi formace jediným furcula ), který by mohl patřit k enantiornithean.[14] Jeden taxon, Mirarce, je popsán jako podobný velikosti jako moderní krůty,[15] zatímco předchozí „největší enantiornithové“ jsou popsáni jako kos velikosti.[12]
Mimořádné enantiornithean pozůstatky byly také zachovány v Barmský jantar vklady datované před 99 miliony let. Tyto pozůstatky patří k nejzachovalejším ze všech mezozoických dinosaurů. První objevené jantarově obalené enantiornitické pozůstatky byly dvě křídla (viz níže) popsaná v roce 2016.[16] V roce 2017 bylo popsáno téměř celé tělo mládě enantiornitheana[17] a další mládě bylo popsáno v roce 2018.[18] V roce 2019 byla popsána převážně úplná noha spolu s křídlem.[19] V roce 2020 bylo popsáno křídlo velkého taxonu.[20]
Lebka

Vzhledem k jejich široké škále stanovišť a stravovacích návyků se lebky enantiornitheans mezi druhy značně lišily. Enantiornithské lebky kombinovaly jedinečnou sadu primitivních a pokročilých funkcí. Stejně jako u primitivnějších avialanů Archeopteryx, zachovali si několik samostatných lebečních kostí, malých premaxillae (kosti špičky čenichu) a většina druhů měla spíše zubaté čelisti než bezzubé zobáky. Pouze několik druhů, jako např Gobipteryx minuta, byli zcela bezzubí a měli zobáky. Měli také jednoduché kvadrátové kosti, kompletní lišta oddělující každý obíhat (oční otvor) od každého antorbitální fenestra a zubary (hlavní ozubené kosti dolní čelisti) bez vidlicových zadních špiček. A dlaždicová kost je zachována v neurčitém juvenilním vzorku, zatímco a postorbitální je zachována v Shenqiornis a Pengornis. U moderních ptáků jsou tyto kosti asimilovány do lebky. Někteří enantiornitheans možná měli své temporální fenestry (díry v boku hlavy) splynuly na oběžné dráhy jako u moderních ptáků, protože postorbitály buď nebyly přítomny, nebo nebyly dostatečně dlouhé na to, aby rozdělily otvory.[21] A čtyřhlavá kost, který je v moderních ptácích spojen s jugal, je zachován v Pterygornis.[22] Přítomnost těchto primitivních rysů lebky by znamenala, že enantiornithové jsou schopní jen omezeně lebeční kineze (schopnost pohybovat čelistí nezávisle na lebce).[23]
Křídlo

Jako velmi velká skupina ptáků vykazovali enantiornitheans vysokou rozmanitost různých tělesných plánů založených na rozdílech v ekologii a krmení, což se odráží ve stejné rozmanitosti forem křídel, mnoho paralelních adaptací na různé životní styly, které se vyskytují u moderních ptáků. Obecně byla křídla enantiornitheanů ve srovnání s primitivnějšími avialanky podobná Archeopteryx, a zobrazeny některé rysy související s letem podobné těm, které byly nalezeny v linii vedoucí k moderním ptákům, Ornithuromorpha. Zatímco většina enantiornitheans měla drápy na alespoň některých prstech, mnoho druhů mělo zkrácené ruce, vysoce mobilní ramenní kloub a proporcionální změny v křídlech kostí podobné moderním ptákům. Stejně jako moderní ptáci měli i enantiornitheans alulas, nebo „bastardská křídla“, malá dopředu směřující uspořádání peří na první číslici, která zaručovala vyšší manévrovatelnost ve vzduchu a napomáhala přesným přistáním.[24]
Několik křídel se zachovaným peřím bylo nalezeno zachováno v roce Barmský jantar. Jedná se o první kompletní Druhohor dinosaurus zůstává zachována tímto způsobem (jinak je známo několik izolovaných peří, která nejsou přiřazena žádnému druhu) a jedna z nejkrásněji známých fosilií dinosaurů.[25] Zachovaná křídla ukazují variace v peří pigmentu a dokazují, že enantiornitheans měli plně moderní peří, včetně ostnů, barbulů a háčků, a moderní uspořádání peří křídla včetně dlouhých letek, krátkých houští, velké aluly a podsady dolů.[16]
Jedna enantiornithská fosilie vykazuje na nohou podobné chuchvalce křídlového peří Archeopteryx. Peří na noze také připomíná čtyřkřídlého dinosaura Mikroraptor v enantiornithean se však liší od peří jsou kratší, neuspořádanější (nevytvářejí jasně křídlo) a sahají pouze po kotník, spíše než po chodidlo[26]
Ocas
Clarke et al. (2006) zkoumali všechny enantiornitheanské fosilie, které byly tehdy známy, a dospěli k závěru, že nikdo nezachoval ocasní peří, které tvořilo ventilátor vytvářející výtah, jako u moderních ptáků. Zjistili, že všichni letci mimo Euornithy (kladu, kterému říkali Ornithurae ) se zachovalými ocasními pery měla pouze krátké návleky nebo protáhlé spárované ocasní pera. Navrhli, aby byl vývoj pygostyle v enantiornitheans muselo být funkcí zkracování ocasu, ne vývoje moderní anatomie ocasního peří. Tito vědci navrhli, aby je musel ovládat fanoušek ocasního peří a souvisejícího svalstva, známého jako rektální žárovka, se vyvinuly vedle krátkého trojúhelníkového pygostyla, jako jsou ty u moderních ptáků, spíše než dlouhé pygostyly ve tvaru tyče nebo dýky u primitivnějších avialanů jako enantiornitheans. Místo vějíře měla většina enantiornitheans pár dlouhých specializovaných pinfeathers podobných těm vyhynulým Konfuciusornis a jisté rajky.[27]
Další objevy však ukázaly, že přinejmenším u primitivních enantiornitů byla anatomie ocasu složitější, než se dříve myslelo. Jeden enantiornithean, Shanweiniao, byl původně interpretován jako mající nejméně čtyři dlouhá ocasní pera, která se navzájem překrývala[28] a mohl by vytvořit povrch vytvářející vztlak podobný ocasním ventilátorům euornitheans,[29] ačkoli pozdější studie to naznačuje Shanweiniao bylo pravděpodobnější rachis -dominované ocasní peří podobné peřím přítomným v Paraprotopteryx.[30] Chiappeavis primitivní pengornithid enantiornithean, měl fanoušek ocasních peří podobných těm primitivnějších avialanů Sapeornis, což naznačuje, že se mohlo jednat o stav předků, přičemž funkce pinfeathers se pro účely zobrazení v časných avialanech několikrát vyvinuly.[30] Další enantiornithean, Feitianius, měl také komplikovaný fanoušek ocasních peří. Ještě důležitější je, že měkká tkáň zachovaná kolem ocasu byla interpretována jako pozůstatky rektální baňky, což naznačuje, že tento rys nebyl ve skutečnosti omezen na druhy s moderně vypadajícími pygostyly, ale mohl se vyvinout mnohem dříve, než se dříve myslelo, a byl přítomen v mnoha enantiornitheans.[31] Alespoň jeden rod enantiornithean, Cruralispennia, měl moderně vypadající pygostyl, ale postrádal ocasní ventilátor.[32]
Biologie
Strava
Vzhledem k velké rozmanitosti tvaru lebky mezi enantiornitheans muselo být ve skupině přítomno mnoho různých dietních specializací. Někteří, jako Shenqiornis, měl velké, robustní čelisti vhodné k jídlu bezobratlých s tvrdým krunýřem. v longipterygids, čenichy byly dlouhé a tenké se zuby omezenými na špičku čelistí, a byly to pravděpodobně bláto-probers (malé zuby) a rybáři (velké zuby). Krátké, tupé zuby Pengornis se pravděpodobně používaly ke krmení členovců s měkkým tělem.[21] Silně zahnuté drápy bohaiornithids naznačují, že se jednalo o predátory malých až středních obratlovců, ale jejich robustní zuby místo toho naznačují stravu tvrdě vylupovaných zvířat.[1]
Několik vzorků zachovává skutečný obsah žaludku. Bohužel žádný z nich nezachovává lebku, takže nelze provést přímou korelaci mezi jejich známou stravou a tvarem čenichu / zubu. Eoalulavis bylo zjištěno, že má pozůstatky exoskeletony z vodních korýši konzervované v trávicím traktu,[33] a Enantiophoenix zachovalé jantarové krvinky mezi zkamenělými kostmi, což naznačuje, že toto zvíře se živilo mízou, podobně jako moderní sapsuckers a další ptáci. Míza by zkameněla a stala by se jantarovou.[34] Avšak v poslední době se předpokládá, že míza se pohnula po smrti, a proto nepředstavovala skutečný obsah žaludku. V kombinaci s předpokládanou rybou pelety z Piscivorenantiornis Ukázalo se, že se jedná o rybí výkaly, což se ukázalo jako podivný žaludeční obsah některých druhů vaječníky a domnělé gastrolity Bohaiornis jsou náhodnými minerálními sraženinami, pouze Eoalulavis zobrazuje skutečný obsah žaludku.[35]
Studie o paravských zažívacích systémech naznačuje, že známým enantiornitheanům chyběla úroda a žaludek, nepoužívali gastrolity a nevyhazovali pelety. To je považováno za v rozporu s velkou rozmanitostí diet, které naznačují jejich různé tvary zubů a lebky,[36] ačkoli někteří moderní ptáci ztratili žaludek a spoléhají se pouze na silné žaludeční kyseliny.[37]
Predace
Sanz uvedl fosilii ze Španělska et al. v roce 2001 zahrnoval pozůstatky čtyř vylíhnutých enantiornithských koster tří různých druhů. Jsou v podstatě úplné, velmi úzce spojené a vykazují povrchovou důlku kostí, což naznačuje částečné trávení. Autoři dospěli k závěru, že tato asociace byla regurgitovaná peleta a z podrobností trávení a velikosti, že mláďata byla spolknuta celá ptakoještěr nebo malé theropod dinosaurus. Toto byl první důkaz Druhohor avialans byli zvířata kořisti, a to někteří druhohor pan-ptáci regurgitované pelety jako sovy dnes.[38]
Životní historie

Známé enantiornithské fosílie zahrnují vejce,[39][40] embrya,[41] a mláďata.[42] Enantiornithské embryo, stále stočené do vajíčka, bylo hlášeno z Yixian formace.[43] Juvenilní vzorky lze identifikovat kombinací faktorů: drsná struktura jejich kostních špiček, která označuje části, které byly v době smrti ještě vyrobeny z chrupavky, relativně malé hrudní kosti, velké lebky a oči a kosti, které k sobě ještě nespojily .[44] Některé vzorky mláďat dostaly oficiální jména, například „Liaoxiornis delicatus "; nicméně, Luis Chiappe a kolegové považovali praxi pojmenovávání nových druhů založených na mladistvých za škodlivou pro studium enantiornitheans, protože je téměř nemožné určit, ke kterému dospělému druhu daný mladistvý exemplář patří, takže jakýkoli druh s vylíhlým holotypem nomen dubium.[44]
Spolu s mláďaty mongolských exemplářů Gobipteryx[45] a Gobipipus,[46][47] tyto nálezy ukazují, že enantiornitheanská mláďata měla kostní osifikaci, dobře vyvinuté křídlové peří a velký mozek, který koreluje s precocial nebo superprecocial vzorce vývoje dnešních ptáků. Jinými slovy, enantiornitheans se pravděpodobně vylíhla z vajíčka již dobře vyvinutého a připraveného k běhu, krmení a případně dokonce létat ve věku pouhých několika dní.[44]
Byly provedeny analýzy histologie kostí za účelem stanovení rychlosti růstu těchto zvířat. Studie z roku 2006 Concornis kosti vykazovaly růstový vzor odlišný od moderních ptáků; ačkoli růst byl několik týdnů po vylíhnutí rychlý, pravděpodobně do opeření, tento malý druh nedosahoval dospělé velikosti po dlouhou dobu, pravděpodobně několik let.[48] Všechny další studie podporovaly názor, že růst do velikosti dospělého byl pomalý, jako je tomu u živých precocialních ptáků (na rozdíl od altriciální ptáci, o nichž je známo, že rychle dosahují velikosti dospělých).[33] Studie rychlosti růstu kostí u různých enantiornitheans ukázaly, že menší druhy měly tendenci růst rychleji než ty větší, což je opak vzoru pozorovaného u primitivnějších druhů, jako je Jeholornis a u jiných než avialanských dinosaurů.[49] Některé analýzy interpretovaly histologii kostí, což naznačuje, že enantiornitheans nemuseli mít úplně ptačí endotermie, místo toho mít meziprodukt rychlost metabolismu.[50]
Důkazy o koloniálním hnízdění byly nalezeny v enantiornitheans, v sedimentech z Pozdní křída (Maastrichtian ) z Rumunsko.[51] Důkazy z hnízdišť ukazují, že enantiornithové pohřbívali vajíčka jako moderní megapody, což je v souladu s jejich odvozenými superprecociálními adaptacemi.[52]
Studie z roku 2020 o enantiornitheanských mladistvých peřích dále zdůrazňuje ontologické podobnosti s moderními megapody, ale varuje před několika rozdíly, jako je stromová povaha většiny enantiornithů na rozdíl od pozemského životního stylu megapodů.[53]
Let
Protože mnoho enantiornitheans postrádalo složité ocasy a vlastnilo radikálně odlišnou anatomii křídla ve srovnání s moderními ptáky, byli předmětem několika studií testujících jejich letové schopnosti.
Tradičně byly považovány za podřadné letáky, protože anatomie ramenního pletence byla považována za primitivnější a neschopnou podporovat pozemní spouštěcí mechanismus,[54] stejně jako kvůli absenci rektifikace u mnoha druhů.[27][29][55]
Několik studií však ukázalo, že se jednalo o efektivní letáky, stejně jako moderní ptáci, kteří mají podobně složitý nervový systém a vazy křídlového peří. Navíc se zdá, že nedostatek komplexního ocasu nebyl příliš relevantní ptačí let jako celek - někteří vyhynulí ptáci mají rádi lithornidy také chyběly složité ocasní pera, ale byly to dobré letáky,[56] a zdá se, že byli schopni pozemního startu.[57]
Kvůli rozdílu v anatomii hrudního a ramenního pletence mnoho enantiornitheans používalo letový styl na rozdíl od jakéhokoli moderního druhu ptáka[je zapotřebí objasnění ], ačkoli byly přítomny také typičtější letové styly.[58]
Alespoň Elsornis se zdá být sekundárně nelétavý.[59]
Klasifikace
Někteří vědci zařazují do třídy enantiornitheans spolu se skutečnými ptáky Aves. Jiní používají přísnější skupina korun definice Aves (která zahrnuje pouze neornithes, anatomicky moderní ptáci) a zařadit enantiornitheans do inkluzivnější skupiny Avialae. Enantiornitheans byli pokročilejší než Archeopteryx, Konfuciusornis, a Sapeornis, ale v několika ohledech byli primitivnější než moderní ptáci, možná po střední evoluční cestě.
Konsenzus vědeckých analýz naznačuje, že Enantiornithes je jednou ze dvou hlavních skupin v rámci větší skupiny Ornithothoraces. Druhou ornithotoracinovou skupinou je Euornithy nebo Ornithuromorpha, který zahrnuje všechny živé ptáky jako podmnožinu. To znamená, že enantiornitheans byli úspěšnou větví avialanské evoluce, ale ta, která se diverzifikovala zcela odděleně od linie vedoucí k moderním ptákům.[3] Jedna studie však zjistila, že sdílená anatomie hrudní kosti byla získána nezávisle a je třeba tento vztah znovu přezkoumat.[60]
Enantiornithean klasifikace a taxonomie byla historicky komplikována řadou faktorů. V roce 2010 paleontologové Jingmai O'Connor a Gareth Dyke nastínili řadu kritik proti převládající praxi vědců, kteří nedokázali dostatečně podrobně popsat mnoho vzorků, aby je mohli ostatní důkladně vyhodnotit. Některé druhy byly popsány na základě vzorků, které jsou drženy v soukromých sbírkách, což znemožňuje další studium nebo přezkoumání předchozích nálezů. Vzhledem k tomu, že pro ostatní vědce je často nemožné osobně studovat každý vzorek vzhledem k celosvětové distribuci Enantiornithů, a vzhledem k mnoha neinformativním popisům, které byly zveřejněny na možná důležitých exemplářích, se mnoho z těchto vzorků stává „funkčními“ nomina dubia ".[61] Kromě toho bylo mnoho druhů pojmenováno na základě extrémně fragmentárních vzorků, které by nebyly vědecky příliš informativní, i kdyby byly dostatečně popsány. Více než třetina všech jmenovaných enantiornitových druhů je založena pouze na fragmentu jedné kosti. O'Connor a Dyke tvrdili, že zatímco tyto vzorky mohou pomoci rozšířit znalosti o časovém rozpětí nebo geografickém rozsahu Enantiornithů a je důležité je popsat, pojmenování těchto vzorků je „neospravedlnitelné“.[61]
Vztahy
Enantiornithes je sesterská skupina Euornithy a společně tvoří a clade volala Ornithothoraces (i když viz výše). Většina fylogenetické studie objevily Enantiornithes jako monofyletickou skupinu odlišnou od moderních ptáků a jejich nejbližších příbuzných. Fylogenetická analýza z roku 2002, kterou provedli Clarke a Norell, však snížila počet enantiornitheanů autapomorphies jen čtyři.[62]
Enantiornitheanská systematika je velmi prozatímní a notoricky obtížně studovatelná, vzhledem k tomu, že enantiornithové bývají extrémně extrémní homoplastický, nebo jsou si navzájem velmi podobné ve většině svých kosterních rysů díky konvergentní evoluci spíše než společnému původu.[30] To, co se nyní jeví jako docela jisté, je, že v rámci enantiornitheanů existovaly další členění, které možná kromě pokročilejších euenantiornithů zahrnovalo i některé menší bazální linie. Podrobnosti o vzájemném vztahu všech těchto linií, vlastně o platnosti většiny, jsou zpochybňovány, i když se zdá, že Avisauridae například představují platnou skupinu. Fylogenetičtí taxonomové dosud se zdráhali navrhovat vymezení enantiornithských subtypů.[63]
Jedno takové vymezení pojmenovalo Euenantiornithes, byla definována Chiappe (2002) jako zahrnující všechny druhy blíže k Sinornis než do Iberomesornis. Protože Iberomesornis je často považována za nejprimitivnější nebo bazální enantiornithean, Euenantiornithes může být extrémně inkluzivní skupina, složená ze všech Enantiornithes kromě Iberomesornis sám. Přesto, že je v souladu s fylogenetická nomenklatura, tato definice Euenantiornithes byla ostře kritizována některými vědci, jako např Paul Sereno, který to nazval „špatně definovaným“ clade [...] dobrý příklad špatné volby ve fylogenetické definici “.[63]
The kladogram níže byl nalezen analýzou Wanga et al. v roce 2015 aktualizováno z předchozí sady dat vytvořené Jingmai O'Connorem.[22]
| Ornithothoraces |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Seznam rodů
Incertae sedis
Enantiornithean taxonomie je obtížné vyhodnotit, a v důsledku toho několik subtypů ve skupině jsou důsledně nalezeny fylogenetickými analýzami. Většina enantiornitheans není zahrnuta v žádné konkrétní rodině a jako taková je zde uvedena. Mnoho z nich bylo považováno za euenantiornitheans, ačkoli kontroverze za tímto jménem znamená, že se ve studiích enantiornitheans nepoužívá důsledně.
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Abavornis | 1998 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z coracoids | ||
| Alethoalaornis | 2007 | Formace Jiufotang (Raná křída, Aptian ) | Špatně známé | ||
| Alexornis | 1976 | Formace La Bocana Roja (Pozdní křída, Campanian ) | Jeden z prvních známých enantiornitheanů. Kdysi byl považován za starodávného příbuzného válečky a datle | ||
| Avimaia | 2019 | Formace Xiagou (Pozdní křída, Aptian ) | Jeden exemplář z tohoto rodu zemřel s nepokládaným vejcem v těle |  | |
| Bauxitornis | 2010 | Csehbánya formace (Pozdní křída, Santonian ) | Fragmentární, ale jedinečný ve struktuře svého tarsometatarsusu |  | |
| Catenoleimus | 1998 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z a coracoid | ||
| Cathayornis | 1992 | Formace Jiufotang (Raná křída, Aptian ) | Jeden z prvních popsaných Jehol biota enantiornitheans. Známý z mnoha druhů, i když některé jsou nyní umístěny do svých vlastních rodů. Mohl mít podobný vzhled a životní styl jako a pitta |  | |
| Concornis | 1992 | Las Hoyas (Raná křída, Barremian ) | Jeden z nejúplnějších enantiornitheanů Las Hoyas |  | |
| Cratoavis[64] | 2015 | Santana formace (Raná křída, Aptian ) | Velmi zachovalý jihoamerický člen skupiny, doplněný ocasními pery ve tvaru pásky | ||
| Cruralispennia[32] | 2017 | Huajiying formace (Raná křída, Hauterivian ) | Měl neobvyklý pygostyl podobný ornituromorfům a stehenní peří. Jeden z nejstarších enantiornitheans |  | |
| Cuspirostrisornis | 1997 | Formace Jiufotang (Raná křída, Aptian ) | Původně se mylně věřilo, že vlastnil špičatý zobák | ||
| Dalingheornis | 2006 | Yixian formace (Raná křída, Aptian ) | Byl dobře přizpůsoben pro lezení kvůli jeho heterodaktylové nohy, jako ti z trogon | ||
| Dunhuangia[65] | 2015 | Formace Xiagou (Raná křída, Aptian ) | Enantiornithean z Changma pánve, oblast, která je neobvykle dominuje ornithuromorphs | ||
| Elbretornis | 2009 | Lecho formace (Pozdní křída, Maastrichtian ) | Známý pouze z křídel. Může být synonymem pro ostatní encyklopedie formace Lecho | ||
| Elektorornis | 2019 | Barmská jantar (Pozdní křída, Cenomanský ) | Známé z nohy uchované v jantaru s podlouhlým středním prstem | ||
| Elsornis | 2007 | Formace Djadochta (Pozdní křída, Campanian ) | Přestože je jeho kostra neúplná, má trojrozměrné zachování. Možná nelétavý díky rozměrům křídel | ||
| Enantiornis | 1981 | Lecho formace (Pozdní křída, Maastrichtian ) | Ačkoli je tento rod znám jen z několika kostí, je jmenovcem Enantiornithes. Byl to také jeden z největších a posledních představitelů skupiny před jejich zánikem | ||
| Eoalulavis | 1996 | Las Hoyas (Raná křída, Barremian ) | Zachovává peří včetně an alula, speciální typ peří, které řídí proudění vzduchu přes křídlo | ||
| Eocathayornis | 2002 | Formace Jiufotang (Raná křída, Aptian ) | Poté, co byl považován za bazálního blízkého příbuzného Cathayornis, i když je nyní považován za vzdáleněji příbuzný | ||
| Eoenantiornis | 1999 | Yixian formace (Raná křída, Aptian ) | Zachovalý, ale nekonzistentní ve fylogenetickém umístění |  | |
| Evgenavis | 2014 | Ilek formace (Raná křída, Barremian ) | Známý pouze z tarsometatarsus, který sdílí některé rysy s vlastnostmi enantiornitheans | ||
| Explorornis | 1998 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z coracoids | ||
| Falcatakely | 2020 | Formace Maevarano (Pozdní křída, Maastrichtian ) | Vyvinul se mohutný čenich s jediným zubem, navzdory zachování „primitivního“ uspořádání lebky na rozdíl od moderních ptáků | ||
| Feitianius[31] | 2015 | Formace Xiagou (Raná křída, Aptian ) | Měl propracovanou sadu ocasních peří, na rozdíl od spárovaných stužkových peří většiny enantiornitheans | ||
| Flexomornis | 2010 | Formace Woodbine (Pozdní křída, Cenomanský ) | Jeden z nejstarších nalezených severoamerických avialanů, i když známý pouze z úlomků | ||
| Fortunguavis[66] | 2014 | Formace Jiufotang (Raná křída, Aptian ) | Měl robustní kosti, včetně nohou a drápů, které mohly být přizpůsobeny pro lezení po stromech | ||
| Grabauornis[67] | 2015 | Yixian formace (Raná křída, Barremian ) | Proporce křídel tohoto enantiornitheana a přítomnost aluly naznačují, že to byl dobrý letec | ||
| Gracilornis | 2011 | Formace Jiufotang (Raná křída, Aptian ) | Možný příbuzný Cathayornis s charakteristicky štíhlými kostmi | ||
| Gurilynia | 1999 | Němegt formace (Pozdní křída, Maastrichtian ) | Špatně známý enantiornithean, ale evidentně velký a pozdě přežívající člen skupiny | ||
| Hollanda luceria[68] | 2010 | Barunská formace Goyot (Pozdní křída, Campanian ) | Původně identifikován jako ornithuromorph, ale od té doby znovu interpretován jako enantiornithean úzce související s Lectavis.[69] | ||
| Holbotia[70] | 2015 | Andaikhudag formace (Raná křída, Aptian ) | Považováno za malé ptakoještěr od svého objevu v roce 1977, dokud neobdržel formální popis v roce 2015. Vlastnil jedinečné krční obratle a primitivní patro | ||
| Houornis | 1997 | Formace Jiufotang (Raná křída, Aptian ) | Jakmile je považován za pochybný nebo za druh Cathayornis, ačkoli studie z roku 2015 ji považovala za platný rod[71] | ||
| Huoshanornis | 2010 | Formace Jiufotang (Raná křída, Aptian ) | Mohl to být velmi obratný leták díky struktuře jeho ruky a hrudní kosti | ||
| Iberomesornis | 1992 | Las Hoyas (Raná křída, Barremian ) | Jeden z prvních enantiornitheanů známých ze slušných pozůstatků. Také jeden z nejstarších a nejprimitivnějších členů skupiny |  | |
| Incolornis | 1998 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z coracoids. Jeden druh byl jednou považován za patřící k Enantiornis | ||
| Junornis[72] | 2017 | Yixian formace (Raná křída, Aptian ) | Tak dobře zachovalé, že jeho letový vzorec mohl být rekonstruován pomocí proporcí jeho peří a křídel |  | |
| Kizylkumavis | 1984 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha fragmentárních Bissekty enantiornitheans, známých pouze z a humerus fragment | ||
| Largirostrornis | 1997 | Formace Jiufotang (Raná křída, Aptian ) | Možná související s Cuspirostrisornis nebo synonymum Cathayornis | ||
| Lectavis | 1993 | Lecho formace (Pozdní křída, Maastrichtian ) | Velký a dlouhonohý člen skupiny, proporcionálně podobný moderním pobřežním ptákům | ||
| Lenesornis | 1996 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z a synsacrum fragment. Původně se považovalo za součást Ichthyornis | ||
| Liaoningornis | 1996 | Yixian formace (Raná křída, Aptian ) | Původně se věřilo, že je ornithuran, ale nyní se považuje za příbuzného Eoalulavis | ||
| Longchengornis | 1997 | Formace Jiufotang (Raná křída, Aptian ) | Může být synonymem pro Cathayornis | ||
| Martinavis | 2007 | Formace plazů Grès à, Lecho formace (Pozdní křída, Maastrichtian ) | Ačkoli byl tento rod znám pouze od humeri, byl velký a žil v širokém rozmezí | ||
| Microenantiornis | 2017 | Formace Jiufotang (Raná křída, Aptian ) | Malý člen skupiny, který měl několik primitivních a odvozených rysů ve srovnání s jinými enantiornitheans | ||
| Monoenantiornis[73] | 2016 | Yixian formace (Raná křída, Aptian ) | Známý z mladistvého exempláře, který popisuje, jak se různé rysy vyvíjely u enantiornitheanů v průběhu stárnutí | ||
| Nanantius | 1986 | Toolebuc formace (Raná křída, Albian ) | Fragmentární, ale mohl to být mořský pták, protože pozůstatky tohoto rodu byly nalezeny jako ichtyosaurus obsah střev | ||
| Noguerornis | 1989 | El Montsec (Raná křída, Barremian ) | Zachovává dojmy a propatagium, kožní chlopně na rameni, které tvoří část křídla | ||
| Orienantius | 2018 | Huajiying formace (Raná křída, Hauterivian ) | Mnoho detailů měkkých tkání vzorků z tohoto rodu bylo odhaleno UV zářením | ||
| Otogornis | 1993 | Formace Yijinholuo (Raná křída ) | Špatně známé | ||
| Paraprotopteryx | 2007 | Člen Qiaotou Huajiying formace (Raná křída, Aptian ?) | Zdánlivě měl čtyři ocasní pera připomínající stužku, místo pouhých dvou jako u většiny enantiornitheanů | ||
| Parvavis[74] | 2014 | Jiangdihe formace (Pozdní křída, Turonština na Santonian ) | Malý, ale v době své smrti plně dospělý. Jediný popsaný čínský enantiornithean se datuje do pozdní křídy | ||
| Piscivorenantiornis[75] | 2017 | Formace Jiufotang (Raná křída, Aptian ) | Známý z disartikulovaného skeletu dochovaného nad kusem obsahu žaludku složeného z rybích kostí, který mohl být jeho posledním jídlem | ||
| Protopteryx | 2000 | Huajiying formace (Raná křída, Hauterivian ) | Jeden z nejstarších a nejprimitivnějších členů skupiny |  | |
| Pterygornis[22] | 2016 | Formace Jiufotang (Raná křída, Aptian ) | Jedna disartikulovaná kostra tohoto rodu má zachovalé kosti lebky, včetně čtyřhlavého | ||
| Qiliania | 2011 | Formace Xiagou (Raná křída, Aptian ) | Některé z pozůstatků tohoto rodu zahrnují dobře zachovalé zadní končetiny. názvy druhů, Q. graffini, je pojmenována po Greg Graffin z kapely Špatné náboženství | ||
| Sazavis | 1989 | Formace Bissekty (Pozdní křída, Turonština na Coniacian ) | Jeden z mnoha dílčích Bissekty enantiornitheans, známý pouze z a tibiotarsus (holenní kost) | ||
| Shangyang | 2019 | Formace Jiufotang (Raná křída, Aptian ) | Neobvykle byly premaxillae tohoto rodu spojeny | ||
| Sinornis | 1992 | Formace Jiufotang (Raná křída, Aptian ) | Jeden z prvních popsaných Jehol biota enantiornitheans. Podobný Cathayornis ale obvykle se považují za odlišné |  | |
| Xiangornis | 2012 | Formace Jiufotang (Raná křída, Aptian ) | Ruka tohoto rodu byla podobná ruce ornithuromorphů, pravděpodobně skrz konvergentní evoluce. Velký člen skupiny | ||
| Yuanjiawaornis[76] | 2015 | Formace Jiufotang (Raná křída, Aptian ) | Jeden z největších enantiornitheanů známých ze slušných pozůstatků | ||
| Yungavolucris | 1993 | Lecho formace (Pozdní křída, Maastrichtian ) | Měl velký a neobvykle široký tarsometatarsal (kotníková kost) | 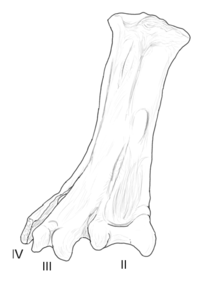 |
Longipterygidae
The Longipterygidae byla rodina raných křídel enantiornitheans s dlouhými čenichy, se zuby pouze na špičkách čenichu. Oni jsou obecně považováni za docela bazální členy skupiny.[28]
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Boluochia | 1995 | Formace Jiufotang (Raná křída, Aptian ) | Původně se mylně věřilo, že vlastnil zahnutý zobák | ||
| Camptodontornis | 2010 | Formace Jiufotang (Raná křída, Aptian ) | Původně volal Camptodontus, ačkoli tento rodový název je obsazen a brouk | ||
| Dapingfangornis | 2006 | Formace Jiufotang (Raná křída, Aptian ) | Možná měla na čele trnovitou strukturu | ||
| Longipteryx | 2001 | Formace Jiufotang (Raná křída, Aptian ) | Nejběžnější a nejznámější člen rodiny |  | |
| Longirostravis | 2004 | Yixian formace (Raná křída, Aptian ) | Stejně jako ostatní longipterygids, to posedlý tenký čenich, který může být použit pro sondování pro bezobratlé v bahně nebo kůře | ||
| Rapaxavis | 2009 | Formace Jiufotang (Raná křída, Aptian ) | Specializovaná na prohlížení díky struktuře nohou | ||
| Shanweiniao | 2009 | Yixian formace (Raná křída, Aptian ) | Získané vícenásobné ocasní peří, které mohlo být schopné generovat vztlak jako u moderních ptáků |  | |
| Shengjingornis | 2012 | Formace Jiufotang (Raná křída, Aptian ) | Velký člen rodiny |
Pengornithidae
The Pengornithidae byla rodina velkých raných enantiornitů. Měli četné malé zuby a četné primitivní rysy, které se ztratily u většiny ostatních enantiornitheans.[2] Některé studie tvrdí, že nemusí být vůbec enantiornitheans, ale spíše ornithuromorphs, blíže k moderním ptákům.
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Chiappeavis[30] | 2015 | Formace Jiufotang (Raná křída, Aptian ) | Měl ocas ve tvaru vějíře složený z mnoha peří | ||
| Eopengornis | 2014 | Huajiying formace (Raná křída, Hauterivian ) | Nejstarší známý člen rodiny a jeden z nejstarších známých domnělých enantiornithů. Vlastnil extrémně zachovalé ocasní pásky | ||
| Parapengornis[77] | 2015 | Formace Jiufotang (Raná křída, Aptian ) | Navrhl, aby měl datel podobný životní styl kvůli vlastnostem nohy a ocasu |  | |
| Pengornis | 2008 | Formace Jiufotang (Raná křída, Aptian ) | První objevený pengornithid a také jeden z největších enantiornitheanů známých ze slušných pozůstatků |
"Bohaiornithidae"
"Bohaiornithids „byli velcí, ale geologicky krátkodobí raní enantiornithové, s dlouhými zahnutými drápy a robustními zuby se zakřivenými špičkami. Mohli být ekvivalentem dravých ptáků, i když tento výklad je předmětem mnoha debat.[1] The monofilie této skupiny je pochybné a ve skutečnosti to může být evoluční stupeň.[78]
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Bohaiornis | 2011 | Formace Jiufotang (Raná křída, Aptian ) | Původně považován za dochovaný s gastrolity, i když později se zjistilo, že jde o minerální konkrementy | ||
| Gretcheniao | 2019 | Yixian formace (Raná křída, Barremian ) | Přizpůsobeno k létání, spíše než k letu. Může navrhovat parafyly nebo polyfyly z „Bohaiornithidae“ | ||
| Linyiornis[79] | 2016 | Formace Jiufotang (Raná křída, Aptian ) | Možný člen rodiny, známý z dobře zachované kostry doplněné strukturami, o nichž se předpokládá, že vyvíjejí vajíčka | ||
| Longusunguis | 2014 | Formace Jiufotang (Raná křída, Aptian ) | Docela typický člen rodiny |  | |
| Parabohaiornis | 2014 | Formace Jiufotang (Raná křída, Aptian ) | Blízký příbuzný Bohaiornis | ||
| Shenqiornis | 2010 | Člen Qiaotou v Huajiying formace (Raná křída, Aptian ?) | První známý člen rodiny, i když není považován za blízkého příbuzného Bohaiornis až o několik let později. Zachovává velkou postorbitální kost | ||
| Sulcavis | 2013 | Yixian formace (Raná křída, Aptian ) | Blízký příbuzný Shenqiornis s drážkou smalt na zubech, jedinečný mezi fosilními ptáky |  | |
| Zhouornis | 2013 | Formace Jiufotang (Raná křída, Aptian ) | Velký člen rodiny se zachovalou mozkovou skříní |
Gobipterygidae
Tato rodina může být monotypický (skládá se pouze z jednoho rodu nebo druhu), protože někteří členové skupiny jsou nejasní nebo špatně popsaní a mohou být synonymem pro její druh, Gobipteryx minuta.
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Gobipteryx | 1974 | Barunská formace Goyot (Pozdní křída, Campanian ) | Bezzubý pokročilý enantiornithean, který má robustní zobák, který se sbližoval s vývojem moderních ptáků | ||
| Jibeinia | 1997 | Člen Qiaotou v Huajiying formace (Raná křída, Aptian ?) | Špatně známé a popsané z kostry, která byla nyní ztracena. Může být synonymem pro Vescornis | ||
| Vescornis | 2004 | Člen Qiaotou v Huajiying formace (Raná křída, Aptian ?) | Malý a krátkosrstý enantiornithean, který může být synonymem Jibeinia |
Avisauridae
Avisauridae podléhá dvěma odlišným definicím různé inkluzivity. Zde je použita inkluzivnější definice, která následuje po Cau & Arduini (2008). Avisaurids byli dlouhotrvající a rozšířené enantiornitheans, které se vyznačují hlavně specifickými rysy jejich tarsometatarsals (kotníkové kosti). Největší a nejpokročilejší členové skupiny přežili v Severní a Jižní Americe až do konce křídy, přesto jsou ve srovnání s některými dřívějšími taxony velmi fragmentární.
| název | Rok | Formace | Umístění | Poznámky | snímky |
|---|---|---|---|---|---|
| Avisaurus | 1985 | Formace Hell Creek (Pozdní křída, Maastrichtian ) | Titulní avisaurid, stejně jako jeden z největších členů rodiny. Původně považován za non-avialanský dinosaurus |  | |
| Enantiophoenix | 2008 | Formace Ouadi al Gabour (Pozdní křída, Cenomanský ) | Mohl se živit mízou, jak byla zachována ve spojení s jantar korálky | ||
| Gettyia | 2018 | Dvě formace medicíny (Pozdní křída, Campanian ) | Nový rod pro Avisaurus gloriae | ||
| Halimornis | 2002 | Moorevilleova křída (Pozdní křída, Campanian ) | Bydleli by v pobřežním prostředí | ||
| Intnernis | 2010 | Formace Las Curtiembres (Pozdní křída, Campanian ) | Ačkoli úzce souvisí s některými z největších avisauridů, členové tohoto rodu byli velmi malí ptáci |  | |
| Mirarce | 2018 | Formace Kaiparowits (Pozdní křída, Campanian ) | Nejúplnější známý severoamerický avisaurid | ||
| Mystiornis | 2011 | Ilek formace (Raná křída, Barremian na Aptian ) | Má nesčetné množství funkcí z různých skupin Paraves, ačkoliv se nejvíce podobá avisauridům mezi vzorkovanými skupinami | ||
| Neuquenornis | 1994 | Formace Bajo de la Carpa (Pozdní křída, Santonian ) | Vlastnil dlouhá křídla a reverzní haluxy, což naznačuje dobrý let a prohlížení schopností | ||
| Soroavisaurus | 1993 | Lecho formace (Pozdní křída, Maastrichtian ) | Velmi blízký příbuzný Avisaurus |
Pochybné rody a pozoruhodné nepojmenované exempláře
- Gobipipus reshetovi: Popsáno v roce 2013 ze vzorků embryí ve skořápkách vajec z Barunská formace Goyot z Mongolsko. Tyto vzorky byly velmi podobné embryonálním Gobipteryx vzorky, ačkoli popisovatelé Gobipipus (skupina kontroverzních paleontologů včetně Evgeny Kurochkin a Sankar Chatterjee ) považují za odlišné.[80]
- Hebeiornis fengningensis: Synonymum pro Vescornis kvůli tomu, že byly popsány ze stejného vzorku. Přesto, že byl popsán v roce 1999, 5 let před popisem Vescornis, popis byl ve srovnání s popisem tak špatný Vescornis že většina autorů považuje toto jméno za prioritní. Výsledkem je název Hebeiornis je považován za nomen nudum („nahé jméno“).
- ‘’Proornis “Je pochybný pták ze Severní Koreje.
- "Liaoxiornis delicatus ": Popsáno v roce 1999 z enantiornithského vzorku nalezeného v Yixian formace. Tento exemplář byl původně považován za malého dospělého, ale později se zjistilo, že je mládě. Other specimens have henceforth been assigned to the genus. Due to a lack of distinguishing feature, many paleontologists have considered this genus an undiagnostic nomen dubium.
- LP-4450: A juvenile of an indeterminate enantiornithean from the El Montsec Vznik Španělsko. Its 2006 description studied the histologie of the skeleton, while later studies reported a squamosal bone present in the specimen but unknown in other enantiornitheans.
- IVPP V 13939: Briefly described in 2004, this Yixian enantiornithean had advanced pennaceous feathers on its legs, similar to (albeit shorter than) those of other paravians such as Mikroraptor a Anchiornis.[26]
- DIP-V-15100 a DIP-V-15101: Two different wings from hatchling specimens which were described in 2015. They attracted a significant amount of media attention upon their description. They were preserved in exceptional details due to having been trapped within Burmese amber for approximately 99 million years.[16]
- HPG-15-1: A partial corpse of a hatchling enantiornithean also preserved in Burmese amber. Although indeterminate, it attracted even more media attention than the two wings upon its description in 2017.[17]
- CUGB P1202: An indeterminate juvenile bohaiornithid z Formace Jiufotang. A 2016 analysis of its feathering found elongated putative melanosomes, suggesting that a large portion of its feathering was duhové.
- DIP-V-15102: Another corpse of an indeterminate hatchling preserved in Burmese amber. Described in early 2018.[18]
- MPCM-LH-26189 a/b: A partial skeleton of a hatchling from Las Hoyas in Spain, including both slab and counter-slab components. Its 2018 description revealed how various features developed in enantiornitheans as they aged. Such features include the zkostnatění z hrudní kost from various smaller bones, and the fusion of tail vertebrae into a pygostyle.
- YLSNHM01001: A foot and tail preserved in Burmese amber.[81]
Reference
- ^ A b C Wang, Min; Zhou, Zhong-He; O'Connor, Jingmai K.; Zelenkov, Nikita V. (2014). „Nová různorodá rodina enantiornithinů (Bohaiornithidae fam. Nov.) Ze spodní křídy v Číně s informacemi o dvou nových druzích“ (PDF). Vertebrata PalAsiatica. 52 (1): 31–76.
- ^ A b Wang, X .; O'Connor, J. K .; Zheng, X .; Wang, M .; Hu, H.; Zhou, Z. (2014). „Pohledy na vývoj rachis ovládaly ocasní pera z nového bazálního enantiornithinu (Aves: Ornithothoraces)“. Biologický žurnál společnosti Linnean. 113 (3): 805–819. doi:10.1111/bij.12313.
- ^ A b C Chiappe, Luis M.; Walker, Cyril A. (2002). "Skeletal Morphology and Systematics of the Cretaceous Euenantiornithes (Ornithothoraces: Enantiornithes)". In Chiappe, Luis M.; Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press. pp. 240–67. ISBN 978-0-520-20094-4.
- ^ Chiappe, Luis M. (2007). Glorified Dinosaurs: The Origin and Early Evolution of Birds. Hoboken, New Jersey: John Wiley and Sons. ISBN 978-0-471-24723-4.[stránka potřebná ]
- ^ O'Connor, Jingmai K.; Chiappe, Luis M.; Gao, Chunling; Zhao, Bo (2011). "Anatomy of the Early Cretaceous enantiornithine bird Rapaxavis pani" (PDF). Acta Palaeontologica Polonica. 56 (3): 463–75. doi:10.4202/app.2010.0047.
- ^ Elzanowski, Andrzej (1974). "Preliminary note on the palaeognathous bird from the Upper Cretaceous of Mongolia" (PDF). Palaeontologia Polonica. 29: 103–9.
- ^ A b Walker, C.A. (1981). "New subclass of birds from the Cretaceous of South America". Příroda. 292 (5818): 51–3. Bibcode:1981Natur.292...51W. doi:10.1038/292051a0.
- ^ Hope, Sylvia (2002). „Mezozoické záření Neornithů“. In Chiappe, Luis M.; Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press. pp. 339–88. ISBN 978-0-520-20094-4.
- ^ Feduccia, Alan (1996). Původ a vývoj ptáků. New Haven: Yale University Press. ISBN 978-0-300-06460-5.[stránka potřebná ]
- ^ Ty, Hai-lu; Lamanna, Matthew C .; Harris, Jerald D .; Chiappe, Luis M.; O'Connor, Jingmai; Ji, Shu-an; Lü, Jun-chang; Yuan, Chong-xi; Li, Da-qing; Zhang, Xing; Lacovara, Kenneth J .; Dodson, Peter; Ji, Qiang (16 June 2006). "A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China". Věda. 312 (5780): 1640–1643. Bibcode:2006Sci...312.1640Y. doi:10.1126/science.1126377. PMID 16778053.
- ^ A b Zhang, Zihui; Chiappe, Luis M.; Han, Gang; Chinsamy, Anusuya (2013). "A large bird from the Early Cretaceous of China: new information on the skull of enantiornithines". Časopis paleontologie obratlovců. 33 (5): 1176–89. doi:10.1080/02724634.2013.762708.
- ^ A b Zhou, Zhonghe; Clarke, Julia; Zhang, Fucheng (May 2008). "Insight into diversity, body size and morphological evolution from the largest Early Cretaceous enantiornithine bird". Anatomy Journal. 212 (5): 565–77. doi:10.1111 / j.1469-7580.2008.00880.x. PMC 2409080. PMID 18397240.
- ^ Hu, Dongyu; Xu, Xing; Hou, Lianhai; Sullivan, Corwin (2012). "A New Enantiornithine Bird from the Lower Cretaceous of Western Liaoning, China, and Its Implications for Early Avian Evolution". Časopis paleontologie obratlovců. 32 (3): 639–45. doi:10.1080/02724634.2012.652321.
- ^ Martin, Anthony J .; Vickers-Rich, Patricia; Rich, Thomas H .; Hall, Michael; Angielczyk, Kenneth (January 2014). "Nejstarší známé ptačí stopy z Austrálie: Eumeralla Formation (Albian), Dinosaur Cove, Victoria". Paleontologie. 57 (1): 7–19. doi:10.1111 / pala.12082.
- ^ Atterholt, Jessie; Hutchison, J. Howard; O’Connor, Jingmai K. (13 November 2018). „Nejúplnější enantiornithin ze Severní Ameriky a fylogenetická analýza Avisauridae“. PeerJ. 6: e5910. doi:10,7717 / peerj.5910. PMC 6238772. PMID 30479894.
- ^ A b C Xing, Lida; McKellar, Ryan C .; Wang, Min; Bai, Ming; O’Connor, Jingmai K.; Benton, Michael J .; Zhang, Jianping; Wang, Yan; Tseng, Kuowei; Lockley, Martin G .; Li, Gang; Zhang, Weiwei; Xu, Xing (28 June 2016). "Mummified precocial bird wings in mid-Cretaceous Burmese amber". Příroda komunikace. 7 (1): 12089. Bibcode:2016NatCo...712089X. doi:10.1038/ncomms12089. PMC 4931330. PMID 27352215.
- ^ A b Xing, Lida; O'Connor, Jingmai K.; McKellar, Ryan C .; Chiappe, Luis M.; Tseng, Kuowei; Li, Gang; Bai, Ming (September 2017). "A mid-Cretaceous enantiornithine (Aves) hatchling preserved in Burmese amber with unusual plumage". Výzkum v Gondwaně. 49: 264–277. Bibcode:2017GondR..49..264X. doi:10.1016/j.gr.2017.06.001.
- ^ A b Xing, Lida; O'Connor, Jingmai K.; McKellar, Ryan C .; Chiappe, Luis M.; Bai, Ming; Tseng, Kuowei; Zhang, Jie; Yang, Haidong; Fang, Jun; Li, Gang (February 2018). "A flattened enantiornithine in mid-Cretaceous Burmese amber: morphology and preservation". Vědecký bulletin. 63 (4): 235–243. doi:10.1016/j.scib.2018.01.019.
- ^ Xing, Lida; McKellar, Ryan C .; O’Connor, Jingmai K.; Bai, Ming; Tseng, Kuowei; Chiappe, Luis M. (30 January 2019). "A fully feathered enantiornithine foot and wing fragment preserved in mid-Cretaceous Burmese amber". Vědecké zprávy. 9 (1): 927. Bibcode:2019NatSR...9..927X. doi:10.1038/s41598-018-37427-4. PMC 6353931. PMID 30700773.
- ^ Xing, Lida; McKellar, Ryan C .; O'Connor, Jingmai K. (June 2020). "An unusually large bird wing in mid-Cretaceous Burmese amber". Křídový výzkum. 110: 104412. doi:10.1016/j.cretres.2020.104412.
- ^ A b O’Connor, Jingmai K.; Chiappe, Luis M. (28 February 2011). "A revision of enantiornithine (Aves: Ornithothoraces) skull morphology". Journal of Systematic Palaeontology. 9 (1): 135–157. doi:10.1080/14772019.2010.526639.
- ^ A b C Wang, Min; Hu, Han; Li, Zhiheng (21 August 2015). "A new small enantiornithine bird from the Jehol Biota, with implications for early evolution of avian skull morphology". Journal of Systematic Palaeontology. 14 (6): 481–497. doi:10.1080/14772019.2015.1073801.
- ^ Wang, Min; Hu, Han (January 2017). "A Comparative Morphological Study of the Jugal and Quadratojugal in Early Birds and Their Dinosaurian Relatives". Anatomický záznam. 300 (1): 62–75. doi:10.1002/ar.23446. PMID 28000410.
- ^ Chiappe, Luis M. (2009). "Downsized Dinosaurs: The Evolutionary Transition to Modern Birds". Evolution: Education and Outreach. 2 (2): 248–56. doi:10.1007/s12052-009-0133-4.
- ^ Becker, Rachel (28 June 2016). "Bird wings trapped in amber are a fossil first from the age of dinosaurs". Příroda. doi:10.1038/nature.2016.20162.
- ^ A b Zhang, Fucheng; Zhou, Zhonghe (October 2004). "Palaeontology: Leg feathers in an Early Cretaceous bird". Příroda. 431 (7011): 925. Bibcode:2004Natur.431..925Z. doi:10.1038/431925a. PMID 15496911.
- ^ A b Clarke, Julia A .; Zhou, Zhonghe; Zhang, Fucheng (March 2006). „Pohled na vývoj ptačího letu z nové kladu raně křídových ornithurinů z Číny a morfologie Yixianornis grabaui". Anatomy Journal. 208 (3): 287–308. doi:10.1111 / j.1469-7580.2006.00534.x. PMC 2100246. PMID 16533313.
- ^ A b O'Connor, Jingmai K.; Wang, Xuri; Chiappe, Luis M.; Gao, Chunling; Meng, Qingjin; Cheng, Xiaodong; Liu, Jinyuan (12 March 2009). "Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species". Časopis paleontologie obratlovců. 29 (1): 188–204. doi:10.1080/02724634.2009.10010371.
- ^ A b Chiappe, Luis M.; Bo, Zhao; O'Connor, Jingmai K.; Chunling, Gao; Xuri, Wang; Habib, Michael; Marugan-Lobon, Jesus; Qingjin, Meng; Xiaodong, Cheng (2014). "A new specimen of the Early Cretaceous bird Hongshanornis longicresta: pohledy na aerodynamiku a stravu bazálního ornithuromorph ". PeerJ. 2: e234. doi:10.7717/peerj.234. PMC 3898307. PMID 24482756.
- ^ A b C d O’Connor, Jingmai K.; Wang, Xiaoli; Zheng, Xiaoting; Hu, Han; Zhang, Xiaomei; Zhou, Zhonghe (January 2016). "An Enantiornithine with a Fan-Shaped Tail, and the Evolution of the Rectricial Complex in Early Birds". Aktuální biologie. 26 (1): 114–119. doi:10.1016 / j.cub.2015.11.036. PMID 26748849.
- ^ A b O’Connor, Jingmai K.; Li, Da-Qing; Lamanna, Matthew C .; Wang, Min; Harris, Jerald D .; Atterholt, Jessie; You, Hai-Lu (30 December 2015). „Nový raně křídový enantiornithin (Aves, Ornithothoraces) ze severozápadní Číny s komplikovanou ocasní výzdobou“. Časopis paleontologie obratlovců. 36 (1): e1054035. doi:10.1080/02724634.2015.1054035.
- ^ A b Wang, Min; O’Connor, Jingmai K.; Pan, Yanhong; Zhou, Zhonghe (2017-01-31). "A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle". Příroda komunikace. 8: 14141. Bibcode:2017NatCo...814141W. doi:10.1038/ncomms14141. PMC 5290326. PMID 28139644.
- ^ A b Sanz, José L .; Chiappe, Luis M.; Buscalioni, Angela D. (1995). "The Osteology of Concornis lacustris (Aves: Enantiornithes) from the Lower Cretaceous of Spain and a Reexamination of its Phylogenetic Relationships". Americké muzeum Novitates (3133): 1–23. hdl:2246/3667.
- ^ Dalla Vecchia, Fabio M .; Chiappe, Luis M. (2003). "First avian skeleton from the Mesozoic of northern Gondwana". Časopis paleontologie obratlovců. 22 (4): 856–60. doi:10.1671/0272-4634(2002)022[0856:FASFTM]2.0.CO;2. JSTOR 4524284.
- ^ O'Connor, Jingmai K. (1 January 2019). "The trophic habits of early birds". Paleogeografie, paleoklimatologie, paleoekologie. 513: 178–195. Bibcode:2019PPP...513..178O. doi:10.1016/j.palaeo.2018.03.006.
- ^ O'Connor, Jingmai K.; Zhou, Zhonghe; Smith, Andrew (6 November 2019). "The evolution of the modern avian digestive system: insights from paravian fossils from the Yanliao and Jehol biotas". Paleontologie. 63 (1): 13–27. doi:10.1111/pala.12453.
- ^ Houston, David C .; Copsey, J. A. (1994). "Bone digestion and intestinal morphology of the Bearded Vulture". The Journal of Raptor Research. 28 (2): 73–78.
- ^ Sanz, José L .; Chiappe, Luis M.; Fernádez-Jalvo, Yolanda; Ortega, Francisco; Sánchez-Chillón, Begoña; Poyato-Ariza1, Francisco J.; Pérez-Moreno, Bernardino P. (February 2001). "An early Cretaceous pellet". Příroda. 409 (6823): 998–1000. Bibcode:2001Natur.409..998S. doi:10.1038/35059172. PMID 11234054.
- ^ Mikhailov, Konstantin E. (1991). "Classification of fossil eggshells of amniotic vertebrates" (PDF). Acta Palaeontologica Polonica. 36 (2): 193–238.
- ^ Mikhailov, Konstantin E. (1996). "New Genera of Fossil Eggs from the Upper Cretaceous of Mongolia". Paleontologický deník. 30 (2): 246–8.
- ^ Elżanowski, Andrzej (1981). "Embryonic bird skeletons from the late Cretaceous of Mongolia" (PDF). Palaeontologia Polonica. 42: 147–79.
- ^ Sanz, José L .; Chiappe, Luis M.; Pérez-Moreno, Bernardino P.; Moratalla, José J .; Hernández-Carrasquilla, Francisco; Buscalioni, Angela D.; Ortega, Francisco; Poyato-Ariza, Francisco J.; Rasskin-Gutman, Diego; Martı́nez-Delclòs, Xavier (June 6, 1997). "A Nestling Bird from the Lower Cretaceous of Spain: Implications for Avian Skull and Neck Evolution". Věda. 276 (5318): 1543–6. doi:10.1126/science.276.5318.1543.
- ^ Zhou, Zhonghe; Zhang, Fucheng (October 22, 2004). "A Precocial Avian Embryo from the Lower Cretaceous of China". Věda. 306 (5696): 653. doi:10.1126/science.1100000. PMID 15499011.
- ^ A b C Chiappe, Luis M.; Shu'an, Ji; Qiang, Ji (2007). "Juvenile Birds from the Early Cretaceous of China: Implications for Enantiornithine Ontogeny". Americké muzeum Novitates. 3594: 1–46. doi:10.1206/0003-0082(2007)3594[1:JBFTEC]2.0.CO;2. hdl:2246/5890.
- ^ Elżanowski, Andrzej (1995). "Cretaceous birds and avian phylogeny". Courier Forschungsinstitut Senckenberg. 181: 37–53.
- ^ Kurochkin, E.N .; Chatterjee, S .; Mikhailov, K.E. (2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontologický deník. 47 (11): 1252–69. doi:10.1134/S0031030113110087.
- ^ Kurochkin, E. N .; Chatterjee, S .; Mikhailov, K. E. (19 December 2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontologický deník. 47 (11): 1252–1269. doi:10.1134/S0031030113110087.
- ^ Cambra-Moo, Oscar; Buscalioni, Ángela Delgado; Cubo, Jorge; Castanet, Jacques; Loth, Marie-Madeleine; de Margerie, Emmanuel; de Ricqlès, Armand (2006). "Histological observations of Enantiornithine bone (Saurischia, Aves) from the Lower Cretaceous of Las Hoyas (Spain)". Comptes Rendus Palevol. 5 (5): 685–91. doi:10.1016/j.crpv.2005.12.018.
- ^ O'Connor, Jingmai K.; Wang, Min; Zheng, Xiao-Ting; Wang, Xiao-Li; Zhou, Zhong-He (2014). "The histology of two female Early Cretaceous birds" (PDF). Vertebrata PalAsiatica. 52 (1): 112–28.
- ^ Chiappe, L.M. (1995). "The phylogenetic position of the Cretaceous birds of Argentina: Enantiornithes and Patagopteryx deferrariisi". Courier Forschungsinstitut Senckenberg. 181: 55–63.
- ^ Dyke, Gareth; Vremir, Mátyás; Kaiser, Gary; Naish, Darren (June 2012). „Utopená druhohorní chovatelská kolonie ptáků z pozdní křídy Transylvánie“. Die Naturwissenschaften. 99 (6): 435–42. Bibcode:2012NW.....99..435D. CiteSeerX 10.1.1.394.9006. doi:10.1007 / s00114-012-0917-1. PMID 22575918.
- ^ Fernández, Mariela S.; García, Rodolfo A.; Fiorelli, Lucas; Scolaro, Alejandro; Salvador, Rodrigo B .; Cotaro, Carlos N.; Kaiser, Gary W .; Dyke, Gareth J .; Farke, Andrew A. (17 April 2013). "A Large Accumulation of Avian Eggs from the Late Cretaceous of Patagonia (Argentina) Reveals a Novel Nesting Strategy in Mesozoic Birds". PLOS ONE. 8 (4): e61030. Bibcode:2013PLoSO...861030F. doi:10.1371/journal.pone.0061030. PMC 3629076. PMID 23613776.
- ^ O'Connor, Jingmai K.; Falk, Amanda; Wang, Min; Zheng, Xiao-Ting (2020). "First report of immature feathers in juvenile enantiornithines from the Early Cretaceous Jehol avifauna". Vertebrata PalAsiatica. 58: 24–44. doi:10.19615/j.cnki.1000-3118.190823.
- ^ Padian, Kevin; Chiappe, Luis M. (11 January 2007). "The origin and early evolution of birds". Biologické recenze. 73 (1): 1–42. doi:10.1111/j.1469-185x.1997.tb00024.x.
- ^ Zhou, Shuang; Zhou, Zhong-He; O'Connor, Jingmai K. (2012). "New, basal beaked, ornithurine, pták, od, ta, dolní, křídový, o, western, Liaoning, China" (PDF). Vertebrata PalAsiatica. 50 (1): 9–24. Shrnutí ležel – Phys.org (February 13, 2012).
- ^ Houde, Peter W. (1988). "Paleognathous Birds from the Early Tertiary of the North Hemisphere". Publications of the Nuttall Ornithological Club (Cambridge Massachusetts, USA: Nuttall Ornithological Club) 22
- ^ Navalón, Guillermo; Marugán-Lobón, Jesús; Chiappe, Luis M.; Luis Sanz, José; Buscalioni, Ángela D. (6 October 2015). "Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight". Vědecké zprávy. 5 (1): 14864. Bibcode:2015NatSR...514864N. doi:10.1038/srep14864. PMC 4594305. PMID 26440221.
- ^ Wang, Xia; McGowan, Alistair J.; Dyke, Gareth J .; Turvey, Samuel T. (7 December 2011). "Avian Wing Proportions and Flight Styles: First Step towards Predicting the Flight Modes of Mesozoic Birds". PLOS ONE. 6 (12): e28672. Bibcode:2011PLoSO...628672W. doi:10.1371/journal.pone.0028672. PMC 3233598. PMID 22163324.
- ^ Chiappe, Luis M.; Suzuki, Shigeru; Dyke, Gareth J .; Watabe, Mahito; Tsogtbaatar, K .; Barsbold, Rinchen (January 2007). "A new Enantiornithine bird from the Late Cretaceous of the Gobi desert". Journal of Systematic Palaeontology. 5 (2): 193–208. doi:10.1017/S1477201906001969.
- ^ Zheng, Xiaoting; Wang, Xiaoli; O'Connor, Jingmai; Zhou, Zhonghe (9. října 2012). "Insight into the early evolution of the avian sternum from juvenile enantiornithines". Příroda komunikace. 3 (1): 1116. Bibcode:2012NatCo ... 3.1116Z. doi:10.1038 / ncomms2104. PMID 23047674.
- ^ A b O'Connor, Jingmai; Dyke, Gareth (2010). „Přehodnocení Sinornis santensis a Cathayornis yandica (Aves: Enantiornithes)". Záznamy o australském muzeu. 62: 7–20. doi:10.3853/j.0067-1975.62.2010.1540.
- ^ Clarke, Julia A .; Norell, Mark A. (2002). "The Morphology and Phylogenetic Position of Apsaravis ukhaana z pozdní křídy Mongolska “. Americké muzeum Novitates. 3387: 1–46. CiteSeerX 10.1.1.693.8475. doi:10.1206/0003-0082(2002)387<0001:TMAPPO>2.0.CO;2.
- ^ A b Sereno, P.C. (2005) TaxonSearch: Stem Archosauria Archivováno 2007-02-19 na Wayback Machine. Version 1.0, 2005-NOV- 7. Retrieved 2006-OCT-02.
- ^ Carvalho; Novas; Agnolín; Isasi; Freitas; Andrade (2015). "A new genus and species of enantiornithine bird from the Early Cretaceous of Brazil". Brazilian Journal of Geology. 45 (2): 161–171. doi:10.1590/23174889201500020001.
- ^ Wang, Li; O'Connor, Zhou; You (2015). "Druhý druh enantiornithinového ptáka z dolní křídy Changma Basin, severozápadní Čína s důsledky pro taxonomickou rozmanitost Changma avifauna". Křídový výzkum. 55: 56–65. doi:10.1016 / j.cretres.2015.01.008.
- ^ Wang, M .; O'Connor, J. K .; Zhou, Z. (2014). "Nový robustní enantiornithinový pták ze spodní křídy v Číně se skenovacími úpravami". Časopis paleontologie obratlovců. 34 (3): 657–671. doi:10.1080/02724634.2013.812101.
- ^ Dalsätt, J.; Ericson, P. G.; Zhou, Z. (2015). „New Enantiornithes (Aves) from the Early Cretaceous of China“. Acta Geologica Sinica. 86 (2): 801–807. doi:10.1111/1755-6724.12270.
- ^ Bell, Alyssa K.; Chiappe, Luis M.; Erickson, Gregory M.; Suzuki, Shigeru; Watabe, Mahito; Barsbold, Rinchen; Tsogtbaatar, K. (February 2010). "Description and ecologic analysis of Hollanda luceria, a Late Cretaceous bird from the Gobi Desert (Mongolia)". Křídový výzkum. 31 (1): 16–26. doi:10.1016 / j.cretres.2009.09.001.
- ^ Hartman, Scott; Mortimer, Mickey; Wahl, William R .; Lomax, Dean R .; Lippincott, Jessica; Lovelace, David M. (10 July 2019). „Nový paravianský dinosaurus z Late Jurassic v Severní Americe podporuje pozdní akvizici ptačího letu“. PeerJ. 7: e7247. doi:10,7717 / peerj.7247. PMC 6626525. PMID 31333906.
- ^ Zelenkov, Nikita V.; Averianov, Alexander O. (13 June 2015). "Historický exemplář enantiornithinového ptáka ze starší křídy Mongolska představující nový taxon se specializovanou morfologií krku". Journal of Systematic Palaeontology. 14 (4): 319–338. doi:10.1080/14772019.2015.1051146.
- ^ Wang, M .; Liu, D. (2015). "Taxonomické přehodnocení Cathayornithidae (Aves: Enantiornithes)". Journal of Systematic Palaeontology. 14: 1–19. doi:10.1080/14772019.2014.994087.
- ^ Liu, Di; Chiappe, Luis M.; Serrano, Francisco; Habib, Michael; Zhang, Yuguang; Meng, Qinjing; Shawkey, Matthew (11 October 2017). "Flight aerodynamics in enantiornithines: Information from a new Chinese Early Cretaceous bird". PLOS One. 12 (10): e0184637. Bibcode:2017PLoSO..1284637L. doi:10.1371/journal.pone.0184637. PMC 5636078. PMID 29020077.
- ^ Hu, Han; O'Connor, Jingmai K. (14 November 2016). „První druh Enantiornithes ze Sihedangu objasňuje vývoj skeletu ve starokřídových enantiornithinech“. Journal of Systematic Palaeontology. 15 (11): 909–926. doi:10.1080/14772019.2016.1246111.
- ^ Wang, Min; Zhou, Zhonghe; Xu, Guanghui (7 January 2014). "The first enantiornithine bird from the Upper Cretaceous of China". Časopis paleontologie obratlovců. 34 (1): 135–145. doi:10.1080/02724634.2013.794814.
- ^ Wang, Min; Zhou, Zhonghe (12 April 2017). "A morphological study of the first known piscivorous enantiornithine bird from the Early Cretaceous of China". Časopis paleontologie obratlovců. 37 (2): e1278702. doi:10.1080/02724634.2017.1278702.
- ^ Hu, Dongyu; Liu, Ying; Li, Jinhua; Xu, Xing; Hou, Lianhai (July 2015). "Yuanjiawaornis viriosus, gen. et sp. nov., a large enantiornithine bird from the Lower Cretaceous of western Liaoning, China". Křídový výzkum. 55: 210–219. doi:10.1016 / j.cretres.2015.02.013.
- ^ Hu, Han; O’Connor, Jingmai K.; Zhou, Zhonghe; Farke, Andrew A. (3 June 2015). "A New Species of Pengornithidae (Aves: Enantiornithes) from the Lower Cretaceous of China Suggests a Specialized Scansorial Habitat Previously Unknown in Early Birds". PLOS One. 10 (6): e0126791. Bibcode:2015PLoSO..1026791H. doi:10.1371/journal.pone.0126791. PMC 4454694. PMID 26039693.
- ^ Chiappe, Luis M.; Qingjin, Meng; Serrano, Francisco; Sigurdsen, Trond; Min, Wang; Bell, Alyssa; Di, Liu (25 October 2019). "Nový Bohaiornis-like bird from the Early Cretaceous of China: enantiornithine interrelationships and flight performance". PeerJ. 7: e7846. doi:10.7717/peerj.7846. PMC 6816414. PMID 31667014.
- ^ Wang, Yan; Wang, Min; O'Connor, Jingmai K.; Wang, Xiaoli; Zheng, Xiaoting; Zhang, Xiaomei (11 January 2016). „Nový Jehol enantiornithine pták s trojrozměrnou konzervací a ovariálními folikuly“. Časopis paleontologie obratlovců. 36 (2): e1054496. doi:10.1080/02724634.2015.1054496.
- ^ Kurochkin, E. N .; Chatterjee, S .; Mikhailov, K. E. (19 December 2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontologický deník. 47 (11): 1252–1269. doi:10.1134 / s0031030113110087.
- ^ Xing, Lida; McKellar, Ryan C .; O’Connor, Jingmai K.; Niu, Kecheng; Mai, Huijuan (29. října 2019). "A mid-Cretaceous enantiornithine foot and tail feather preserved in Burmese amber". Vědecké zprávy. 9 (1): 15513. Bibcode:2019NatSR...915513X. doi:10.1038/s41598-019-51929-9. PMC 6820775. PMID 31664115.
externí odkazy
![]() Média související s Enantiornithes na Wikimedia Commons
Média související s Enantiornithes na Wikimedia Commons

















| Anatomie | |||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chování | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Vývoj | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fosilní ptáci | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Člověk interakce | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Seznamy | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Neornithes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||