Adasaurus - Adasaurus
| Adasaurus | |
|---|---|
 | |
| Rekonstruovaná lebka z holotypu | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Clade: | Dinosaurie |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Rodina: | †Dromaeosauridae |
| Clade: | †Eudromaeosauria |
| Podčeleď: | †Velociraptorinae |
| Rod: | Adasaurus Barsbold, 1983 |
| Zadejte druh | |
| †Adasaurus mongoliensis Barsbold, 1983 | |
Adasaurus (/ˌɑːdəˈsɔːrəs/ AH-də-SAWR-əs; což znamená „ještěrka Ada“) je a rod z dromaeosaurid dinosaurus který žil v Asie Během Pozdní křída období asi před 70 miliony let. Rod je znám ze dvou dílčích vzorků nalezených v Němegt formace z Mongolsko které byly částečně popsány v roce 1983 paleontologem Rinchen Barsbold.
Jednalo se o středně velký dromaeosaurid, který byl dlouhý asi 2,4 m (7,9 ft) a váží 36,4 kg (80 lb). Na rozdíl od jiných dromaeosauridů Adasaurus vyvinul poměrně malý a tupý srpový dráp, který měl pravděpodobně menší použití a zakřivený slzná kost; tato druhá vlastnost je také sdílena s nepříbuznými Austroraptor. I když se srpový dráp zmenšil, zachoval si charakteristické zaoblené členění většiny dromaeosauridů.
Adasaurus byl původně považován za dromaeosaurin Barsbold, skupina, která zahrnuje robustní dromaeosaury s hlubokými čelistmi. Revize provedené na exemplářích ukázaly, že tento dromaeosaurid patří do Velociraptorinae který se skládá z lehce stavěných zvířat jako Velociraptor.
Historie objevů

Adasaurus byl poprvé objeven 1977 podle mongolský paleontolog Rinchen Barsbold na pánevním srovnání s ostatními teropody, ale zůstal by jako neformálně pojmenovaný taxon až do řádného popisu.[1] V roce 1983 Barsbold vydal velkou srovnávací revizi známého mongolský theropod taxony v době, kdy formálně jmenoval Adasaurus a druh druhu A. mongoliensis, který byl založen na dvou dílčích vzorcích. The druhové jméno, Adasaurus, je převzato z zlý duch Ada v mytologie Mongolska a řecký slovo σαῦρος (sauros, což znamená ještěrka). The konkrétní název pro jednotlivé druhy, mongoliensis, odkazuje na zemi objevu Mongolsko. Barsbold stručně popsal Adasaurus jako dromaeosaurid a poznamenal, že tento nový taxon vlastnil výrazně snížený druhý pedál ungual. Vzhledem k tomu, že tento rys kontrastoval s velkým, ostře vyvinutým unguálem většiny členů, Barsbold jej uvedl jako unikátní znak pro Adasaurus.[2] O autentičnosti této neobvyklé redukce však v roce 2010 pochyboval Phil Senter, který tvrdil, že domnělý ungual se nevztahuje k exempláři.[3] Nicméně v revidované diagnóze provedené Turnerem a kolegy v roce 2012 je tento znak stále považován za autentický,[4] který byl široce sledován jinými autory.[5][6]
Adasaurus je známo z holotyp MPC-D 100/20, což představuje dospělého jedince kompromitujícího dílčí část lebka chybí přední oblast, vpravo scapulocoracoid a a sternální dlaha, 8 krčních obratlů, 11 dílčích hřbetních obratlů, křížová kost, 7 ocasních obratlů, částečné zadní končetiny s pravou nohou a téměř úplné pravé pánevní pletenec kompromitující ilium, ischium a pubis. Druhý vzorek je reprezentován méně úplným paratyp MPC-D 100/21, který zahrnuje dva ocasní obratle a téměř úplnou pravou nohu. Oba vzorky byly objeveny z Němegt formace v lokalitě Bügiin Tsav, v poušť Gobi Mongolska.[2][7][4][5] V roce 2004 Philip J. Currie a David J. Varricchio odkázal na další dva vzorky Adasaurus označené jako IGM 100/22 a IGM 100/23. Předpokládá se, že vzorky byly získány ze stejné lokality holotypu.[8] Tyto vzorky jsou však ve skutečnosti známy z lokalit Shine Us Khuduk a Tel Ulan Chaltsai (v uvedeném pořadí) Formace Bayan Shireh a jsou tedy starší než pozůstatky Adasaurus od mladších Němegt formace. Představují odlišný a nový taxon, který se liší od Adasaurus.[9][4]
Popis

Adasaurus byl středně velký dromaeosaurid. Holotyp má odhadovanou délku 2,4 m (7,9 ft) s hmotností 36,4 kg (80 lb).[10] Poměrně větší pedálové prvky paratypu ukazují na mírně větší velikost v tomto druhém vzorku.[5] Kromě sníženého pedálu ungual II, Adasaurus lze rozpoznat podle následujících dalších znaků: rozšířená projekce maxilární; zakřivený slzný; dolní čelist s výrazným surangular foramen; nepravidelný trojúhelníkový výstupek na kvadrát hřídel; pleurocoels jsou přítomny na přední straně křížové obratle; a přední okraj přední čepele v ilium je relativně zkrácen.[4]
Na pravé straně lebka, spodní část jugal je rozšířen shora dolů. Kvadrát je velká a svislá kost s velkým trojúhelníkovým tvarem projekce na jeho boční hranici. Tato trojúhelníková projekce je umístěna na kvadrátovém hřídeli a je ohnutá směrem nahoru. Horní povrch vpravo ektopterygoid - malá kost patra - je zploštělá k patro. Stejně jako u jiných dromaeosauridů má slz obrácený tvar L, ale tenké tělo této kosti je zakřivené, což je také vidět na Austroraptor.[4] The lopatka a coracoid holotypu jsou zcela spojeny a dávají formu scapulocoracoid a steh mezi nimi není přítomen.[5] Pneumatický foramina jsou přítomny v holotypických předních křížových obratlích.[4][5]
The stehenní kost a holenní kost holotypu měří 27,3 cm (273 mm), respektive 30,3 cm (303 mm),[5] a čtvrtý trochanter je prominentní a drsný hřeben, který je umístěn na zadní vnitřní ploše horní oblasti stehenní dříky. Samotná stehenní kost je velmi podobná té neurčité dromaeosaur DGBU-78.[11] Přední povrch dolního konce stehenní kosti má konvexní tvar a boční tuberkulóza horního konce kloubu tibia-fibula je kompaktní konstrukce.[12] Přítomnost zaoblené jámy na vnitřním povrchu spodní části tibiotarsus je často dokumentována u dromaeosauridů, avšak u této funkce není ověřitelná Adasaurus od astragalus oblast je pokryta sedimenty.[5]
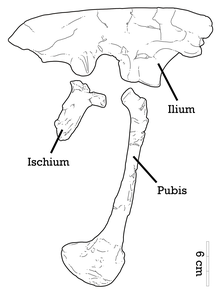
Zadní horní okraj kyčelního kloubu je proporcionálně více zesílen než zadní Achillobátor,[12] a přední okraj přední čepele ilia má podobný tvar jako Saurornitholestes. Tato přední hranice má vroubkované aparát, který je charakteristický Adasaurus. Jako celek je horní okraj narovnán do tvaru. Pubiální stopka - robustní přední extenze, která artikuluje s pubisem - je široká a vyvinutá až ke dnu. Na iliu chybí velké supratrochanterické prodloužení (nad trochanterem stehenní kosti). Stejně jako ostatní dromaeosaurids, pubis je protáhlý s rozšířeným stydké boty (dolní konec) a má opisthopubic (dozadu směřující) stav.[4] Číslice II ungual není hypertrofovaná (protáhlá) jako u většiny dromaeosauridů,[13][4] a ačkoli Adasaurus má podobný metatarzální poměr II-III jako u metatarzu Balaur, je to kvůli zmenšenému srpkovitému drápu číslice II namísto protáhlého úhlu číslice I. Metatarzální III paratypu ukazuje, že nádor je přítomen na povrchu extenzoru a tato tuberosita pravděpodobně pochází z vložení sval tibialis cranialis.[5] Dolní tarzály a horní konce metatarzů jsou poněkud spojené.[7][4]
Klasifikace
Adasaurus je členem Dromaeosauridae, skupina, která úzce souvisí s životem ptactvo. Když byl postaven Barsboldem v roce 1983, Velociraptorinae byl koncipován jako skupina obsahující Velociraptor a blízce příbuzné druhy, které se vyznačovaly menší velikostí a dlouhými úzkými čenichy. Barsbold to však nezahrnul Adasaurus ve skupině to místo toho umístil do Dromaeosaurinae.[2] Teprve v roce 1998 byla tato skupina definována jako clade Paul Sereno. Sereno definoval skupinu jako všechny dromaeosauridy, které mají bližší vztah Velociraptor než do Dromaeosaurus.[14] Tradiční pohled na Velociraptorinae běžně zahrnutý Velociraptor, Tsaagan a Linheraptor, které jsou známé z kompletních lebek, se však většina analýz velmi liší, pokud jde o to, které druhy jsou velociraptoriny ve skutečnosti a které dromaeosauriny. Turner a kolegové v roce 2012 podpořili tradiční monofyletickou skladbu Velociraptorinae.[4] Některé studie však nalezly ve Velociraptorinae velmi odlišnou skupinu dromaeosauridů, jako je Longrich a Currie v roce 2009, které se zotavily Deinonychus mimo Velociraptorine a Dromaeosaurinae.[15] Tradičně, Adasaurus byl přidělen k Dromaeosaurinae, který zahrnuje obří, silně stavěná zvířata jako např Achillobátor a Utahraptor[16] ale několik analýz naznačuje, že patří k Velociraptorinae namísto.[17][18][19]

Níže je a kladogram založeno na fylogenetická analýza provedli Powers a kolegové v roce 2020.[20]
| Velociraptorinae |
| |||||||||||||||||||||||||||||||||
Paleobiologie
Paleopatologie
V roce 1997 Norell a Makovicky uvedli, že holotypový exemplář Adasaurus představuje do značné míry patologické (kvůli zranění nebo choroba ) individuální.[13] Toto pozorování znovu potvrdili v roce 2004 tvrzením, že pánev je patologická.[21] Během rozsáhlé revize Dromaeosauridae Turnerem a kolegy v roce 2012 však byl holotyp znovu prozkoumán a bylo zjištěno, že není patologický. Místo toho, aby odrážely zraněné prvky, může několik povrchů představovat pokročilý věk jedince, jako je například roztavený horní konec metatarzu.[4]
Funkce Sicke Claw

V roce 2011 Denver Fowler a kolegové navrhli novou metodu, pomocí které si dromaeosauridy mohli vzít menší kořist. Tento predátorský model „Raptor Prey Restraint“ (RPR) navrhuje, aby dromaeosaurids zabil svou kořist tím, že skočil do jejich lomu, přitiskl ji pod svou tělesnou váhu a pevně ji sevřel pomocí velkých srpkovitých drápů číslice pedálu II - v způsobem velmi podobným existujícím accipitrid draví ptáci. Stejně jako accipitridy by se dromaeosaurid začal živit ještě živým zvířetem, dokud nakonec nezemřel ztráta krve a selhání orgánů. Tento návrh je založen především na srovnání morfologie a proporcí chodidel a končetin dromaeosauridů s několika skupinami existujících dravých ptáků s poměrně známým dravým chováním. Fowler a collaegues zjistili, že nohy a nohy dromaeosauridů se nejvíce podobají nohám a nohám Orli a jestřábi, zejména pokud jde o zvětšený druhý dráp a podobný rozsah uchopovacího pohybu, ale krátký metatarzu a síla chodidla by byla více podobná síle nohy sovy. Model RPR by byl v souladu s dalšími aspekty anatomie dromaeosauridů, jako je jejich neobvyklý chrup a morfologie paží. Paže byly pokryty dlouhým peřím a mohly být použity jako klopení stabilizátorů pro rovnováhu, zatímco na vrcholu bojující kořisti, spolu s tuhým vyvážením ocasu. A konečně, srovnatelně slabé čelisti by byly užitečné pro konzumaci kořisti naživu, ale ne tak užitečné pro násilné odeslání kořisti.[22]

V roce 2019 Peter Bishop rekonstruoval kostru nohy a svalstvo Deinonychus pomocí trojrozměrných modelů svaly, šlachy, a kosti. S přidáním matematických modelů a rovnic Bishop simuloval podmínky, které by poskytovaly maximální sílu na špičce srpového drápu a tedy nejpravděpodobnější funkci. Mezi navrhované způsoby použití srpového drápu patří: kopání do řezu, sekání nebo vykořisťování kořisti; za uchopení na bocích kořisti; piercing podporovaný tělesnou hmotností; zaútočit na životně důležité oblasti kořisti; zadržovat kořist; uvnitř - nebo mezidruhová soutěž; a vykopat kořist z úkrytů. Výsledky získané Bishopem ukázaly, že přikrčená poloha zvýšila drápové síly, ale tyto síly zůstaly relativně slabé, což naznačuje, že drápy nebyly dostatečně silné, aby mohly být použity při sekání úderů. Spíše než kosatky se používaly na sekání, bylo pravděpodobnější, že budou užitečné v ohnutých úhlech nohou, jako je zadržování kořisti a bodnutí kořisti v těsné blízkosti. Tyto výsledky jsou v souladu s „Boj s dinosaury "vzorek, který zachovává a Velociraptor a Protoceratops uzamčen v boji, přičemž první sevřel drápy v nevytažené poloze nohou. Navzdory dosaženým výsledkům měl Bishop za to, že schopnosti srpového drápu se mohly v rámci taxonů lišit, protože mezi dromaeosauridy Adasaurus měl neobvykle menší srpový dráp, který si udržel charakteristický ginglymoid - strukturu rozdělenou na dvě části - a hyperextensibilní kloubní povrch předposlední falangy. Nemohl potvrdit ani ignorovat, že číslice pedálu II může ztratit nebo si ji funkčně udržet.[6] Kubota a Barsbold dříve uvedli, že vysoce redukovaný srpový dráp Adasaurus mohly být používány s menší frekvencí než jiné deinonychosaurů jako spodní povrch spodní paty na předposlední falanga nemá žádné zjevné asymetrické hřebeny jako ostatní dromaeosaurids a troodontids.[7]
Paleoenvironment
Adasaurus je známo z Pozdní křída Němegt formace, jehož věk byl považován za pozdní Campanian do raného středu Maastrichtian etapy, asi 70 milionů a 68 před miliony let. Prostředí, které bylo na formaci přítomno, zahrnovalo potoky a říční kanály, mělčiny a mělká jezera. Velká část sedimentace také naznačuje, že existovalo bohaté stanoviště, které nabízí rozsáhlou vegetaci v hojném množství, které by dokázalo udržet většinu býložravých dinosaurů. Většina říčních systémů fungovala jako oázy pro oviraptorosaurs.[23][24][25] Další dinosauři v tomto formace patří ornithomimosaurs Anserimimus, Gallimimus, a taxon, který zůstává nepojmenovaný;[26] různé oviraptorosaury jako např Nemegtomaia, Elmisaurus a Gobiraptor;[24][23] a troodontid Zanabazar.[27] Velký dinosauři v této formaci jsou zastoupeni Deinocheirus, Saurolophus, Tarbosaurus a Therizinosaurus.[28][29]
Viz také
Reference
- ^ Barsbold, R. (1977). „Эволюции Хищных Динозавров“ [Vývoj dravých dinosaurů]. Transakce společné sovětsko-mongolské paleontologické expedice (v Rusku). 4: 48−56. Přeložený papír
- ^ A b C Barsbold, R. (1983). „Хищные динозавры мела Монголии“ [Masožraví dinosauři z křídy Mongolska] (PDF). Transakce společné sovětsko-mongolské paleontologické expedice (v Rusku). 19: 89. Přeložený papír
- ^ Senter, P. (2010). „Použití vědy o stvoření k prokázání evoluce: Aplikace kreacionistické metody pro vizualizaci mezer ve fosilním záznamu na fylogenetickou studii coelurosaurianských dinosaurů“. Journal of Evolutionary Biology. 23 (8): 1732−1743. doi:10.1111 / j.1420-9101.2010.02039.x. PMID 20561133.
- ^ A b C d E F G h i j k Turner, A. H .; Makovicky, P. J .; Norell, M. A. (2012). „Recenze systematiky dromaeosauridů a paravianské fylogeneze“. Bulletin of the American Museum of Natural History. 371 (371): 19−22. doi:10.1206/748.1. hdl:2246/6352. S2CID 83572446.
- ^ A b C d E F G h Brusatte, S.L .; Vremir, M .; Csiki-Sava, Z .; Turner, A. H .; Watanabe, A .; Erickson, G. M .; Norell, M. A. (2013). "Osteologie Balaur bondoc, ostrovního obydlí dromaeosaurida (Dinosauria, Theropoda) z pozdní křídy Rumunska". Bulletin of the American Museum of Natural History (374): 1−100. hdl:2246/6419.
- ^ A b Bishop, P. J. (2019). „Testování funkce„ srpkovitých drápů “dromaeosauridů (Dinosauria, Theropoda) prostřednictvím muskuloskeletálního modelování a optimalizace“. PeerJ. 7: e7577. doi:10,7717 / peerj.7577. PMC 6717499. PMID 31523517.
- ^ A b C Kubota, K .; Barsbold, R. (2006). „Reexamination of Adasaurus mongoliensis (Dinosauria: Theropoda) from the Upper Cretaceous Nemegt Formation of Mongolia“. Časopis paleontologie obratlovců. 26 (doplněk 03): 88A. doi:10.1080/02724634.2006.10010069.
- ^ Currie, P. J .; Varricchio, D. J. (2004). "New dromaeosaurid from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta, Canada". Feathered Dragons. Indiana University Press. str. 128. ISBN 0-253-34373-9. Náhled knihy
- ^ Kubota, K .; Barsbold, R. (2007). „New dromaeosaurid (Dinosauria Theropoda) from the Upper Cretaceous Bayanshiree Formation of Mongolia“. Časopis paleontologie obratlovců. 27 (doplň. do 3): 102A. doi:10.1080/02724634.2007.10010458.
- ^ Turner, A. H .; Pol, D .; Clarke, J. A .; Erickson, G. M .; Norell, M. A. (2007). „Bazální dromaeosaurid a vývoj velikosti před letem ptáků“. Věda. 317 (5843): 1378−1381. Bibcode:2007Sci ... 317.1378T. doi:10.1126 / science.1144066. PMID 17823350.
- ^ Kim, H. M .; Gishlick, A. D .; Tsuihiji, T. (2005). „První jiné než ptačí kosterní pozůstatky maniraptoranu ze spodní křídy v Koreji“. Křídový výzkum. 26 (2): 299−306. doi:10.1016 / j.cretres.2005.01.001. OCLC 717973416.
- ^ A b Perle, A .; Norell, M. A .; Clark, J. M. (1999). „Nový maniraptoran Theropod - Achillobator giganticus (Dromaeosauridae) - z horní křídy Burkhantu v Mongolsku“. Příspěvky předsedy geologie a mineralogie, Mongolského národního muzea (101): 1−105. OCLC 69865262.
- ^ A b Norell, M. A .; Makovicky, P. J. (1997). „Důležité vlastnosti kostry dromaeosaurů: informace z nového vzorku“. Americké muzeum Novitates (3215): 1−28. hdl:2246/3557.
- ^ Sereno, P. C. (1998). „Odůvodnění fylogenetických definic s aplikací na taxonomii vyšší úrovně Dinosaurie“. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 210 (1): 41–83. doi:10.1127 / njgpa / 210/1998/41.
- ^ Longrich, N.R .; Currie, P. J. (2009). „Microraptorine (Dinosauria – Dromaeosauridae) z pozdní křídy Severní Ameriky“. Sborník Národní akademie věd. 106 (13): 5002−5007. doi:10.1073 / pnas.0811664106. PMC 2664043. PMID 19289829.
- ^ Makovicky, P. J .; Apesteguía, S .; Agnolín, F. L. (2005). „Nejstarší dromaeosaurid theropod z Jižní Ameriky“. Příroda. 437 (7061): 1007–1011. Bibcode:2005 Natur.437.1007M. doi:10.1038 / nature03996. PMID 16222297. S2CID 27078534.
- ^ Senter, P .; Kirkland, J. I .; Deblieux, D. D .; Madsen, S .; Toth, N. (2012). „New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail“. PLOS ONE. 7 (5): e36790. doi:10.1371 / journal.pone.0036790. PMC 3352940. PMID 22615813.
- ^ Cau, A .; Beyrand, V .; Voeten, D .; Fernandez, V .; Tafforeau, P .; Stein, K .; Barsbold, R .; Tsogtbaatar, K .; Currie, P .; Godrfroit, P. (2017). „Synchrotronové skenování odhaluje obojživelnou ekomorfologii v nové kladu ptačích dinosaurů“. Příroda. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. doi:10.1038 / příroda24679. PMID 29211712. S2CID 4471941.
- ^ Hartman, S .; Mortimer, M .; Wahl, W. R .; Lomax, D. R .; Lippincott, J .; Lovelace, D. M. (2019). „Nový paravianský dinosaurus z Late Jurassic v Severní Americe podporuje pozdní akvizici ptačího letu“. PeerJ. 7: e7247. doi:10,7717 / peerj.7247. PMC 6626525. PMID 31333906.
- ^ Powers, M. A .; Sullivan, C .; Currie, P. J. (2020). „Přezkoumání premaxilárních a maxilárních znaků založených na poměru v Eudromaeosauria (Dinosauria: Theropoda): Odlišné trendy morfologie čenichu mezi asijskými a severoamerickými taxony“. Paleogeografie, paleoklimatologie, paleoekologie. 547 (109704): 109704. Bibcode:2020PPP ... 547j9704P. doi:10.1016 / j.palaeo.2020.109704.
- ^ Norell, M. A .; Makovicky, P.J. (2004). "Dromaeosauridae". Ve Weishampel, D.B .; Dodson, P .; Osmólska, H. (eds.). Dinosaurie (2. vyd.). Berkley: University of California Press. 196-209. ISBN 9780520941434.
- ^ Fowler, D. W .; Freedman, E. A .; Scannella, J. B .; Kambic, R. E. (2011). „Dravá ekologie Deinonychus a původ mávání ptáky“. PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO ... 628964F. doi:10,1371 / journal.pone.0028964. PMC 3237572. PMID 22194962.
- ^ A b Funston, G. F .; Mendonca, S.E .; Currie, P. J .; Barsbold, R .; Barsbold, R. (2018). "Oviraptorosaur anatomie, rozmanitost a ekologie v Němegt Basin". Paleogeografie, paleoklimatologie, paleoekologie. 494: 101–120. doi:10.1016 / j.palaeo.2017.10.023.
- ^ A b Lee, S .; Lee, Y. N .; Chinsamy, A .; Lü, J .; Barsbold, R .; Tsogtbaatar, K. (2019). „Nový malý dinosaurus oviraptorid (Dinosauria: Theropoda) z formace Mongolska z horní křídy Němegt“. PLOS ONE. 14 (2): e0210867. doi:10.1371 / journal.pone.0210867. PMC 6364893. PMID 30726228.
- ^ Owocki, K .; Kremer, B .; Cotte, M .; Bocherens, H. (2020). „Dietní preference a klima odvozené z izotopů kyslíku a uhlíku zubní skloviny Tarbosaurus bataar (formace Nemegt, svrchní křída, Mongolsko)“. Paleogeografie, paleoklimatologie, paleoekologie. 537: 109190. doi:10.1016 / j.palaeo.2019.05.012.
- ^ Chinzorig, T .; Kobayashi, Y .; Tsogtbaatar, K .; Currie, P. J .; Takasaki, R .; Tanaka, T .; Iijima, M .; Barsbold, R. (2018). „Ornithomimosaurs from the Nemegt Formation of Mongolia: manus morpholo variation andiversity“. Paleogeografie, paleoklimatologie, paleoekologie. 494: 91–100. doi:10.1016 / j.palaeo.2017.10.031.
- ^ Norell, M. A .; Makovicky, P. J .; Bever, G. S .; Balanoff, A. M .; Clark, J. M .; Barsbold, R .; Rowe, T. (2009). "Recenze mongolských křídových dinosaurů Saurornithoides (Troodontidae: Theropoda)". Americké muzeum Novitates. 3654: 1–63. doi:10.1206/648.1. hdl:2246/5973.
- ^ Holtz, T. R. (2014). „Tajemství hrozných rukou vyřešeno“. Příroda. 515 (7526): 203−205. Bibcode:2014 Natur.515..203H. doi:10.1038 / příroda13930. PMID 25337885.
- ^ Fanti, F .; Bell, P. R .; Currie, P. J .; Tsogtbataar, K. (2020). „Němegt Basin - jedna z nejlepších polních laboratoří pro interpretaci pozdně křídových suchozemských ekosystémů“. Paleogeografie, paleoklimatologie, paleoekologie. 494: 1–4. doi:10.1016 / j.palaeo.2017.07.014.

















{kind=link}