Megafauna - Megafauna

Pozemní zoologie, megafauna (z řecký μέγας megas "velké" a Nová latina fauna „zvířecí život“) zahrnuje velká nebo obří zvířata oblasti, stanoviště nebo geologického období. Nejběžnější používané prahové hodnoty jsou hmotnost přes 40 kilogramů (90 lb)[1] nebo 44 kilogramů (100 lb)[2][3] (tj. s hmotností srovnatelnou nebo větší než a člověk ) nebo nad a tuna, 1 000 kilogramů[1][4][5] (tj. s hmotností srovnatelnou nebo větší než vůl ). První z nich zahrnuje mnoho druhů, o nichž se běžně nepíše, že jsou příliš velké, jako např jelenec běloocasý a červený klokan.
V praxi nejčastější použití, se kterým se setkáváme v akademickém a populárním psaní, popisuje suchozemské savce zhruba větší než člověk, kteří nejsou (pouze) domestikováni. Termín je zvláště spojován s Pleistocene megafauna - suchozemská zvířata často větší než moderní protějšky považované za archetypické poslední doba ledová, jako mamuti, z nichž většina je v severní Eurasii, Americe a Austrálie vyhynuli za posledních čtyřicet tisíc let.[6] Mezi živými zvířaty je termín megafauna nejčastěji používán pro největší existující suchozemští savci, kterými jsou sloni, žirafy, hrochy, nosorožci a velké skot. Pouze z těchto pěti kategorií velkých býložravců skot se v současné době nacházejí mimo Afrika a jižní Asie, ale všechny ostatní byly dříve rozsáhlejší. Druhy Megafaunal lze rozdělit do kategorií podle jejich stravovacího typu: megaherbivores (např. sloni ), megacarnivores (např. lvi ) a vzácněji megaomnivory (např. medvědi ).
Jiná běžná použití jsou zejména pro obří vodní druhy velryby, jakékoli z větších divokých nebo domestikovaných suchozemských zvířat, jako jsou větší antilopa a dobytek, stejně jako dinosauři a další vyhynulé obří plazy.
Termín megafauna se velmi zřídka používá k popisu bezobratlých, ačkoli se občas používal pro některé druhy vyhynulých bezobratlých, které byly mnohem větší než všechny dnes žijící podobné druhy bezobratlých, například 1 m (3 ft) vážky z Karbon doba.
Ekologická strategie
Megafauna - ve smyslu největších savců a ptáků - je obecně K-stratégové s vysokou dlouhověkostí, pomalým tempem růstu populace, nízkou úmrtností a (alespoň u největších) několika nebo žádnými přirozenými predátory schopnými zabíjet dospělé.[7] Tyto vlastnosti, i když nejsou výlučné pro takovou megafaunu, je činí zranitelnými vůči člověku nadměrné vykořisťování, zčásti kvůli jejich pomalé obnově populace.[8][9]
Vývoj velké velikosti těla
Jedno pozorování, které bylo učiněno o vývoji větší velikosti těla, spočívá v tom, že rychlé rychlosti růstu, které jsou často pozorovány v relativně krátkých časových intervalech, nejsou udržitelné po mnohem delší časová období. Při zkoumání změn tělesné hmotnosti savců v průběhu času bylo zjištěno, že maximální možné zvýšení v daném časovém intervalu je v měřítku s délkou intervalu zvýšenou na 0,25 síly.[10] Předpokládá se, že to odráží vznik, během trendu zvyšování maximální velikosti těla, řady anatomických, fyziologických, environmentálních, genetických a dalších omezení, která musí být překonána evolučními inovacemi, než bude možné další zvětšení velikosti. Bylo zjištěno nápadně rychlejší tempo změn u velkého úbytku tělesné hmotnosti, který může být spojen s fenoménem ostrovní nanismus. Při normalizaci na generační délku bylo zjištěno, že maximální rychlost snížení tělesné hmotnosti je více než 30krát větší než maximální rychlost zvýšení tělesné hmotnosti pro desetinásobnou změnu.[10]
U suchozemských savců

Následně po Událost vyhynutí křída – paleogen který eliminoval jiné než ptačí dinosaury asi 66 Ma (před miliony let) zemští savci prošli téměř exponenciálním zvětšením velikosti těla, když se diverzifikovali, aby obsadili ekologické mezery, které zůstaly prázdné.[11] Počínaje pouhými několika kg před událostí dosáhla maximální velikost ~ 50 kg o několik milionů let později a ~ 750 kg do konce roku Paleocen. Zdá se, že se tento trend zvyšování tělesné hmotnosti ustálil asi před 40 Ma (pozdě Eocen ), což naznačuje, že bylo dosaženo fyziologických nebo ekologických omezení po zvýšení tělesné hmotnosti o více než tři řády.[11] Když se však vezme v úvahu z hlediska rychlosti nárůstu velikosti na generaci, je zjištěno, že exponenciální nárůst pokračuje, dokud se neobjeví Indricotherium Před 30 Ma. (Vzhledem k tomu, že se generuje časové měřítko s tělesná hmotnost0.259, zvyšující se časy generování se zvětšující se velikostí způsobí, že křivka logaritmu vs. času se zakřiví dolů z lineárního uložení.[10]
Megaherbivores nakonec dosáhli tělesné hmotnosti přes 10 000 kg. Největší z nich, indricotheres a proboscidy, byly fermentory zadního střeva, o nichž se věří, že mají výhodu nad fermentory předního střeva pokud jde o schopnost zrychlit průchod trávicím traktem, aby bylo možné pojmout velmi velký příjem potravy.[12] Podobný trend se objevuje, když u různých savců dochází k nárůstu maximální tělesné hmotnosti za generaci klady jsou porovnávány (pomocí průměrovaných sazeb za makroevoluční časové stupnice). Mezi suchozemskými savci je nejvyšší rychlost nárůstu o tělesná hmotnost0.259 vs. čas (v Ma) nastal v perissodaktyly (sklon 2,1), následovaný hlodavci (1.2) a proboscidy (1.1),[10] všechny jsou fermentory zadního střeva. Míra růstu pro artiodaktyly (0,74) byla asi třetina ve srovnání s perissodaktyly. Míra pro šelmy (0,65) byla zatím o něco nižší primáti, možná omezen jejich stromový návyky, měly nejnižší míru (0,39) ze studovaných skupin savců.[10]
Pozemní savčí šelmy z několika eutherian skupiny ( artiodactyl Andrewsarchus - dříve považován za mesonychid, oxyaenid Sarkastodon a šelmy Amphicyon a Arctodus ) všichni dosáhli maximální velikosti asi 1 000 kg[11] (šelma Arctotherium a hyaenodontid Simbakubwa může být o něco větší). Největší známý metatherian masožravec, Proborhyaena gigantea, zjevně dosáhl 600 kg, rovněž blízko této hranice.[13] Podobná teoretická maximální velikost pro savčí masožravce byla předpovězena na základě rychlosti metabolismu savců, energetických nákladů na získání kořisti a maximálního odhadovaného koeficientu rychlosti příjmu kořisti.[14] Rovněž bylo navrženo, že maximální velikost pro savčí masožravce je omezena stresem humerus vydrží při nejvyšší rychlosti jízdy.[13]
Analýza variací maximální velikosti těla za posledních 40 Ma naznačuje, že se snižováním teploty a zvětšováním kontinentální oblasti pevniny je spojena se zvyšováním maximální velikosti těla. Dřívější korelace by byla v souladu s Bergmannova vláda,[15] a může souviset s termoregulační výhoda velké tělesné hmotnosti v chladném podnebí,[11] lepší schopnost větších organismů vyrovnat se se sezónností v zásobování potravinami,[15] nebo jiné faktory;[15] druhou korelaci lze vysvětlit z hlediska rozsahu a omezení zdrojů.[11] Oba parametry však spolu souvisejí (kvůli poklesům hladiny moře doprovázeným zvýšeným zaledněním), což ztěžuje identifikaci hnacích sil trendů maximální velikosti.[11]
U mořských savců

Vzhledem k tomu, tetrapods (první plazi, později savci ) se vrátili do moře v pozdním permu, ovládli horní konec rozsahu velikosti mořského těla díky efektivnějšímu příjmu kyslíku pomocí plic.[16][17] Předci kytovci jsou považovány za semiaquatické pakicetidy, ne větší než psi, asi před 53 miliony let (Ma).[18] Před 40 Ma dosáhli kytovci délky 20 m nebo více Basilosaurus, podlouhlá hadovitá velryba, která se v mnoha ohledech odlišovala od moderních velryb a nebyla pro ně předky. V návaznosti na to se zdá, že vývoj velké velikosti těla u kytovců se dočasně zastavil a poté ustoupil, i když dostupné fosilní záznamy jsou omezené. V období před 31 Ma (v Oligocen ) až do současnosti došlo u kytovců k podstatně rychlejšímu trvalému přírůstku tělesné hmotnosti (tempo přírůstku) tělesná hmotnost0.259 faktorem 3,2 na milion let), než kterého dosáhla jakákoli skupina suchozemských savců.[10] Tento trend vedl k největšímu zvířeti všech dob, modernímu modrá velryba. Existuje několik důvodů pro rychlejší vývoj velké velikosti těla u kytovců. Méně biomechanické omezení nárůstu velikosti těla mohou být spojena se suspendováním ve vodě, na rozdíl od postavení proti gravitační síle, as plavecké pohyby naproti tomu pozemská lokomoce. Také větší tepelná kapacita a tepelná vodivost vody ve srovnání se vzduchem může zvýšit termoregulační výhoda velké velikosti těla v námořní dopravě endotermy, ačkoli platí klesající výnosy.[10]
U ozubených velryb se zdá, že maximální velikost těla je omezena dostupností potravy. Větší velikost, jako v spermie a zobákovitý velryby, usnadňuje hlubší potápění s přístupem k relativně snadno ulovené velké kořisti hlavonožců v méně konkurenčním prostředí. Ve srovnání s odontocetami je účinnost baleen velryb krmení filtru příznivěji se zvyšuje s rostoucí velikostí, když je planktonové jídlo husté, takže je větší velikost výhodnější. The výpad krmení technika rorquals se zdá být energeticky účinnější než krmení berana z balaenid velryby; druhá technika se používá u méně hustého a nerovnoměrného planktonu.[19] Trend ochlazování v nedávné historii Země mohl generovat více lokalit s vysokým výskytem planktonu pomocí větru upwellings, což usnadňuje vývoj gigantických velryb.[19]
Kytovci nejsou jedinými mořskými savci, kteří dosáhli ohromné velikosti. Největší šelmy všech dob jsou námořní ploutvonožci, z nichž největší je tuleň jižní, který může dosáhnout délky 6 metrů a vážit až 5 000 kilogramů (11 000 lb). Mezi další velké ploutvonožce patří tuleň severní při 4 000 kilogramech (8 800 lb), mrož při 2 000 kilogramech (4 400 lb) a Steller lachtan při 1135 kilogramech (2502 lb). The sireniani jsou další skupinou mořských savců, kteří se adaptovali na plně vodní život přibližně ve stejné době jako kytovci. Sirenians jsou úzce spjaty se slony. Největší sirenian byl Stellerova mořská kráva, který dosahoval délky až 10 metrů a vážil 8 000 až 10 000 kilogramů (18 000 až 22 000 lb) a byl loven k vyhynutí v 18. století. Semi-vodní hroch, který je suchozemským savcem nejblíže příbuzným kytovcům, může dosáhnout 3 200 kilogramů (7 100 lb).
U nelétavých ptáků

Vzhledem k malé počáteční velikosti všech savců po vyhynutí nelidských dinosaurů měli bezobratlí obratlovci zhruba deset milionů let dlouhou příležitost (během paleocénu) pro vývoj gigantismu bez velké konkurence.[20] Během tohoto intervalu vrcholový predátor výklenky byly často obsazeny plazy, například pozemskými krokodýli (např. Pristichampsus ), velcí hadi (např. Titanoboa ) nebo varanidové ještěrky, nebo nelétavými ptáky[11] (např. Paleopsilopterus v Jižní americe). To je také období, kdy megafaunal nelétavý býložravý gastornithid ptáci se vyvinuli na severní polokouli, zatímco nelétali paleognáti se vyvinul do velké velikosti dne Gondwanan zemské masy a Evropa. Gastornithids a alespoň jedna linie nelétavých paleognathských ptáků pocházela z Evropy, obě linie dominovaly ve výklencích velkých býložravců, zatímco savci zůstali pod 45 kg (na rozdíl od jiných suchozemských mas jako Severní Amerika a Asie, který viděl dřívější vývoj větších savců) a byli největšími evropskými tetrapody v Paleocen.[21]
Pilotní paleognáti bez letu, nazývané ptáci nadřádu běžci, byly tradičně považovány za představující linii oddělenou od linie jejich malých příbuzných na útěku, Neotropické tinamous. Nicméně, nedávné genetické studie zjistily, že tinamous hnízdí dobře uvnitř stromu běžců a jsou sesterská skupina zaniklý moa Nového Zélandu.[20][22][23] Podobně malé kiwi Nového Zélandu bylo zjištěno, že je sesterskou skupinou vyhynulých sloní ptáci Madagaskaru.[20] Tato zjištění tomu nasvědčují nelétost a gigantismus vznikl nezávisle několikrát mezi ptáky nadřádu běžci prostřednictvím paralelní vývoj.
Draví megafaunální nelétaví ptáci byli často schopni konkurovat savcům na počátku Kenozoikum. Později v kenozoiku však byli vysídleni pokročilými šelmy a vymřeli. V severní Americe bathornithids Paracrax a Bathornis byli vrcholní dravci, ale vyhynuli Raný miocén. V Jižní Americe související phorusrhacids sdílel dominantní dravé výklenky s metatherianem sparassodonts během většiny kenozoika, ale klesal a nakonec vyhynul poté, co eutherští predátoři dorazili ze Severní Ameriky (jako součást Skvělá americká výměna ) Během Pliocén. Naproti tomu velké býložravé ptáky nadřádu běžci přežily až do současnosti.
Žádný z nelétavých ptáků kenozoika, včetně dravců Brontornis, možná všežravý Dromornis[24] nebo býložravé Vorombe, kdy dorostl do hmotnosti mnohem vyšší než 500 kg, a tak nikdy nedosáhl velikosti největších savčích masožravců, natož největších savčích býložravců. Bylo navrženo, že rostoucí tloušťka ptačích skořápek v poměru k vaječné hmotě se zvyšující se velikostí vajec stanoví horní hranici velikosti ptáků.[25][poznámka 1] Největší druh Dromornis, D. stirtoni, možná vyhynul poté, co dosáhl maximální ptačí tělesné hmotnosti a poté byl vačnatcem překonán diprotodonty které se vyvinuly na několikanásobně větší velikosti.[28]
V obřích želvách
Obří želvy byly důležitými složkami pozdě Kenozoikum megafauny, přítomné na každém nepolárním kontinentu až do příjezdu homininans.[29][30] Největší známá suchozemská želva byla Atlas Megalochelys, zvíře, které pravděpodobně vážilo asi 1 000 kg.
Některé dřívější vodní testudiny, např. mariňák Archelon křídy a sladké vody Stupendemys miocénu, byly podstatně větší a váží více než 2 000 kg.
Masové vyhynutí megafaunalů
Načasování a možné příčiny
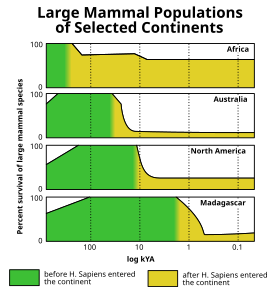
The Vyhynutí holocénu (viz také Kvartérní událost zániku ), došlo na konci poslední ledová doba ledová (aka Würmovo zalednění ) když mnoho obřích ledových savců, jako např vlněné mamuty, vyhynul v Amerika a severní Eurasie. Analýza události vyhynutí v Severní Americe zjistila, že je jedinečná mezi kenozoickými extinkčními impulsy v selektivitě pro velká zvířata.[31](Obr) Vlny vyhynutí přisuzovaly různé teorie lidský lov, klimatická změna, choroba, a domnělý mimozemský dopad nebo z jiných příčin. Toto vyhynutí se však blížilo ke konci Pleistocén byl jen jedním z řady megafaunálních extinkčních impulsů, ke kterým došlo během posledních 50 000 let na velké části zemského povrchu, s Afrika a jižní Asie (kde měla místní megafauna šanci vyvinout se spolu s moderními lidmi) byla ovlivněna relativně méně. Druhé oblasti trpěly postupným úbytkem megafauny, zejména pomaleji se pohybujících druhů (třída zranitelné megafauny ztělesněná obří želvy ), za posledních několik milionů let.[32][33]
Mimo pevninu Afro-Eurasie, tato megafaunální vyhynutí následovala vysoce charakteristický vzorec zemské masy, který úzce odpovídá šíření lidí do dříve neobydlených oblastí světa a který nevykazuje žádnou celkovou korelaci s klimatickou historií (kterou lze vizualizovat pomocí pozemků v posledních geologických časových obdobích) klimatických značek, jako je mořské izotopy kyslíku nebo atmosférické hladiny oxidu uhličitého ).[34][35] Austrálie[36] a blízké ostrovy (např. Flores[37]) byly zasaženy jako první zhruba před 46 000 lety, poté následovaly Tasmánie asi před 41 000 lety (po vytvoření pozemního mostu do Austrálie asi před 43 000 lety),[38][39][40] Japonsko zjevně asi před 30 000 lety,[41] Severní Amerika Před 13 000 lety,[poznámka 2] Jižní Amerika asi o 500 let později,[43][44] Kypr Před 10 000 lety,[45][46] the Antily Před 6000 lety[47][48] Nová Kaledonie[49] a blízké ostrovy[50] Před 3000 lety Madagaskar Před 2000 lety[51] Nový Zéland Před 700 lety[52] the Mascarenes Před 400 lety[53] a Velitelské ostrovy Před 250 lety.[54] Téměř všechny izolované ostrovy na světě by mohly poskytnout podobné příklady vyhynutí, ke kterým došlo krátce po příjezdu lidé, ačkoli většina z těchto ostrovů, jako je Havajské ostrovy, nikdy neměl pozemskou megafaunu, takže jejich vyhynulá fauna byly menší.[34][35]
Analýza načasování Holarctic megafaunální vymírání a vyhlazování za posledních 56 000 let odhalilo tendenci k tomu, aby se takové události shlukovaly mezistadiály období prudkého oteplování, ale pouze za přítomnosti lidí. Lidé mohli bránit procesům migrace a rekolonizace, které by jinak umožnily megafaunálním druhům přizpůsobit se změně klimatu.[55] Alespoň v některých oblastech byly interštadiály obdobím rozšiřování lidské populace.[56]
Analýza Sporormiella houbové spory (které pocházejí hlavně z hnoje megaherbivorů) v bažinatých sedimentárních jádrech překlenujících posledních 130 000 let od Lynchův kráter v Queensland Austrálie ukázala, že megafauna tohoto regionu prakticky zmizela asi před 41 000 lety, v době, kdy klimatické změny byly minimální; změna byla doprovázena nárůstem dřevěného uhlí a po ní následoval přechod z deštného pralesa na oheň odolný sklerofyl vegetace. Chronologie změn s vysokým rozlišením podporuje hypotézu, že samotný lov člověka eliminoval megafaunu a že následná změna flóry byla s největší pravděpodobností důsledkem eliminace prohlížečů a nárůstu požáru.[57][58][59][60] Nárůst ohně zpožďoval zmizení megafauny přibližně o sto let a nejpravděpodobněji byl důsledkem hromadění paliva, jakmile se procházení zastavilo. Během příštích několika staletí tráva rostla; vegetace sklerofylu rostla se zpožděním dalšího století a sklerofylový les se vyvinul asi po dalších tisících letech.[59] Během dvou období změny podnebí asi před 120 000 a 75 000 lety se na místě také zvýšila vegetace sklerofylů v reakci na přechod k chladnějším a suchším podmínkám; ani jedna z těchto epizod neměla významný dopad na hojnost megafaunů.[59] Podobné závěry týkající se zavinění lidských lovců zmizením pleistocénní megafauny byly odvozeny z chronologií s vysokým rozlišením získaných analýzou velké sbírky fragmentů skořápky nelétavého australského ptáka Genyornis newtoni,[61][62][60] z analýzy Sporormiella houbové spory z jezera ve východní části Severní Ameriky[63][64] a ze studia vkladů Shasta pozemní lenost trus odešel ve více než půl tuctu jeskyní na americkém jihozápadě.[65][66]
Pokračující lov lidí a narušení životního prostředí vedly k dalším megafaunální vyhynutí v nedávné minulosti, a vytvořil vážné nebezpečí dalších vyhynutí v blízké budoucnosti (viz příklady níže). Přímé zabíjení lidmi, zejména na maso, je nejdůležitějším faktorem současného úpadku megafaunů.[67][68]
Řada dalších hromadné vymírání došlo dříve v geologické historii Země, ve které také vymizela některá nebo celá megafauna té doby. Skvěle, v Událost vyhynutí křída – paleogen nelidské dinosaury a většina ostatních obřích plazů byly odstraněny. Dřívější masové vymírání však bylo globálnější a pro megafaunu nebylo tak selektivní; tj. mnoho druhů jiných typů, včetně rostlin, mořských bezobratlých[69] a plankton také vyhynuli. Dřívější události tedy musely být způsobeny obecnějšími typy narušení biosféra.
Důsledky vyčerpání megafauny
Vliv na transport živin
Megafauna hraje významnou roli v bočním transportu minerálních živin v ekosystému a má tendenci je přemisťovat z oblastí s vysokou mírou výskytu do oblastí s nízkým výskytem. Činí tak svým pohybem mezi časem, který živinu konzumují, a časem, kdy ji uvolní eliminací (nebo v mnohem menší míře rozkladem po smrti).[70] V Jižní Americe Amazonská pánev, odhaduje se, že taková boční difúze byla snížena o více než 98% po megafaunálních vyhynutích, ke kterým došlo zhruba před 12 500 lety.[71][72] Vzhledem k tomu fosfor Předpokládá se, že dostupnost omezuje produktivitu ve velké části regionu, pokles její dopravy ze západní části povodí a z niv (obě tyto zdroje jsou odvozeny od pozvednutí Andy ) do dalších oblastí se má za to, že významně ovlivnily ekologii regionu a účinky ještě nemusí dosáhnout svých limitů.[72] V moři se předpokládá, že kytovci a ploutvonožci, kteří se živí v hloubce, přemisťují dusík z hluboké do mělké vody, což zvyšuje oceánská produktivita a působí proti činnosti společnosti zooplankton, které mají tendenci dělat opak.[73]
Vliv na emise metanu
Velká populace megaherbivorů má potenciál významně přispět k atmosférické koncentraci metan, což je důležité skleníkový plyn. Moderní přežvýkavec býložravci vyrábět metan jako vedlejší produkt fermentace předního střeva při trávení a uvolněte jej říháním nebo nadýmáním. Dnes kolem 20% ročně emise metanu pocházejí z uvolňování metanu hospodářských zvířat. V Druhohor Odhaduje se, že sauropody mohl ročně vypustit do atmosféry 520 milionů tun metanu,[74] přispívá k teplejšímu podnebí času (až o 10 ° C teplejší než v současnosti).[74][75] Tato velká emise vyplývá z enormně odhadované biomasy sauropodů, a protože se předpokládá, že produkce metanu u jednotlivých býložravců je téměř úměrná jejich hmotnosti.[74]
Nedávné studie ukázaly, že vyhynutí megafaunálních býložravců mohlo způsobit snížení výskytu atmosférický metan. Tato hypotéza je relativně nová.[76] Jedna studie zkoumala emise metanu z bizon který obsadil Great Plains Severní Ameriky před kontaktem s evropskými osadníky. Studie odhaduje, že odstranění bizonů způsobilo pokles až o 2,2 milionu tun ročně.[77] Další studie zkoumala změnu koncentrace metanu v atmosféře na konci roku 2006 Pleistocene epocha po zániku megafauny v Severní a Jižní Americe. Poté, co raní lidé migrovali do Ameriky asi 13 000 BP, jejich lov a další související ekologické dopady vedly k vyhynutí mnoha druhů tamních megafaunalů. Výpočty naznačují, že tento zánik snížil produkci metanu přibližně o 9,6 milionu tun ročně. To naznačuje, že absence megafaunálních emisí metanu mohla přispět k náhlému klimatickému ochlazení na počátku Mladší Dryas.[76] Snížení atmosférického methanu, ke kterému došlo v té době, jak bylo zaznamenáno v ledová jádra, byl 2-4krát rychlejší než jakýkoli jiný pokles za poslední půl milionu let, což naznačuje, že fungoval neobvyklý mechanismus.[76]
Příklady
Následuje několik pozoruhodných příkladů zvířat, která jsou často považována za megafauny (ve smyslu definice „velkého zvířete“). Tento seznam nemá být vyčerpávající:
- Clade Synapsida
- Třída Mammalia (fylogeneticky, a clade v Therapsida; viz. níže)
- Infraclass Metatheria
- Objednat Diprotodontie
- The červený klokan (Macropus rufus) je největší žijící Australan savec a vačnatec při hmotnosti až 85 kg (187 lb). Nicméně, jeho zaniklý příbuzný, obří klokan s krátkou tváří Procoptodon goliah dosáhl 230 kg (510 lb),[78] zatímco zaniklý diprotodonty dosáhl největší velikosti ze všech vačnatců v historii, až do odhadovaných 2750 kg (6060 lb). Vyhynulý lev vačnatec (Thylacoleo carnifex ), až 160 kg (350 lb) byl mnohem větší než jakýkoli existující masožravý vačnatec.
- Objednat Diprotodontie
- Infraclass Eutheria
- Superřádek Afrotheria
- Objednat Proboscidea
- Sloni jsou největší žijící suchozemská zvířata. Oni a jejich příbuzní povstali v roce Afrika, ale donedávna měla téměř celosvětovou distribuci. The Slon africký (Loxodonta africana) má výšku ramene až 4,3 m (14 ft) a váží až 10,4 tuny (11,5 čistých tun).[79] Mezi nedávno vyhynulými proboscideany mamuti (Mammuthus) byli blízcí příbuzní slonů, zatímco mastodonti (Mammut) byly mnohem vzdáleněji příbuzné. The stepní mamut (M. trogontherii) se odhaduje, že běžně vážil kolem 10 tun, což z něj činí pravděpodobně největší proboscid, což by z něj učinilo druhého největšího suchozemského savce indricotheriny.
- Objednat Sirenia
- Největší sirenian s hmotností do 1 500 kg (3 300 lb) je Západoindický kapustňák (Trichechus manatus). Stellerova mořská kráva (Hydrodamalis gigas) byl pravděpodobně asi pětkrát tak obrovský, ale byl vyhuben lidmi do 27 let od jeho objevení mimo dálkový ovladač Velitelské ostrovy v prehistorických dobách tato mořská kráva žila také podél pobřeží severovýchodu Asie a severozápadní Severní Amerika; to bylo zjevně vyloučeno z těchto přístupnějších míst domorodými lovci.
- Objednat Proboscidea
- Superřádek Xenarthra
- Objednat Cingulata
- The glyptodonty byla skupina velkých, těžce obrněných ankylosaur -jako xenarthrans související s bydlením pásovci. Vznikly v Jižní Amerika, napadl Severní Ameriku během Skvělá americká výměna a vyhynuli na konci Pleistocén epocha.[80]
- Objednat Pilosa
- Broušení lenosti byly další skupinou pomalých, pozemských xenarthranů, příbuzných moderním lenost stromu. Měli podobnou historii, i když se dříve dostali do Severní Ameriky a rozšířili se dále na sever (např. Megalonyx ). Největší rody, Megatherium a Eremotherium, dosáhly velikostí srovnatelných se slony.[80]
- Objednat Cingulata
- Superřádek Euarchontoglires
- Objednat Primáti
- Největší žijící primát, s hmotností až 266 kg, je gorila (Gorilla beringei a Gorila gorila, přičemž tři ze čtyř poddruhů jsou kriticky ohrožený ). Zaniklý Madagaskar lenost lemur Archaeoindris dosáhl podobné velikosti, zatímco zaniklý Gigantopithecus blacki z Jihovýchodní Asie Předpokládá se, že byl ještě větší, i když pravděpodobně méně než dvakrát tak velký, na rozdíl od prvních odhadů (absence postkraniálních pozůstatků ztěžuje posouzení jeho velikosti).[81] Některé populace archaický Homo byly v průměru výrazně větší než v nedávné době Homo sapiens;[82][83] například, Homo heidelbergensis v jižní Africe mohla běžně dosáhnout výšky 2,1 m,[84] zatímco Neandertálci byly asi o 30% masivnější.[85]
- Objednat Rodentia
- Existující kapybara (Hydrochoerus hydrochaeris) Jižní Ameriky, největšího žijícího hlodavce, váží až 80 kg (180 lb).[86] Několik nedávno vyhynulých severoamerických forem bylo větších: kapybara Neochoerus pinckneyi (další Neotropické migrant) byl v průměru asi o 40% těžší; the obří bobr (Castoroides ohioensis) byl podobný. Zaniklý hutia s tupými zuby (Amblyrhiza inundata) z několika karibský ostrovy mohly být ještě větší. Před několika miliony let však Jižní Amerika chovala mnohem masivnější hlodavce. Phoberomys pattersoni, známý z téměř plné kostry, pravděpodobně dosáhl 700 kg (1 500 lb). To naznačují fragmentární pozůstatky Josephoartigasia monesi vzrostl na více než 1 000 kg (2 200 lb).
- Objednat Primáti
- Superřádek Laurasiatheria
- Objednat Carnivora
- Největší existující kočky jsou rodu Panthera, včetně tygr (P. tigris) a lev (P. leo).[87] The sibiřský tigr (P. t. altajka) by měla být největší divoká kočka podle Bergmannova vláda, a byl některými považován za takový[88][89] ale to je sporné.[90] Historicky se počet divokých sibiřských tygrů zmenšil a nyní jsou menší než bengálští tygři (P. t. tigris);[91] nicméně, sibiřští tygři stále inklinují být největší z tygrů v zajetí a dosahují hmotnosti asi 320 kg (710 lb).[92] Panthera druhy se vyznačují morfologické funkce, které jim umožňují řvát. Mezi větší vyhynulé kočky patří Americký lev (P. atrox) a jihoamerický šavlozubá kočka (Smilodon populator ).
- Medvědi jsou velcí šelmy z caniform podřád. Největší živé formy jsou lední medvěd (Ursus maritimus), s tělesnou hmotností až 800 kg (1 800 lb),[93] a téměř stejně velký Medvěd Kodiak (Ursus arctos middendorffi),[94] v souladu s Bergmannovým pravidlem. Arctotherium augustany, zaniklý medvěd krátký z Jižní Ameriky, byl největší dravý suchozemský savec, který kdy měl odhadovanou průměrnou hmotnost 1600 kg (3500 lb).[95]
- Těsnění, lachtani a mrože jsou obojživelní mořští šelmy, kteří se vyvinuli z medvědích předků. The tuleň jižní (Mirounga leonina) z antarktický a subantarctic Waters je největším masožravcem všech dob, přičemž býčí samci dosahují maximální délky 6–7 m (20–23 ft) a maximální hmotnosti 5 000 kg (11 000 lb).
- Objednat Perissodactyla
- Tapíry procházejí zvířata s krátkým prehensilním čenichem a podobou prasete, která se za 20 milionů let zdála jen málo změněna. Obývají tropické lesy jihovýchodní Asie a jižní a střední Ameriky a zahrnují největší přežívající suchozemská zvířata z posledních dvou regionů. Existují čtyři druhy.
- Nosorožci jsou lichokopytníci s rohy z keratin, stejného typu protein skládání vlasů. Patří mezi druhé největší žijící suchozemské savce s hmotností 850–3 800 kg. Tři z pěti existujících druhů jsou kriticky ohrožený. Jejich vyhynulý střední Asie příbuzní indricotheriny byli největší suchozemští savci všech dob.
 Nosorožec, od Dürerův dřevoryt
Nosorožec, od Dürerův dřevoryt
- Objednat Artiodactyla
- Žirafy (Giraffa spp.) jsou nejvyšší žijící suchozemská zvířata, dosahující výšky až téměř 6 m (20 ft). Průměrná hmotnost je 1 192 kg (2628 lb) pro dospělého muže a 828 kg (1825 lb) pro dospělou ženu s maximální hmotností 1,930 kg (4250 lb) a 1180 kg (2600 lb) zaznamenaných pro muže, respektive ženy.
- Skot kopytníci zahrnují největší přežívající suchozemská zvířata z Evropa a Severní Amerika. The vodní buvol (Bubalis arnee), bizon (Bison bison a B. bonasus ), a gaur (Bos gaurus) mohou všichni dorůst do hmotnosti přes 1000 kg (2200 lb).
- The semiaquatic hroch (Hippopotamus amphibius) je nejtěžší žijící člen řádu Cetartiodactyla po kytovcích. Průměrná hmotnost dospělých je kolem 1 500 kg (3300 lb) u mužů a 1300 kg (2900 lb) u mužů, respektive u žen, přičemž velcí muži dosahují přes 3 200 kg (7 100 lb). Hroch a mnohem menší kriticky ohrožený trpasličí hroch (Choeropsis liberiensis) jsou považovány za nejbližší existující příbuzní kytovců. Hroši patří mezi megafaunální druhy nejnebezpečnější lidem.[96]
- Infraorder Kytovci
- Velryby, delfíni a sviňuchy jsou mořští savci. The modrá velryba (Balaenoptera musculus) je největší baleen velryba a největší zvíře, jaké kdy žilo, 30 metrů (98 stop)[97] na délku a 170 tun (190 malých tun)[98] nebo více na váhu. The vorvaně (Physeter macrocephalus) je největší ozubená velryba a jeden z největších predátorů v historii obratlovců, stejně jako nejhlasitější a nejsilnější planeta nejrozumnější zvíře (s mozek asi pětkrát tak masivní jako a lidské ). The zabijácká velryba (Orcinus orca) je největší delfín.
- Objednat Carnivora
- Superřádek Afrotheria
- Infraclass Metatheria
- Objednat Pelycosauria (tradiční; paraphyletic )
- Cotylorhynchus byl velký, s drápy, býložravý ID případu brzy Permu Severní Amerika, dosahující 6 m (2 stopy) a 2 tuny.
- Objednat Therapsida
- Anteosaurus byl bití hlavy, semiaquatic, masožravý dinocephalian z Střední permu Jižní Afrika. Dosahoval délky 5–6 m (16–20 ft) a vážil asi 500–600 kg (1100–1300 lb).[99]
- Lisowicia byl býložravý o velikosti slona (9 tun) kannemeyeriiform dicynodont pozdně triasové Evropy.[100][101]
- Třída Mammalia (fylogeneticky, a clade v Therapsida; viz. níže)
- Clade Sauropsida
- Třída Aves (fylogeneticky, kladu uvnitř Coelurosauria, a taxon v rámci objednávky Saurischia; viz. níže)
- Objednat Struthioniformes
- The ptáci nadřádu běžci jsou starodávnou a různorodou skupinou nelétaví ptáci které se nacházejí na fragmentech prvního superkontinent Gondwana. Největší žijící pták, pštros (Struthio camelus) byl vyhynulý Vorombe z Madagaskar, nejtěžší ze skupiny až do (860 kg (1900 lb)), a zaniklý obří moa (Dinornis) z Nový Zéland, nejvyšší, dorůstající do výšek 3,4 m (11 ft). Poslední dva jsou příklady ostrovní gigantismus.
- Objednat Gastornithiformes
- Vyhynulý dromornithids Austrálie jako např Dromornis se přiblížil k největším běžcům ve velikosti. (Vzhledem ke své malé rozloze pro kontinent a jeho izolaci se na Austrálii někdy pohlíží jako na největší ostrov na světě; tyto druhy tedy lze také považovat za ostrovní obry.)
- Objednat Cathartiformes
- Zaniklý kondor -jako teratorn Argentavis Jižní Amerika měla odhadované rozpětí křídel 5 až 6 m (16 až 20 stop) a hmotnost přibližně 70 kg (150 lb), což z něj činí nejlepší příklad megafaunálního létajícího ptáka.
- Objednat Struthioniformes
- Třída Reptilia (tradiční; paraphyletic )
- Objednat Saurischia
- Saurischian dinosauři z jurský a Křídový zahrnout sauropody, nejdelší (až do 40 m nebo 130 stop) a nejmohutnější známá suchozemská zvířata (Argentinosaurus dosáhl 80–100metrických tun nebo 90–110tun ), stejně jako teropody, největší suchozemští masožravci (Spinosaurus vzrostl na 7–9 tun; slavnější Tyranosaurus, na 6,8 tuny).
- Objednat Pterosaurie
- Největší azhdarchid ptakoještěr, jako Hatzegopteryx a Quetzalcoatlus, dosáhl rozpětí křídel kolem 11–12 m (36–39 ft) a hmotnosti pravděpodobně v rozmezí 70–250 kg (150–550 lb). První z nich je považován za vrcholného dravce ostrovní ekosystém.
- Objednat Crocodilia
- Aligátoři a krokodýli jsou velcí semiaquatičtí plazi, z nichž největší je slané vody krokodýl (Crocodylus porosus), může dorůst do hmotnosti 1360 kg (3000 lb). Vzdálení předkové krokodýlů a jejich příbuzní, pseudosuchians (tradiční crurotarsans ), ovládl svět na konci roku Trias až do Událost zániku triasu – jury dovolil dinosaurům je předjet. Během později zůstali různorodí Druhohor, když krokodýli jako Deinosuchus a Sarkosuchus dosáhl délky 12 m. Podobně velcí krokodýli, jako např Mourasuchus a Purussaurus, byli přítomni teprve nedávno Miocén v Jižní americe.
- Objednat Squamata
- Zatímco největší existující ještěrka, varan komodský (Varanus komodoensis), další ostrovní gigant, může dosáhnout délky 3 m (10 ft), svého zaniklého australského příbuzného Megalania může dosáhnout více než dvojnásobku této velikosti. Tyto monitorovat ještěrky "mořští příbuzní, mosasaurs, byli vrcholní dravci v pozdních křídových mořích.
- Nejtěžší existující had je považován za zelená anakonda (Eunectes murinus), zatímco síťovaný python (Python reticulatus), až 8,7 m nebo více, je považován za nejdelší. Zaniklý Australan Pliocén druhy Liasis, Bluff Downs obří python, dosáhl 10 m, zatímco Paleocen Titanoboa Jižní Ameriky dosáhlo délky 12–15 m a odhadované hmotnosti asi 1 135 kilogramů (2 500 liber).
- Objednat Testudiny
- Největší želva je kriticky ohrožený námořní kožená želva (Dermochelys coriacea) o hmotnosti až 900 kg (2 000 lb). Liší se od ostatních mořské želvy jeho nedostatkem a kostnatý skořápka. Nejmocnější suchozemští cheloniáni jsou obří želvy z Galapágy (Chelonoidis nigra ) a Aldabra Atoll (Aldabrachelys gigantea ), až do 300 kg (660 lb). Tyto želvy jsou největšími, kteří přežili sortiment obřích druhů želv, které byly široce přítomny na kontinentálních pevninách[29][30] a další ostrovy[29] během pleistocénu.
- Objednat Saurischia
- Třída Aves (fylogeneticky, kladu uvnitř Coelurosauria, a taxon v rámci objednávky Saurischia; viz. níže)
- Třída Obojživelníci (v širokém, pravděpodobně parafyletickém smyslu)
- Objednat Temnospondyli (vztah k existující obojživelníci je nejasný)
- Permský temnospondyl Prionosuchus, největší známý obojživelník, dosáhl délky 9 m a byl vodním predátorem připomínajícím krokodýla. Po objevení skutečných krokodýlů temnospondyly jako např Koolasuchus (5 m dlouhý) křída ustoupila do oblasti Antarktidy, než vyhynula.
- Objednat Temnospondyli (vztah k existující obojživelníci je nejasný)
- Třída Actinopterygii
- Objednat Tetraodontiformes
- Největší existující kostnatá ryba je mořské mušle (Mola mola), jehož průměrná hmotnost dospělého je 1 000 kg (2200 lb). I když je fylogeneticky „kostnatá ryba“, její kostra je především chrupavka (který je lehčí než kost ). Má tělo ve tvaru disku a pohání se svým dlouhým, tenkým hřbetní a anální ploutve; živí se především Medúza. V těchto třech ohledech (stejně jako jeho velikost a potápěčské návyky) připomíná koženou želvu.
- Objednat Lampriformes
- The obří oarfish (Regalecus glesne) je nejdelší kostnatá ryba, která dosahuje 11 m (36 ft).
- Objednat Acipenseriformes
- The kriticky ohrožený beluga (Jeseter evropský, Huso huso) až do 1 476 kg (3 254 lb) je největší jeseter (které jsou také většinou chrupavčité) a je považován za největší anadromní Ryba.
- Objednat Siluriformes
- The kriticky ohrožený Obří sumec Mekong (Pangasianodon gigas), s hmotností až 293 kg (646 lb), je často považován za největší sladkovodní ryba.
- Objednat Tetraodontiformes
- Třída Chondrichthyes
- Objednat Lamniformes
- Největší žijící dravá ryba, velký bílý žralok (Carcharodon carcharias), reaches weights up to 2,240 kg (4,940 lb). Its extinct relative C. megalodon (the disputed genus being either Carcharodon nebo Carcharocles) was more than an řádově larger, and is the largest predatory shark or fish of all time (and one of the largest predators in vertebrate history); it preyed on whales and other mořští savci.
- Objednat Orectolobiformes
- The largest extant žralok, paryby, a Ryba overall is the žralok velrybí (Rhincodon typus), which reaches weights in excess of 21.5 tonnes (47,000 pounds). Like baleen whales, it is a podavač filtrů and primarily consumes plankton.
- Objednat Rajiformes
- Objednat Lamniformes
- Třída Placodermi
- Objednat Arthrodira
- The largest armored fish, Dunkleosteus, arose during the late Devonian. At up to 10 metres (33 ft) in length[102] and 3.6 tun (4.0 malé tuny ) in mass,[103] bylo to hyperkaravský vrcholový predátor that employed sání krmení.[104][105] Its contemporary, Titanichthys, apparently an early filter feeder, rivaled it in size. The arthrodires were eliminated by the environmental upheavals of the Pozdní devonský zánik, after existing for only about 50 million years.
- Objednat Arthrodira
- Třída Cephalopoda
- Objednat Ammonitida
- The Late Cretaceous amonit Parapuzosia seppenradensis reached a shell diameter of over 2 m.
- Objednat Teuthida
- A number of deep ocean creatures exhibit abyssal gigantism. Mezi ně patří obrovská oliheň (Architeuthis) a colossal squid (Mesonychoteuthis hamiltoni); both (although rarely seen) are believed to attain lengths of 12 m (39 ft) or more. The latter is the world's largest bezobratlý, and has the largest oči of any animal. Both are preyed upon by sperm whales.
- Objednat Ammonitida
- Stem-group Arthropoda
- Objednat Radiodonta
- Anomalocarids were a group of very early legless marine arthropods that included the largest predators of the Kambrijský, jako Anomalocaris. Brzy Ordovik they had evolved into giant (for the time) filter feeders, apparently in response to the proliferation of plankton during the Skvělá událost biodiverzifikace ordoviků. Aegirocassis grew to over 2 m in length.[106]
- Objednat Eurypterida
- Eurypterids (sea scorpions) were a diverse group of aquatic and possibly amphibious predators that included the most massive členovci to have existed. They survived over 200 million years, but finally died out in the Událost vyhynutí perm-trias spolu s trilobiti and most other forms of life present at the time, including most of the dominant terrestrial therapsids. Raný Devonský Jaekelopterus reached an estimated length of 2.5 m (8.2 ft), not including its raptorial chelicerae, and is thought to have been a freshwater species.
- Objednat Radiodonta

Galerie
Vyhynulý

Nějaký Paleozoikum sea scorpions (Eurypterus shown) were larger than a human.

Dunkleosteus was a 10 m (33 ft) long toothless obrněný predatory Devonian placoderm Ryba.

Sail-backed pelycosaur Dimetrodon a temnospondyl Eryops from North America's Permu.

Leedsichthys, a mid-jurský podavač filtrů fish, may have reached sizes of 7–16.5 m (23–54 ft).

Macronarian sauropody; from left, Camarasaurus, Brachiosaurus, Žirafa, Euhelopus.

Tyranosaurus was a 12.3 m (40 ft) long theropod dinosaur, an vrcholový predátor z west North America.

asijský indricothere nosorožec Paraceratherium was among the largest land mammals,[107] about twice a bush elephant's mass.

The Pozdní miocén teratorn Argentavis of South America had a 7 m (23 ft) wingspan.

Reconstructed jaws of C. megalodon (Baltimore ).

Deinotherium had downward-curving tusks and ranged widely over Afro-Eurasie.

Titanis walleri, jediný teroristický pták known to have napadl North America, was 2.5 m (8.2 ft) tall.

Hippo-sized Diprotodon of Australia, the largest vačnatec of all time, became extinct 40,000 years ago.

Glyptodon, from South America's Pleistocén, was an auto-sized cingulate, příbuzný pásovci.

Američtí lvi exceeded existující lvi in size and ranged over much of N. America until 11,000 BP.

Vlnití mamuti vanished after humans invaded their habitat in Eurasia and N. America.[1]

Haastův orel, the largest eagle known, attacking moa (which included the tallest bird known).


















Živobytí

The eastern gorilla is the largest and one of the more endangered primates na planetě.

Nejčastější tiger subspecies, Bengálští tygři jsou ohrožený podle poaching a ničení stanovišť.

Lední medvědi, among the largest medvědi (consistent with Bergmann's rule ), jsou zranitelný na globální oteplování.

The kriticky ohrožený černý nosorožec, up to 3.75 metres (12.3 ft) long, is vyhrožoval by poaching.

Wild Bactrian camels are critically endangered. Their ancestors originated in North America.

Unlike woolly nosorožci a mamuti, Pižma narrowly survived the Quaternary extinctions.[1]

Hippopotamuses, the heaviest and most aquatic sudokopytníci, jsou velryby ' closest living relatives.

The vorvaně, největší ozubená velryba and toothed dravec, má biggest brain.

The orca, největší delfín a zabalit dravce, je highly intelligent a žije v complex societies.

The kasowary, the heaviest non-African bird, can run at 50 km/h through dense deštný prales.

The slané vody krokodýl is the largest living plaz and a dangerous predator of humans.

The varan komodský, an insular giant a největší ještěrka, má zoubkovaný teeth and a venomous bite.

The zelená anakonda, vodní svěrač, is the heaviest had, weighing up to 97.5 kg (215 lb) or more.

The deep-diving mořské mušle je největší kostnatá ryba, but its skeleton is mostly chrupavčitý.

The Okoun nilský, one of the largest freshwater fish, is also a damaging invazivní druhy.[Poznámka 3]

The velký bílý, the largest macropredatory fish, is more endangered than the tygr.[110]

Examination of a 9 m obrovská oliheň, an abyssal giant and the second largest hlavonožce.


















{kind=link}
{kind=link}
Viz také
- Australská megafauna
- Bergmann's rule
- Charismatic megafauna
- Copeovo pravidlo
- Hlubinný gigantismus
- Fauna
- Ostrovní nanismus
- Ostrovní gigantismus
- Největší organismy
- Největší prehistorické organismy
- List of heaviest land mammals
- List of largest mammals
- List of megafauna discovered in modern times
- Megafauna (mythology)
- Megafaunal wolf
- Megaflora
- Megaherb
- New World Pleistocene extinctions
- Pleistocene megafauna
- Kvartérní událost zániku
Poznámky
- ^ Nonavian dinosaur size was not similarly constrained because they had a different relationship between body mass and egg size than birds. The 400 kg Aepyornis had larger eggs than nearly all dinosaurs.[26][27]
- ^ Analysis indicates that 35 genera of North American mammals went extinct more or less simultaneously in this event.[42]
- ^ Perspektivní makes the fish appear larger relative to the man standing behind it (another example of a megafaunal species) than it actually is.
Reference
- ^ A b C d Stuart, A. J. (November 1991). "Mammalian extinctions in the Late Pleistocene of northern Eurasia and North America". Biologické recenze. 66 (4): 453–562. doi:10.1111/j.1469-185X.1991.tb01149.x. PMID 1801948. S2CID 41295526.
- ^ Martin, P. S. (1984). "Prehistoric overkill: The global model". In Martin, P. S.; Klein, R. G. (eds.). Quaternary Extinctions: A Prehistoric Revolution. University of Arizona Press. pp. 354–403. ISBN 978-0-8165-1100-6. OCLC 258362030.
- ^ Martin, P. S.; Steadman, D. W. (1999-06-30). „Prehistorické vyhynutí na ostrovech a kontinentech“. V MacPhee, R. D. E (ed.). Vymírání v blízké době: příčiny, souvislosti a důsledky. Pokroky v paleobiologii obratlovců. 2. New York: Kluwer / Plenum. pp. 17–56. ISBN 978-0-306-46092-0. OCLC 41368299. Citováno 2011-08-23. see page 17
- ^ Richard A. Farina, Sergio F. Vizcaino, Gerry De Iuliis (2013). "The Great American Biotic Interchange". Megafauna: Obří bestie pleistocénu v Jižní Americe. Indiana University Press, Bloomington, Indiana. p. 150. ISBN 978-0-253-00230-3.CS1 maint: používá parametr autoři (odkaz)
- ^ Bernhard A. Huber, Bradley J. Sinclair, Karl-Heinz Lampe (2005). "Historical Determinants of Mammal Species in Africa". African Biodiversity: Molecules, Organisms, Ecosystems. Springer. p. 294. ISBN 978-0387243153.CS1 maint: používá parametr autoři (odkaz)
- ^ Ice Age Animals. Státní muzeum v Illinois
- ^ https://www.britannica.com/science/K-selected-species. Britannica. Retrieved 2017-4-2.
- ^ Barnosky, A. D. (2004-10-01). "Assessing the Causes of Late Pleistocene Extinctions on the Continents". Věda. 306 (5693): 70–75. Bibcode:2004Sci...306...70B. CiteSeerX 10.1.1.574.332. doi:10.1126/science.1101476. PMID 15459379. S2CID 36156087.
- ^ Brook, B. W .; Johnson, C. N. (2006). "Selective hunting of juveniles as a cause of the imperceptible overkill of the Australian Pleistocene megafauna". Alcheringa: Australasian Journal of paleontologie. 30 (sup1): 39–48. doi:10.1080/03115510609506854. S2CID 84205755.
- ^ A b C d E F G Evans, A. R.; Jones, D .; Boyer, A. G.; Brown, J. H .; Costa, D. P.; Ernest, S. K. M.; Fitzgerald, E. M. G.; Fortelius, M.; Gittleman, J.L .; Hamilton, M. J.; Harding, L. E.; Lintulaakso, K.; Lyons, S. K.; Okie, J. G.; Saarinen, J. J.; Sibly, R. M.; Smith, F. A.; Stephens, P. R.; Theodor, J. M.; Uhen, M. D. (2012-01-30). "The maximum rate of mammal evolution". PNAS. 109 (11): 4187–4190. Bibcode:2012PNAS..109.4187E. doi:10.1073/pnas.1120774109. PMC 3306709. PMID 22308461.
- ^ A b C d E F G Smith, F. A.; Boyer, A. G.; Brown, J. H .; Costa, D. P.; Dayan, T.; Ernest, S. K. M.; Evans, A. R.; Fortelius, M.; Gittleman, J.L .; Hamilton, M. J.; Harding, L. E.; Lintulaakso, K.; Lyons, S. K.; McCain, C.; Okie, J. G.; Saarinen, J. J.; Sibly, R. M.; Stephens, P. R.; Theodor, J.; Uhen, M. D. (2010-11-26). "The Evolution of Maximum Body Size of Terrestrial Mammals". Věda. 330 (6008): 1216–1219. Bibcode:2010Sci...330.1216S. CiteSeerX 10.1.1.383.8581. doi:10.1126/science.1194830. PMID 21109666. S2CID 17272200.
- ^ Clauss, M.; Frey, R.; Kiefer, B.; Lechner-Doll, M.; Loehlein, W.; Polster, C.; Roessner, G. E.; Streich, W. J. (2003-04-24). "The maximum attainable body size of herbivorous mammals: morphophysiological constraints on foregut, and adaptations of hindgut fermenters" (PDF). Ekologie. 136 (1): 14–27. Bibcode:2003Oecol.136...14C. doi:10.1007/s00442-003-1254-z. PMID 12712314. S2CID 206989975.
- ^ A b Sorkin, B. (2008-04-10). "A biomechanical constraint on body mass in terrestrial mammalian predators". Lethaia. 41 (4): 333–347. doi:10.1111/j.1502-3931.2007.00091.x.
- ^ Carbone, C.; Teacher, A; Rowcliffe, J. M. (2007-01-16). "The Costs of Carnivory". PLOS Biology. 5 (2, e22): 363–368. doi:10.1371/journal.pbio.0050022. PMC 1769424. PMID 17227145.
- ^ A b C Ashton, K. G.; Tracy, M. C.; de Queiroz, A. (October 2000). "Is Bergmann's Rule Valid for Mammals?". Americký přírodovědec. 156 (4): 390–415. doi:10.1086/303400. JSTOR 10.1086/303400. PMID 29592141.
- ^ Webb, J. (2015-02-19). "Evolution 'favours bigger sea creatures'". BBC novinky. BBC. Citováno 2015-02-22.
- ^ Heim, N. A.; Knope, M. L.; Schaal, E. K.; Wang, S. C.; Payne, J. L. (2015-02-20). "Cope's rule in the evolution of marine animals". Věda. 347 (6224): 867–870. Bibcode:2015Sci...347..867H. doi:10.1126/science.1260065. PMID 25700517.
- ^ Thewissen, J. G. M.; Bajpai, S. (1 January 2001). "Whale Origins as a Poster Child for Macroevolution". BioScience. 51 (12): 1037–1049. doi:10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2. ISSN 0006-3568.
- ^ A b Goldbogen, J. A .; Cade, D. E.; Wisniewska, D. M.; Potvin, J.; Segre, P. S.; Savoca, M. S.; Hazen, E. L.; Czapanskiy, M. F.; Kahane-Rapport, S. R.; DeRuiter, S. L.; Gero, S.; Tønnesen, P.; Gough, W. T.; Hanson, M. B.; Holt, M. M.; Jensen, F. H.; Simon, M.; Stimpert, A. K.; Arranz, P.; Johnston, D. W.; Nowacek, D. P.; Parks, S. E.; Visser, F.; Friedlaender, A. S.; Tyack, P. L.; Madsen, P. T.; Pyenson, N. D. (2019). "Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants". Věda. 366 (6471): 1367–1372. Bibcode:2019Sci...366.1367G. doi:10.1126/science.aax9044. hdl:10023/19285. PMID 31831666. S2CID 209339266.
- ^ A b C Mitchell, K. J .; Llamas, B .; Soubrier, J .; Rawlence, N.J .; Worthy, T. H .; Wood, J .; Lee, M. S. Y .; Cooper, A. (2014-05-23). "Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution" (PDF). Věda. 344 (6186): 898–900. Bibcode:2014Sci...344..898M. doi:10.1126/science.1251981. hdl:2328/35953. PMID 24855267. S2CID 206555952.
- ^ Buffetaut, E .; Angst, D. (listopad 2014). „Stratigrafická distribuce velkých nelétavých ptáků v paleogenu Evropy a její paleobiologické a paleogeografické důsledky“. Recenze vědy o Zemi. 138: 394–408. Bibcode:2014ESRv..138..394B. doi:10.1016/j.earscirev.2014.07.001.
- ^ Phillips MJ, Gibb GC, Crimp EA, Penny D (January 2010). "Tinamous and moa flock together: mitochondrial genome sequence analysis reveals independent losses of flight among ratites". Systematická biologie. 59 (1): 90–107. doi:10.1093/sysbio/syp079. PMID 20525622.
- ^ Baker, A. J.; Haddrath, O.; McPherson, J. D .; Cloutier, A. (2014). "Genomic Support for a Moa-Tinamou Clade and Adaptive Morphological Convergence in Flightless Ratites". Molekulární biologie a evoluce. 31 (7): 1686–1696. doi:10.1093/molbev/msu153. PMID 24825849.
- ^ Murray, Peter F.; Vickers-Rich, Patricia (2004). Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime. Indiana University Press. pp. 51, 314. ISBN 978-0-253-34282-9. Citováno 7. ledna 2012.
- ^ Ibid (2004). p. 212. ISBN 978-0253342829.
- ^ Kenneth Carpenter (1999). Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction. Indiana University Press. p.100. ISBN 978-0-253-33497-8. OCLC 42009424. Citováno 6. května 2013.
- ^ Jackson, F. D .; Varricchio, D. J .; Jackson, R. A .; Vila, B.; Chiappe, L. M. (2008). "Comparison of water vapor conductance in a titanosaur egg from the Upper Cretaceous of Argentina and a Megaloolithus siruguei egg from Spain". Paleobiologie. 34 (2): 229–246. doi:10.1666/0094-8373(2008)034[0229:COWVCI]2.0.CO;2. ISSN 0094-8373.
- ^ Ibid (2004). p. 277. ISBN 978-0253342829.
- ^ A b C Hansen, D. M .; Donlan, C. J .; Griffiths, C. J .; Campbell, K. J. (duben 2010). "Ecological history and latent conservation potential: large and giant tortoises as a model for taxon substitutions" (PDF). Ekografie. 33 (2): 272–284. doi:10.1111 / j.1600-0587.2010.06305.x. Archivovány od originál (PDF) 24. července 2011. Citováno 2011-02-26.
- ^ A b Cione, A. L .; Tonni, E. P .; Soibelzon, L. (2003). „The Broken Zig-Zag: Late Cenozoic large savec and tortoise extinction in South America“ (PDF). Mus. Argentino Cienc. Nat., N.s. 5 (1): 1–19. doi:10.22179 / REVMACN.5.26. ISSN 1514-5158. Archivovány od originál (PDF) 6. července 2011. Citováno 2011-02-06.
- ^ Alroy, J. (1999), "Putting North America's End-Pleistocene Megafaunal Extinction in Context: Large-Scale Analyses of Spatial Patterns, Extinction Rates, and Size Distributions", in MacPhee, R. D. E. (ed.), Extinctions in Near Time: Causes, Contexts, and Consequences, Advances in Vertebrate Paleobiology, 2, New York: Plenum, pp. 105–143, doi:10.1007/978-1-4757-5202-1_6, ISBN 978-1-4757-5202-1, OCLC 41368299
- ^ Corlett, R. T. (2006). "Megafaunal extinctions in tropical Asia" (PDF). Tropinet. 17 (3): 1–3. Citováno 2010-10-04.
- ^ Edmeades, Baz. "Megafauna — First Victims of the Human-Caused Extinction". megafauna.com. (internet-published book with Foreword by Paul S. Martin ). Archivovány od originál dne 2014-12-25. Citováno 2020-02-13.
- ^ A b Martin, P. S. (2005). "Chapter 6. Deadly Syncopation". Twilight of the Mammoths: Ice Age Extinctions and the Rewilding of America. University of California Press. pp. 118–128. ISBN 978-0-520-23141-2. OCLC 58055404. Citováno 2014-11-11.
- ^ A b Burney, D. A .; Flannery, T. F. (Červenec 2005). „Padesát tisíciletí katastrofických vyhynutí po kontaktu s lidmi“ (PDF). Trendy v ekologii a evoluci. 20 (7): 395–401. doi:10.1016 / j.tree.2005.04.022. PMID 16701402. Archivovány od originál (PDF) dne 10.06.2010. Citováno 2014-11-11.
- ^ Roberts, R. G .; Flannery, T. F.; Ayliffe, L. K.; Yoshida, H .; Olley, J. M .; Prideaux, G. J .; Laslett, G. M .; Baynes, A .; Smith, M. A .; Jones, R .; Smith, B.L. (06.06.2001). „Nové věky poslední australské megafauny: vyhynutí na celém kontinentu před zhruba 46 000 lety“ (PDF). Věda. 292 (5523): 1888–1892. Bibcode:2001Sci ... 292.1888R. doi:10.1126 / science.1060264. PMID 11397939. S2CID 45643228. Citováno 2011-08-26.
- ^ Callaway, E. (2016-09-21). "Human remains found in hobbit cave". Příroda. doi:10.1038/nature.2016.20656. S2CID 89272546.
- ^ Diamond, Jared (2008-08-13). "Paleontologie: Poslední klokan obrovský". Příroda. 454 (7206): 835–836. Bibcode:2008Natur.454..835D. doi:10.1038 / 454835a. PMID 18704074. S2CID 36583693.
- ^ Turney, C. S. M .; Flannery, T. F.; Roberts, R. G .; Reid, C .; Fifield, L. K.; Higham, T. F. G .; Jacobs, Z .; Kemp, N.; Colhoun, E. A.; Kalin, R. M.; Ogle, N. (2008-08-21). „Pozdně přežívající megafauna v Tasmánii v Austrálii implikuje lidskou účast na jejich vyhynutí“. PNAS. 105 (34): 12150–12153. Bibcode:2008PNAS..10512150T. doi:10.1073 / pnas.0801360105. PMC 2527880. PMID 18719103.
- ^ Roberts, R.; Jacobs, Z. (October 2008). "The Lost Giants of Tasmania" (PDF). Australasian Science. 29 (9): 14–17. Archivovány od originál (PDF) on 2011-09-27. Citováno 2011-08-26.
- ^ Norton, C. J .; Kondo, Y .; Ono, A .; Zhang, Y .; Diab, M. C. (2009-05-23). „Povaha vyhynutí megafaunů během přechodu MIS 3–2 v Japonsku“. Kvartérní mezinárodní. 211 (1–2): 113–122. Bibcode:2010QuInt.211..113N. doi:10.1016 / j.quaint.2009.05.002.
- ^ Faith, J. T.; Surovell, T. A. (2009-12-08). "Synchronous extinction of North America's Pleistocene mammals". Sborník Národní akademie věd. 106 (49): 20641–20645. Bibcode:2009PNAS..10620641F. doi:10.1073/pnas.0908153106. PMC 2791611. PMID 19934040.
- ^ Haynes, Gary (2009). "Introduction to the Volume". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Paleobiologie a paleoantropologie obratlovců. Springer. str. 1–20. doi:10.1007/978-1-4020-8793-6_1. ISBN 978-1-4020-8792-9.
- ^ Fiedel, Stuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Paleobiologie a paleoantropologie obratlovců. Springer. pp. 21–37. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Simmons, A. H. (1999). Faunální vyhynutí v ostrovní společnosti: kyperský lovec hrochů. Interdisciplinární příspěvky do archeologie. Nakladatelé Kluwer Academic / Plenum. p. 382. doi:10.1007 / b109876. ISBN 978-0-306-46088-3. OCLC 41712246.
- ^ Simmons, A. H.; Mandel, R. D. (December 2007). "Not Such a New Light: A Response to Ammerman and Noller". Světová archeologie. 39 (4): 475–482. doi:10.1080/00438240701676169. JSTOR 40026143. S2CID 161791746.
- ^ Steadman, D. W.; Martin, P. S.; MacPhee, R. D. E .; Jull, A. J. T .; McDonald, H. G .; Woods, C. A .; Iturralde-Vinent, M .; Hodgins, G. W. L. (2005-08-16). „Asynchronní vyhynutí pozdních kvartérních lenosti na kontinentech a ostrovech“. Proc. Natl. Acad. Sci. USA. 102 (33): 11763–11768. Bibcode:2005PNAS..10211763S. doi:10.1073 / pnas.0502777102. PMC 1187974. PMID 16085711.
- ^ Cooke, S. B .; Dávalos, L. M .; Mychajliw, A. M .; Turvey, S. T .; Upham, N. S. (2017). „Antropogenní vyhubení dominuje poklesům holocénu západoindických savců“. Výroční přehled ekologie, evoluce a systematiky. 48 (1): 301–327. doi:10.1146 / annurev-ecolsys-110316-022754.
- ^ Anderson, A .; Sand, C .; Petchey, F .; Worthy, T. H. (2010). „Faunal extinction and human habitable in New Caledonia: Initial results and implications of new research at the Pindai Caves“. Journal of Pacific Archaeology. 1 (1): 89–109. hdl:10289/5404.
- ^ White, A. W .; Worthy, T. H .; Hawkins, S .; Bedford, S .; Spriggs, M. (2010-08-16). „Megafaunal meiolaniid rohaté želvy přežily až do raného lidského osídlení ve Vanuatu v jihozápadním Pacifiku“. Proc. Natl. Acad. Sci. USA. 107 (35): 15512–15516. Bibcode:2010PNAS..10715512W. doi:10.1073 / pnas.1005780107. PMC 2932593. PMID 20713711.
- ^ Burney, D. A .; Burney, L. P .; Godfrey, L. R .; Jungers, W. L .; Goodman, S. M .; Wright, H. T .; Jull. A. J. T. (July 2004). "Chronologie pro pozdně prehistorický Madagaskar". Journal of Human Evolution. 47 (1–2): 25–63. doi:10.1016 / j.jhevol.2004.05.005. PMID 15288523.
- ^ Holdaway, R. N .; Jacomb, C. (2000-03-24). „Rapid Extinction of the Moas (Aves: Dinornithiformes): Model, Test, and Implications“. Věda. 287 (5461): 2250–2254. Bibcode:2000Sci ... 287,2250H. doi:10.1126 / science.287.5461.2250. PMID 10731144.
- ^ Janoo, A. (April 2005). "Discovery of isolated dodo bones (Raphus cucullatus (L.), Aves, Columbiformes) from Mauritius cave shelters highlights human predation, with a comment on the status of the family Raphidae Wetmore, 1930". Annales de Paléontologie. 91 (2): 167–180. doi:10.1016/j.annpal.2004.12.002.
- ^ Anderson, P. K. (July 1995). „Konkurence, predace a vývoj a zánik Stellerovy mořské krávy, Hydrodamalis gigas". Věda o mořských savcích. 11 (3): 391–394. doi:10.1111 / j.1748-7692.1995.tb00294.x. Archivovány od originál dne 2011-05-11. Citováno 2011-08-30.
- ^ Cooper, A .; Turney, C.; Hughen, K. A.; Brook, B. W .; McDonald, H. G .; Bradshaw, C. J. A. (2015-07-23). "Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover". Věda. 349 (6248): 602–6. Bibcode:2015Sci...349..602C. doi:10.1126/science.aac4315. PMID 26250679. S2CID 31686497.
- ^ Müller, U. C.; Pross, J.; Tzedakis, P. C.; Gamble, C.; Kotthoff, U.; Schmiedl, G.; Wulf, S.; Christanis, K. (February 2011). "The role of climate in the spread of modern humans into Europe". Kvartérní vědecké recenze. 30 (3–4): 273–279. Bibcode:2011QSRv...30..273M. doi:10.1016/j.quascirev.2010.11.016.
- ^ Biello, D. (2012-03-22). "Big Kill, Not Big Chill, Finished Off Giant Kangaroos". Scientific American news. Citováno 2012-03-25.
- ^ McGlone, M. (2012-03-23). "The Hunters Did It". Věda. 335 (6075): 1452–1453. Bibcode:2012Sci...335.1452M. doi:10.1126/science.1220176. PMID 22442471. S2CID 36914192.
- ^ A b C Rule, S.; Brook, B. W .; Haberle, S. G.; Turney, C. S. M .; Kershaw, A. P. (2012-03-23). "The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia". Věda. 335 (6075): 1483–1486. Bibcode:2012Sci...335.1483R. doi:10.1126/science.1214261. PMID 22442481. S2CID 26675232.
- ^ A b Johnson, C. N .; Alroy, J .; Beeton, N.J .; Bird, M. I .; Brook, B. W .; Cooper, A .; Gillespie, R .; Herrando-Pérez, S .; Jacobs, Z .; Miller, G. H .; Prideaux, G. J .; Roberts, R. G .; Rodríguez-Rey, M .; Saltré, F .; Turney, C. S. M .; Bradshaw, C. J. A. (10. února 2016). „Co způsobilo vyhynutí pleistocénní megafauny Sahul?“. Sborník Královské společnosti B: Biologické vědy. 283 (1824): 20152399. doi:10.1098 / rspb.2015.2399. PMC 4760161. PMID 26865301.
- ^ Miller, G. H .; Magee, J. W.; Johnson, B. J.; Fogel, M. L.; Spooner, N. A.; McCulloch, M. T.; Ayliffe, L. K. (1999-01-08). "Pleistocene Extinction of Genyornis newtoni: Human Impact on Australian Megafauna". Věda. 283 (5399): 205–208. doi:10.1126/science.283.5399.205. PMID 9880249.
- ^ Miller, G.; Magee, J.; Smith, M.; Spooner, N.; Baynes, A .; Lehman, S.; Fogel, M.; Johnston, H.; Williams, D .; Clark, P.; Florian, C.; Holst, R.; DeVogel, S. (2016-01-29). "Human predation contributed to the extinction of the Australian megafaunal bird Genyornis newtoni ∼47 ka". Příroda komunikace. 7: 10496. Bibcode:2016NatCo...710496M. doi:10.1038/ncomms10496. PMC 4740177. PMID 26823193.
- ^ Johnson, C. (2009-11-20). "Megafaunal Decline and Fall". Věda. 326 (5956): 1072–1073. Bibcode:2009Sci...326.1072J. doi:10.1126/science.1182770. PMID 19965418. S2CID 206523763.
- ^ Gill, J. L.; Williams, J. W.; Jackson, S. T.; Lininger, K. B.; Robinson, G. S. (2009-11-20). "Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America" (PDF). Věda. 326 (5956): 1100–1103. Bibcode:2009Sci...326.1100G. doi:10.1126/science.1179504. PMID 19965426. S2CID 206522597.
- ^ Fiedal, Stuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Paleobiologie a paleoantropologie obratlovců. Springer. pp. 21–37. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Martin, P. S. (2005). "Chapter 4. Ground Sloths at Home". Twilight of the Mammoths: Ice Age Extinctions and the Rewilding of America. University of California Press. pp. 78–99. ISBN 978-0-520-23141-2. OCLC 58055404. Citováno 2014-11-11.
- ^ Milman, Oliver (February 6, 2019). "The killing of large species is pushing them towards extinction, study finds". Opatrovník. Citováno 13. února 2019.
- ^ Ripple, W. J.; et al. (2019). "Are we eating the world's megafauna to extinction?". Dopisy o ochraně: e12627. doi:10.1111/conl.12627.
- ^ Alroy, J. (2008-08-12). "Dynamics of origination and extinction in the marine fossil record". PNAS. 105 Suppl 1 (Supplement_1): 11536–11542. Bibcode:2008PNAS..10511536A. doi:10.1073/pnas.0802597105. PMC 2556405. PMID 18695240.
- ^ Wolf, A.; Doughty, C. E.; Malhi, Y. (2013). "Lateral Diffusion of Nutrients by Mammalian Herbivores in Terrestrial Ecosystems". PLOS ONE. 8 (8): e71352. Bibcode:2013PLoSO...871352W. doi:10.1371/journal.pone.0071352. PMC 3739793. PMID 23951141.
- ^ Marshall, M. (2013-08-11). "Ecosystems still feel the pain of ancient extinctions". Nový vědec. Citováno 2013-08-12.
- ^ A b Doughty, C. E.; Wolf, A.; Malhi, Y. (2013-08-11). "The legacy of the Pleistocene megafauna extinctions on nutrient availability in Amazonia". Nature Geoscience. 6 (9): 761–764. Bibcode:2013NatGe...6..761D. doi:10.1038/ngeo1895.
- ^ Roman, J.; McCarthy, J.J. (2010). "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007.
- ^ A b C Wilkinson, D. M.; Nisbet, E. G.; Ruxton, G. D. (2012-05-08). "Could methane produced by sauropod dinosaurs have helped drive Mesozoic climate warmth?". Aktuální biologie. 22 (9): R292–R293. doi:10.1016/j.cub.2012.03.042. PMID 22575462. Citováno 2012-05-08.
- ^ "Dinosaur gases 'warmed the Earth'". BBC Nature News. 2012-05-07. Citováno 2012-05-08.
- ^ A b C Smith, F. A.; Elliot, S. M.; Lyons, S. K. (2010-05-23). "Methane emissions from extinct megafauna". Nature Geoscience. 3 (6): 374–375. Bibcode:2010NatGe...3..374S. doi:10.1038/ngeo877.
- ^ Kelliher, F. M.; Clark, H. (2010-03-15). "Methane emissions from bison—An historic herd estimate for the North American Great Plains". Zemědělská a lesní meteorologie. 150 (3): 473–577. Bibcode:2010AgFM..150..473K. doi:10.1016/j.agrformet.2009.11.019.
- ^ Helgen et. all, Kristofer M. (2006). "Ecological and evolutionary significance of sizes of giant extinct kangaroos" (PDF). Australian Journal of Zoology. 54 (4): 293–301. doi:10.1071 / ZO05077 – via si.edu.
- ^ Larramendi, A. (2016). "Shoulder height, body mass and shape of proboscideans" (PDF). Acta Palaeontologica Polonica. 61 (3): 537–574. doi:10.4202 / app.00136.2014. S2CID 2092950. Citováno 2018-03-22.
- ^ A b Fariña, Richard A .; Vizcaíno, Sergio F .; De Iuliis, Gerry (22. května 2013). Megafauna: Obří bestie pleistocénu v Jižní Americe. Indiana University Press. ISBN 978-0-253-00719-3. OCLC 779244424.
- ^ Zhang, Y .; Harrison, T. (2017). "Gigantopithecus blacki: obří lidoop z pleistocénu v Asii se vrátil “. American Journal of Physical Anthropology. 162 (S63): 153–177. doi:10.1002 / ajpa.23150. PMID 28105715.
- ^ Ruff, C. B .; Trinkaus, E .; Holliday, T. W. (08.05.1997). "Tělesná hmotnost a encefalizace v pleistocénu Homo". Příroda. 387 (6629): 173–176. Bibcode:1997 Natur.387..173R. doi:10.1038 / 387173a0. PMID 9144286. S2CID 4320413.
- ^ Grine, F. E .; Jumgers, W. L .; Tobias, P. V .; Pearson, O. M. (červen 1995). "Fosilní Homo stehenní kost z Berg Aukas v severní Namibii “. American Journal of Physical Anthropology. 97 (2): 151–185. doi:10.1002 / ajpa.1330970207. PMID 7653506.
- ^ Smith, Chris; Burger, Lee (listopad 2007). „Náš příběh: zkameněliny předků člověka“. Nahí vědci. Citováno 2011-02-19.
- ^ Kappelman, John (08.05.1997). „Mohli by to být obři“. Příroda. 387 (6629): 126–127. Bibcode:1997 Natur.387..126K. doi:10.1038 / 387126a0. PMID 9144276. S2CID 4328242.
- ^ de Barros Ferraz, K.M.P.M .; Bonach, K .; Verdade, L.M. (2005). "Vztah mezi tělesnou hmotností a délkou těla v kapybarách (Hydrochoerus hydrochaeris)". Biota Neotropica. 5 (1): 197–200. doi:10.1590 / S1676-06032005000100020.
- ^ Kitchener, A.C., Breitenmoser-Würsten, C., Eizirik, E., Gentry, A., Werdelin, L., Wilting, A. a Yamaguchi, N. (2017). „Revidovaná taxonomie kočkovitých šelem: závěrečná zpráva pracovní skupiny pro klasifikaci koček skupiny odborníků na kočky IUCN“ (PDF). Cat News (Zvláštní vydání 11).CS1 maint: používá parametr autoři (odkaz)
- ^ Brakefield, Tom (1993). Big Cats: Kingdom of Might. Voyageur Press. p.44. ISBN 978-0-89658-329-0.
- ^ Nowell, Kristin; Jackson, Peter (1996). Divoké kočky: Akční plán průzkumu stavu a ochrany přírody (PDF). Gland, Švýcarsko: IUCN / SSC Cat Specialist Group. p. 56. ISBN 978-2-8317-0045-8.
- ^ Kitchener, A. a Yamaguchi, N. (2009). „Co je to tygr? Biogeografie, morfologie a taxonomie“. In Tilson, R .; Nyhus, P. J. (eds.). Tigers of the World: The Science, Politics and Conservation of Panthera tigris. Akademický tisk. 53–84. ISBN 978-0-08-094751-8.CS1 maint: používá parametr autoři (odkaz)
- ^ Slaght, J. C., Miquelle, D. G., Nikolaev, I. G., Goodrich, J. M., Smirnov, E. N., Traylor-Holzer, K., Christie, S., Arjanova, T., Smith, J. L. D. a Karanth, K. U. (2005). „Kapitola 6. Kdo je králem zvířat? Historická a současná data o tělesné hmotnosti divokých a zajatých amurských tygrů ve srovnání s jinými poddruhy.“ (PDF). V D. G. Miquelle; E. N. Smirnov; J.M. Goodrich (eds.). Tigers in Sikhote-Alin Zapovednik: Ecology and Conservation (v Rusku). Vladivostok, Rusko: PSP. s. 25–35.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ http://www.ligerworld.com/samson-the-biggest-tiger.html
- ^ DeMaster, D.P .; Stirling, já (8. května 1981). "Ursus maritimus". Savčí druhy (145): 1–7. doi:10.2307/3504138. JSTOR 3503828.
- ^ Pasitschniak-Arts, M. (23. dubna 1993). "Ursus arctos". Savčí druhy (439): 1–10. doi:10.2307/3503828. JSTOR 3504138.
- ^ Soibelzon, L. H .; Schubert, B. W. (leden 2011). „Největší známý medvěd, Arctotherium angustidens, z raně pleistocénní pampejské oblasti Argentiny: s diskusí o trendech velikosti a stravování u medvědů “. Journal of Paleontology. 85 (1): 69–75. doi:10.1666/10-037.1. S2CID 129585554. Citováno 2011-06-01.
- ^ Swift, E. M. (1997-11-17). „Jaké velké ústa mají: Cestovatelé v Africe, kteří se dostanou do konfliktu s hrochy, se toho nemusí dočkat“. Sports Illustrated Vault. Time Inc. Citováno 2011-11-16.
- ^ ^ J. Calambokidis a G. Steiger (1998). Modré velryby. Voyageur Press. ISBN 0-89658-338-4.
- ^ ^ „Záznamy o zvířatech“. Smithsonianský národní zoologický park. Citováno 2007-05-29.
- ^ Anteosaurus Archivováno 2016-03-14 na Wayback Machine. Palaeos.org (22.04.2013)
- ^ Sulej, T .; Niedźwiedzki, G. (2019). „Pozdní triasový synapsid velikosti slona se vztyčenými končetinami“. Věda. 363 (6422): 78–80. Bibcode:2019Sci ... 363 ... 78S. doi:10.1126 / science.aal4853. PMID 30467179.
- ^ St. Fleur, Nicholas (4. ledna 2019). „Sloní příbuzný savců, kteří se pásli podél dinosaurů“. The New York Times. Citováno 6. ledna 2019.
- ^ Palmer, D. (1. července 2002). Marshallova ilustrovaná encyklopedie dinosaurů a prehistorických zvířat. Nové řádkové knihy. ISBN 978-1-57717-293-2. OCLC 183092423. Citováno 2013-06-10.
- ^ Ryby netvora rozdrtily odpor nejsilnějším skusem, jaký kdy byl. The Sydney Morning Herald. 30. listopadu 2006.
- ^ Anderson, P. S.L .; Westneat, M. W (2006-11-28). "Mechanika podávání a modelování kousací síly lebky Dunkleosteus terrelli, starodávný vrcholový predátor ". Biologické dopisy. 3 (1): 77–80. doi:10.1098 / rsbl.2006.0569. ISSN 1744-9561. PMC 2373817. PMID 17443970.
- ^ Anderson, P.S.L. (2010-05-04). "Použití vazebných modelů k prozkoumání kinematické rozmanitosti lebky a funkční konvergence v plaododermech arthrodire". Journal of Morphology. 271 (8): 990–1005. doi:10,1002 / jmor.10850. ISSN 0362-2525. PMID 20623651. S2CID 46604512.
- ^ Van Roy, P .; Daley, A. C .; Briggs, D. E. G. (11. března 2015). „Homologie anomalokaridových kmenových končetin byla odhalena obrovským podavačem filtrů se spárovanými klapkami“. Příroda. 522 (7554): 77–80. Bibcode:2015Natur.522 ... 77V. doi:10.1038 / příroda14256. PMID 25762145. S2CID 205242881.
- ^ Tsubamoto, T. (2012). "Odhad tělesné hmotnosti z astragalu u savců". Acta Palaeontologica Polonica: 259–265. doi:10.4202 / app.2011.0067. S2CID 54686160.
- ^ Palmer, D., ed. (1999). Marshallova ilustrovaná encyklopedie dinosaurů a prehistorických zvířat. London: Marshall Editions. p. 248. ISBN 978-1-84028-152-1.
- ^ Moyano, S.R .; Giannini, N.P. (10.10.2018). "Lebeční znaky spojené s postnatálním vývojem proboscis v Tapiru (Perissodactyla: Tapiridae) a srovnání s jinými existujícími a fosilními spárkavými savci". Zoologischer Anzeiger. 277 (7554): 143–147. doi:10.1016 / j.jcz.2018.08.005. ISSN 0044-5231.
- ^ Sample, Ian (19. února 2010). „Velký bílý žralok je více ohrožen než tygr, tvrdí vědec“. Opatrovník. Citováno 14. srpna 2013.