Radiodonta - Radiodonta
| Radiodonta | |
|---|---|
 | |
| Amplectobelua symbrachiata, Anomalocaris canadensis, Aegirocassis benmoulai, Peytoia nathorsti, Lyrarapax unguispinus, a Hurdia victoria | |
| Vědecká klasifikace | |
| Království: | |
| Kmen: | |
| Třída: | |
| Objednat: | †Radiodonta Collins, 1996 |
| Rodiny | |
Radiodonta je objednat skupiny kmenů členovci který byl celosvětově úspěšný během Kambrijský období a zahrnoval nejdříve známé velké predátory. Mohou být označovány jako radiodontů,[1][2][3] radiodontans,[4][5] radiodontidy,[6] anomalokaridy,[7] nebo anomalokarididy,[8][9][10] ačkoli tito původně odkazovali na rodinu Anomalocarididae, který dříve zahrnoval všechny druhy tohoto řádu, ale nyní je omezen pouze na několik druhů.[7] Mezi nejznámější druhy radiodontů patří Kambrijský taxony Anomalocaris canadensis, Hurdia victoria, Peytoia nathorsti, a Amplectobelua symbrachiata, Ordovik Aegirocassis benmoulai a Devonský Schinderhannes bartelsi.
Etymologie
Jméno Radiodonta (latinsky pro poloměr „mluvil o kole“ a řecky pro odoús „zub“) označuje radiální uspořádání zubních destiček (orální kužel) obklopujících ústa,[6] ačkoli tyto rysy jsou navrhovány být chybějící v některých druzích radiodont.[4][1]
Definice
Původní diagnóza řádu Radiodonta z roku 1996 je následující:[6]
Radiodontids jsou bilaterálně symetrické, protáhlé členovci s nemineralizovanou kutikulou, typicky nejrobustnější v čelistech a drápech. Tělo je rozděleno na dvě části tagmata, podobně jako prosoma a opisthosoma z chelicerate členovci. Přední část obvykle nevykazuje žádnou vnější segmentaci, nese jeden pár preorálních drápů, pár prominentních očí a břišní čelisti se zářivými zuby. Některé formy mají další řady zubů a tři nebo čtyři postorální gnathobasic páry končetin. Kmen je metamerické, typicky s přibližně 13 segmenty bočně se rozvíjejícími imobilizujícími laloky pro plavání a žábry pro dýchání, a mohou končit prominentním třídílným ocasem. Některé formy mají gnathobasické kmenové končetiny.
V roce 2014 byla definována clade Radiodonta fylogeneticky jako subtyp včetně jakýchkoli taxonů blíže k Anomalocaris canadensis než Paralithodes camtschaticus.[7] V roce 2019 byla nově definována morfologicky jako komplex krunýře hlavy s ložiskem zvířete s centrálními (H-) a postranními (P-) prvky; výrůstky (endity) z čelních přívěsků nesoucích pomocné trny; a snížené přední chlopně nebo pásy lamel (setální čepele) a silné zúžení těla od přední k zadní.[3]

Popis

Většina radiodontů byla podstatně větší než ostatní Kambrijský fauna s typickými délkami těla pohybujícími se od 30 do 50 centimetrů.[2] Největší popsaný radiodont je Ordovik Aegirocassis benmoulai, které mohly dorůst až dva metry.[10][2] Nejmenší hlásil radiodont, mladistvý Lyrarapax unguispinus, bylo jen asi 18 milimetrů.[11]
Tělo radiodontu lze rozdělit do dvou oblastí: hlavy a trupu. Hlava se skládá pouze z jednoho segmentu těla[12] známý jako oční somit, pokrytý sklerity (komplex krunýře hlavy), nesl artropodizované čelní přídavky, ventrální ústní ústrojí (orální kužel) a pronásledoval složené oči. Zatímco zužující se kufr se skládá z několika segmentů těla, z nichž každý je spojen s dvojicemi chlopní a žábrovitých struktur (setálních čepelí).[3]
Čelní přívěsek




Přední struktury na hlavě jsou dvojicí čelních přívěsků. V předchozích studiích mohou být označovány jako „drápy“, „uchopení přídavky“, „krmení přídavky“ nebo „velké přídavky“, ale v poslední době se již tyto přídavky nepoužívají, protože čelní přídavky byly považovány za nehomologické megacheiran skvělé přílohy.[12] Jsou sklerotizované a segmentované (arthropodized ), nesoucí ventrální trny (endity) na většině jejich segmentových jednotek (podomeres) a endity mohou nést další řady trnů (pomocné trny) na jejich předních a zadních okrajích.[13][3] Čelní přívěsek lze rozdělit do dvou oblastí: dříku (v předchozích studiích stopka) a distální kloubní oblasti.[13] Mezi ventrální stranou každého podomeru se může objevit trojúhelníková oblast pokrytá měkkou kutikulou (artrodiální membránou), která poskytuje pružnost.[14] Jejich preokulární a protocerebrální původ naznačuje, že jsou homologní s primárními anténami Onychophora a labrum z Euarthropoda (vše vzniklo z očního somitu) a není homologní s chelicerae z Chelicerata ani antény a skvělé přílohy dalších členovců, kteří jsou deutocerebrálně (také z postokulárního somitu 1).[9][12] Vzhledem k tomu, že morfologie frontálních přídavků, zejména enditů, se mezi druhy vždy liší, jsou jedním z nejdůležitějších znaků při identifikaci druhů.[13] Ve skutečnosti je mnoho radiodontů znám pouze z hrstky zkamenělých čelních přídavků.[14][13]

Čelní přídavky Anomalocarididae, Amplectobeluidae a případně příbuzné druhy.

Čelní přídavky Tamisiocarididae (="Cetiocaridae ").

Čelní přídavky Hurdiidae.



Orální kužel


Ústa jsou na břišní straně hlavy, za připojovacím bodem čelních přívěsků a jsou obklopena prstencem zubních destiček, které tvoří ústní část známou jako ústní kužel („čelisti“ v předchozích studiích[6]). Mohly by se zvětšit 3 nebo 4 zubové destičky, což by dalo orální kužel triradiální ( Anomalocaris ) nebo tetraradial (např. Hurdiidae, Lyrarapax ) vzhled.[15][11] Vnitřní okraj zubních destiček má hroty směřující k otvoru úst. U některých se mohou objevit další řady vnitřních zubových desek hurdiid rody.[8][3] Podrobná rekonstrukce amplectobeluid orální kužely jsou spekulativní, ale pravděpodobně nepředstavovaly typické radiální uspořádání.[4][1]
Sklerity hlavy, oči a trup




Tři sklerity hlavy (komplex krunýře hlavy) tvořící se centrálním H-prvkem (přední sklerit nebo štít hlavy) a dvojicí P-prvků (laterální sklerity) pokrývají dorzální a laterální povrch hlavy zvířete.[3] P-prvky se mohou navzájem spojovat, stejně jako H-prvky, úzkým předním prodloužením (hrdlo P-prvku nebo „zobák“).[8][3] Komplex hlavy krunýře je malý a vejčitý Anomalocarididae a Amplectobeluidae,[4][3] ale často se zvětší Hurdiidae.[3] Hlava nesla dvě pronásledoval složené oči, které mohou mít mobilitu,[16] a nachází se mezi mezerami, které se tvoří zadními oblastmi H-prvku a P-prvku.[8][3]


Na rozdíl od původní diagnózy může být rozdělení segmentů těla (segmentové hranice) viditelné zvenčí[10][5][3] a žádný známý člen Radiodonty (kromě domnělého radiodontu Cucumericrus[10][17]) je známo, že má přídavné části kufru (nohy).[18] Kmen má četné segmenty těla (somity ), zužující se od přední k zadní, s předními 3 nebo 4 segmenty výrazně zúženými do oblasti krku.[3]

Přídavky trupu byly ploutvové chlopně těla (v některých studiích „boční chlopně“ nebo „laloky“), obvykle jeden pár ventrální chlopně na každý segment těla, každý se mírně překrýval na jednom před ním, ale další, nepřekrývající se u některých se může objevit sada malých hřbetních chlopní Hurdiid druh.[10] Klapky mohou mít četné žilní struktury známé jako zesilující paprsky.[10][5][3] Chlopně v oblasti krku, nazývané přední chlopně nebo chlopně na krku, jsou výrazně sníženy. U některých druhů se z každé základny zmenšených chlopní krku zvedly přídavky na krmení podobné čelisti, které se nazývaly struktury podobné gnathobase (GLS).[4][1] Četné protáhlé čepelkovité prodloužení (kopinaté čepele) uspořádané v řadě, tvořící pásy žábry -jako struktura známá jako setální čepele nebo lamely,[3] pokrýval hřbetní povrch každého segmentu těla.[10] Alespoň v Aegirocassis, každá kopinatá čepel je pokryta vráskami.[10]
Ventrální chlopně mohou být homologní s endopodem biramních končetin euarthropodů a lobopodními končetinami (lobopody) žaber lobopodians a hřbetní laloky a lopatky setálu mohou být homologní s exopodem a hřbetními laloky nesoucími žáby dřívějších taxonů.[19][10] Kufr může končit buď 1 až 3 páry ocasního ventilátoru,[18][3] dvě dlouhé furcae,[20][3] podlouhlá koncová struktura,[18] nebo nevýrazný tupý tip.[10]

Aegirocassis, Peytoia a Hurdia. Tři hurdiid rody nesoucí hřbetní laloky a zvětšený komplex krunýře hlavy.

Anomalocaris canadensis, an Anomalokaridid radiodont s malým vejcovitým komplexem krunýře hlavy, 3 páry ocasního vějíře a koncovým struníkem.

Amplectobelua symbrachiata. An Amplectobeluid radiodont se strukturami podobnými gnathobase a dvěma furcae.



Vnitřní struktury

Stopy po svaly, zažívací ústrojí a nervový systém byly popsány z některých fosilií radiodontu. Páry dobře vyvinutých svalů, které byly spojeny s ventrálními chlopněmi, umístěnými v bočních dutinách každého segmentu těla.[18][9] Mezi bočními svaly je sofikovaná zažívací soustava, vytvořená rozšířením předního a zadního střeva, které jsou spojeny úzkým středním střevem spojeným se 6 páry střevní divercula (trávicí žlázy).[18][5][21] Ve srovnání s třísegmentovým mozek z euarthropod a dvousegmentový mozek onychophoran, mozek radiodontu se skládá pouze z jednoho mozkového segmentu pocházejícího z očního somitu, z protocerebra. Nervy čelních přívěsků a složených očí vznikly z přední a boční oblasti mozku.[9][12] Zadní část mozku byla dvojice zjevně nespojených ventrální nervový kabel který prošel oblastí krku zvířete.[9]
Paleoekologie
Fyziologie
Radiodonty byly interpretovány jako nektonický nebo nektobentický zvířata, jejichž morfologie naznačuje aktivní plavecký životní styl. Svalnaté, překrývající se ventrální chlopně mohly zvíře pohánět vodou, pravděpodobně pohybem ve vlnovité formaci připomínající moderní paprsky a sépie obecná.[22] Páry hřbetních chlopní, které u některých druhů tvoří ocasní ventilátor, mohou pomoci při řízení a / nebo stabilizaci zvířete během lokomoce.[10][23] v Anomalocaris, morfologie ocasního ventilátoru dokonce naznačuje, že by mohl rychle a efektivně měnit směr plavání.[24] Pásy setálních čepelí s vrásčitými kopinatými čepelemi mohou zvětšovat povrch, což naznačuje, že tomu tak bylo žábry, poskytující zvíře respirační funkce.[18][10] Hojnost pozůstatků sklerotizovaných struktur, jako jsou disartikulované čelní přídavky a komplex krunýře hlavy, naznačuje, že hmota línání k událostem mohlo dojít u radiodontů,[10][3] chování, které bylo také hlášeno u některých jiných Kambrijský členovci jako např trilobiti.[25]
Strava

Radiodonti měli různé stravovací strategie, které lze kategorizovat jako raptorial dravci, třídiče sedimentů nebo zavěšení, podavače filtrů.[2][26] Například draví dravci mají rádi Anomalocaris a Amplectobeluidy mohl být schopen chytit hbitou kořist pomocí svých raptoriálních frontálních přídavků, ten dokonce nesl robustní endite pro držení kořisti jako kleště.[17][14][4] S komplexem menšího krunýře hlavy a velkou plochou artrodiálních membrán měly čelní přídavky těchto taxonů větší flexibilitu.[20] Stout čelní přídavky prosévačů sedimentu jako Hurdia a Peytoia mají zoubkované endity s meziálním zakřivením, které by mohly tvořit košovou past na hrabání sedimentem a vedení potravin k dobře vyvinutému orálnímu kužele.[3] Některé druhy mohou mít jiné rysy, které se významně specializují na životní styl prosévání sedimentů, například Cambroraster s kopulovitým H-prvkem podobným krunýř a krab podkovy.[3] Endity čelních přívěsků z podavačů / filtrů Tamisiocaris a Aegirocassis mají pružné, hustě zabalené pomocné trny, které by mohly odfiltrovat organické složky, jako jsou mesozooplankton a fytoplankton až 0,5 mm.[7][10] Čelní přídavky Caryosyntrips, které jsou pro radiodonty neobvyklé tím, že mají směr koncových nosných povrchů proti sobě a mohly být schopné manipulovat a drtit kořist nůžkovým pohybem nebo krájením.[14][27]


Peytoia nathorsti, prosévání sedimentu hurdiid radiodont s tlustými čelními přídavky, úzce spojený s tetraradiálním orálním kuželem.

Čelní přívěsek zavěšení krmení radiodont Tamisiocaris borealis, ukazující protáhlý endit s hustě zabalenými pomocnými trny.
Spárované čelní přídavky Caryosyntrips, ukazující opačný směr.



Orální kužely radiodontů mohou být schopny sát a / nebo kousat.[15][26][3] Vedle čelních přídavků naznačuje diferenciace orálních kuželů mezi rody také preference odlišné stravy. Například triradiální orální kužel Anomalocaris s nepravidelnými, tuberkulovanými zuby a menším otvorem lze přizpůsobit malé a aktivní kořisti;[26] zatímco tuhé tetraradiální orální kužele Hurdia a Cambroraster s větším otvorem a dalšími zubovými destičkami může konzumovat větší potravinu.[26][3]
Klasifikace
Taxomomické spříznění
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Shrnutá fylogeneze mezi Radiodonta a jinými taxony Ecdysozoan.[28] |





Většina analýz naznačuje, že radiodonty jsou kmenové skupiny členovci[8][7][9][10][2][3] a sestra deuteropoda, clade včetně horního dříku (např. fuxianhuiids a škeble členovci) a koruna Euarthropoda (např. Artiopoda, Chelicerata a Mandibulata ).[28] Tuto interpretaci podporují četné rysy podobné členovcům, které se vyskytují na radiodontech, jako např složené oči,[16] zažívací žlázy,[21] artropodizace (na čelních přídavcích),[28][3] kmenové přídavky tvořené dorzálními a ventrálními prvky (předchůdci přídavků členovců biramus),[10][3] kutikularizovaný přední a zadní břicho,[18] stejně jako protocerebrální přední sklerit.[29] Zúžená oblast krku s doplňujícími se strukturami některých radiodontů může také osvětlit původ hlavy sophiciated arthropod, která je tvořena fúzí několika předních segmentů těla.[4][12]

"Žábry." lobopodians ' Opabinie (horní), Pambdelurion (vlevo dole) a Kerygmachela (vpravo dole).

Sibiřští lobopodiáni Siberion (horní levá), Megadictyon (dole uprostřed) a Jianshanopodia (vpravo nahoře).


Taxony jen bazální k radiodontu a větvi euarthropod jsou Pambdelurion, Kerygmachela a Opabinie, tři rádiodontové dinokaridid rody, které se obvykle označují jako „goblovité lobopodyny“[28][10] Mají chlopně těla, zažívací žlázy a specializované čelní přídavky jako radiodont, ale čelní přídavky nejsou artropodizované a pod každou z jejich chlopní nesou lobopod.[10] Opabinie je více odvozen tím, že má radiodont-like pronásledoval oči, ocasní ventilátor, setal lopatky, a dokonce i euarthropod-jako zadní ústa otevření.[28] Taxony, dokonce i bazální pro „goblovité lobopodiány“, jsou jako siberiidy Megadictyon a Jianshanopodia,[28] skupina lobopodians nesl robustní čelní přívěsky a zažívací žlázy, ale žádné klapky na těle. Tyto mezilehlé formy mezi lobopodiánem a radiodontem / euarthropodem naznačují, že celá skupina arthropoda vznikla z paraphyletic lobopodianský stupeň, vedle dalších dvou existujících panartropod phyla Tardigrada a Onychophora.[12]


Předchozí studie mohou navrhnout radiodonty jako skupinu jinou než kmenové členovce, jako je např cycloneuralia n červi prošli konvergentní s členovci (na základě cyklonuralistických radiálních ústí);[30] zastavit chelicerates vedle megacheira ns aka skvělý přívěsek členovci (na základě podobnosti mezi čelními přídavky radiodont, skvělými přídavky megacheiran a chelicerae );[31] nebo Schinderhannes bartelsi, který vyřešen jako a hurdiid radiodont v nedávných analýzách[28][7][10][2][3], jako druh více příbuzný euarthropodům než jiným radiodontům (na základě některých domnělých rysů podobných euarthropodům Schinderhannes).[23] Ani jeden z nich však nebyl podporován pozdějšími vyšetřováními. Radiální ústní ústrojí nejsou výlučně cyklonuralistická a je pravděpodobnějším současným výsledkem konvergentní evoluce nebo ecdysozoa n plesimorphy, protože byly také nalezeny v panarthropods jako tardigrade a nějaký lobopodia ns;[32] velké přídavky megacheiranů byly považovány za deutocerebrální,[33][34] které nejsou homologní s radiodontovými protocerebrálními čelními přídavky;[9][12] domnělé znaky euarthropod nalezené na singlu Schinderhannes fosilie je sporná a může představovat další struktury podobné radiodontům.[28]
Vzájemné vztahy
Radiodonta tradičně zahrnula všechny taxony do třídy Dinocaridida a taxony, které jsou v současné době zahrnuty do Radiodonty, byly umístěny do jedné rodiny, Anomalocarididae,[8][17] proto běžný název „anomalokaridid“ a stále se občas používal k označení celé objednávky i po reklasifikaci.[9][10] Nedávno bazální dinokarididní rody Pambdelurion, Kerygmachela a Opabinie byly umístěny mimo Radiondontu,[7][10][2][3] a většina druhů radiodontů byla překlasifikována do tří nových rodin: Amplectobeluidae, Tamisiocarididae[2][3] (dříve Cetiocaridae[7]), a Hurdiidae. Včetně Anomalocarididae, čtyři nedávné rodiny radiodontů mohou tvořit clade Anomalocarida.[7]

Včetně původního popisu objednávky Radiodonta Anomalocaris, Laggania (později známý jako Peytoia ), Hurdia, Proboscicaris, Amplectobelua, Cucumericrus, a Parapeytoia.[6] Nicméně, Proboscicaris je nyní považován za juniorské synonymum pro Hurdia, a Parapeytoia je považován za megacheiran.[8][18][10] Pozice Cucumericrus uvnitř Radiodonty je nejasný, protože byl buď nevybrán fylogenetickou analýzou[3] nebo vyřešen v polytomii s Radiodonta a Euarthropoda.[10]


Pod Radiodonta, Caryosyntrips je vedle sebe rodem nejvíce bazálním Cucumericrus (je-li součástí balení). Rod Anomalocaris vždy se ukázalo, žemonofyletický, obvykle s Anomalocaris kunmingensis a Anomalocaris briggsi vyřešen jako člen Amplectobeluidae a Tamisiocarididae resp.[7][9][10][2][3] Vzájemný vztah Amplectobeluidae je nejistý, protože afinita amplectobeluidů k Lyrarapax a Ramskoeldia byli občas vyslýcháni.[1][3] Monofilie ze zvláštní rodiny Hurdiidae je dobře podporováno několika odvozenými znaky (např. distální kloubní oblast čelního přívěsku s proximálními 5 podomery nesoucími podekvátní endity[13][3]), s Tamisiocarididae často navrhl být jeho sesterská skupina.[7][10][2][3]
- Radiodonta
- ?Cucumericrus
- Caryosyntrips
- Anomalocarida
- Paranomalocaris (některé studie uvádějí v rámci Anomalocarididae.[20])
- Laminacaris (některé studie uvádějí v rámci Amplectobeluidae.[2])
- Anomalocarididae
- Anomalocaris (některé druhy mohou být umístěny v jiných rodinách.[7][10])
- Amplectobeluidae
- Lyrarapax (pozice zpochybněná některými studiemi.[1])
- Amplectobelua
- Ramskoeldia (pozice zpochybněná některými studiemi.[3])
- Tamisiocarididae
- Hurdiidae
První hloubkovou fylogenetickou analýzu Radiodonty provedl Vinther et al. v roce 2014 a byla rozšířena o Cong et al. později ten rok přidáním Lyrarapax unguispinus.[9] Analýza byla v roce 2015 dále upravena Van Royem et al. s upravenými znaky a zahrnutím Cucumericrus decoratus a Aegirocassis benmoulai.[10]
|




| Druh | Deskriptory | Rok pojmenovaný | Rodina | Stáří | Umístění | Čelní přívěsek |
|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström a Ahlberg | 1995 | Neznámý | |||
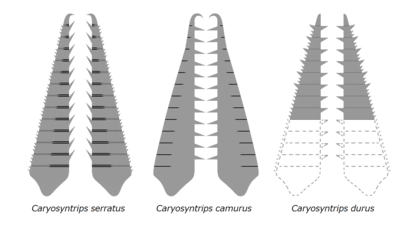
| Caryosyntrips serratus | Daley a Budd | 2010 | Wuliuan –Drumian |  | ||
| Caryosyntrips camurus | Pates & Daley | 2017 | Wuliuan |  | ||
| Caryosyntrips durus | Pates & Daley | 2017 | Drumian |  | ||
| Paranomalocaris multisegmentalis | Wang, Huang a Hu | 2013 |  | |||
| Laminacaris chiméra | Guo, Pates, Cong, Daley, Edgecombe, Chen a Hou | 2018 |  | |||
| Anomalocaris canadensis | Whiteaves | 1892 | Anomalocarididae |  | ||
| Anomalocaris pennsylvanica | Resser | 1929 |  | |||
| Anomalocaris kunmingensis | Wang, Huang a Hu | 2013 | ||||
| Anomalocaris magnabasis | Pates, Daley, Edgecombe, Cong & Lieberman | 2019 |  | |||
| Anomalocaris saron | Hou, Bergström a Ahlberg | 1995 |  | |||
| Anomalocaris briggsi | Nedin | 1995 | Tamisiocarididae |  | ||
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pates a Hou | 2018 | Amplectobeluidae |  | ||
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pates a Hou | 2018 | Amplectobeluidae |  | ||
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe a Strausfield | 2014 | Amplectobeluidae | Cambrian Fáze 3 |  | |
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou a Chen | 2016 | Amplectobeluidae | Cambrian Fáze 3 |  | |
| Amplectobelua symbrachiata | Hou, Bergström a Ahlberg | 1995 | Amplectobeluidae |  | ||
| Amplectobelua stephenensis | Daley a Budd | 2010 | Amplectobeluidae |  | ||
| Tamisiocaris borealis | Daley & Peel | 2010 | Tamisiocarididae |  | ||
| Ursulinacaris grallae | Paštiky, Daley & Butterfield | 2019 | Hurdiidae | Wuliuan |  | |
| Schinderhannes bartelsi | Kühl, Briggs a Rust | 2009 | Hurdiidae | Emsian | ||
| Stanleycaris hirpex | Pates, Daley a Ortega-Hernández | 2018 | Hurdiidae | |||
| Peytoia nathorsti | Walcott | 1911 | Hurdiidae |  | ||
| Peytoia infercambriensis | Lendzion | 1975 | Hurdiidae | Cambrian Fáze 3 | ||
| Aegirocassis benmoulai | Van Roy, Daley a Briggs | 2015 | Hurdiidae | Tremadocian |  | |
| Hurdia victoria | Walcott | 1912 | Hurdiidae | Wuliuan –Drumian |  | |
| Hurdia triangulata | Walcott | 1912 | Hurdiidae | Wuliuan | | |
| Cambroraster falcatus | Moysiuk & Caron | 2019 | Hurdiidae | Wuliuan |  | |
| Pahvantia hastata | Robison & Richards | 1981 | Hurdiidae | Drumian | ||
| Cordaticaris striatus | Slunce, Zeng a Zhao | 2020 | Hurdiidae | Drumian | ||
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier a Wang | 2006 | Hurdiidae | Neznámý |
Dějiny
Historie radiodontů je složitá. Neúplné vzorky týkající se různých částí těla stejného druhu byly historicky interpretovány jako patřící k různým druhům a dokonce k různým kmenům.[6][8] Před tím, než byly uznány jako skupina, byly radiodontové vzorky přiřazeny k pěti různým kmenům: Porifera, Cnidaria, Ostnokožci, Annelida, a Arthropoda.[6]
První známé radiodontní vzorky byly odebrány z trilobitové postele z Mount Stephen Richard G. McConnell z Geologická služba Kanady v roce 1886[6] nebo 1888.[36] Tyto vzorky byly pojmenovány Anomalocaris canadensis v roce 1892 paleontologem GSC Joseph Whiteaves.[36] Whiteaves interpretoval vzorky, nyní známé jako izolované čelní přílohy, jako břicho a fylokarid korýš. Další vzorky radiodontů byly popsány v roce 1911 autorem Charles Walcott. Interpretoval izolovaný orální kužel, který pojmenoval Peytoiajako medúza a špatně zachovaný, ale relativně úplný exemplář, který pojmenoval Lagganiajako holothurian. V roce 1912 pojmenoval Walcott Hurdia na základě izolovaného h-prvku, který interpretoval jako krunýř korýšů. A Hurdia P-prvek byl pojmenován Proboscicaris v roce 1962, a interpretován jako krunýř dvojzubce.
Geologická služba Kanady zahájila v roce 1966 revizi fosilií Burgess Shale, na kterou dohlížel Cambridge University paleontolog Harry B. Whittington.[6] Tato revize by nakonec vedla k objevu úplného plánu těla radiodontu. V roce 1978 Simon Conway Morris poznal, že ústa z Laggania byly Peytoia- jako, ale interpretoval to jako důkaz, že se jednalo o kompozitní fosilii tvořenou a Peytoia medúzy a houba.[37] V roce 1979 Derek Briggs poznal, že fosilie z Anomalocaris byly přívěsky, ne břicha, ale interpretovaly je jako chodící nohy.[38] Až v roce 1985 byla skutečná povaha fosilií Anomalocaris, Laggania, a Peytoia byl uznán a všichni byli přiřazeni k jedinému rodu, Anomalocaris. Následně to bylo uznáno Anomalocaris byla odlišná forma od ostatních dvou, což vedlo k rozdělení na dva rody, z nichž druhý byl různě pojmenován Laggania a Peytoia dokud nebylo rozhodnuto Peytoia měl přednost. Později bylo zjištěno, že některé fosilie přiřazené k těmto taxonům patří do jiné formy, která byla uznána jako nesoucí krunýř složený z Hurdia a Proboscicaris elementy. A konečně, v roce 2009 byly tyto vzorky přepsány jako Hurdia.[8] Radiodonta byla původně považována za obsahující jednu rodinu, Anomalocarididae, ale v roce 2014 byla rozdělena do čtyř rodin.[7]
Reference
- ^ A b C d E F Cong, Pei-Yun; Edgecombe, Gregory D .; Daley, Allison C .; Guo, Jin; Pates, Stephen; Hou, Xian-Guang (2018). „Nové radiodonty se strukturami podobnými gnathobase z kambrijské biosy Chengjiang a důsledky pro systematiku Radiodonty“. Články z paleontologie. 4 (4): 605–621. doi:10,1002 / spp2,1219. ISSN 2056-2802.
- ^ A b C d E F G h i j k Lerosey-Aubril, Rudy; Pates, Stephen (14.09.2018). „Nový radiodont s podáváním suspenze naznačuje vývoj mikroplanktivity v kambrijském makronektonu“. Příroda komunikace. 9 (1): 3774. doi:10.1038 / s41467-018-06229-7. ISSN 2041-1723. PMC 6138677. PMID 30218075.
- ^ A b C d E F G h i j k l m n Ó str q r s t u proti w X y z aa ab ac inzerát ae af ag Moysiuk, J .; Caron, J.-B. (2019-08-14). „Nový hurdiid radiodont z Burgess Shale dokazuje využívání kambrijských infaunálních zdrojů potravy“. Sborník Královské společnosti B: Biologické vědy. 286 (1908): 20191079. doi:10.1098 / rspb.2019.1079. PMC 6710600. PMID 31362637.
- ^ A b C d E F G Cong, Peiyun; Daley, Allison C .; Edgecombe, Gregory D .; Hou, Xianguang (2017-08-30). „Funkční hlava kambrijského radiodontanu (kmenová skupina Euarthropoda) Amplectobelua symbrachiata“. BMC Evoluční biologie. 17 (1): 208. doi:10.1186 / s12862-017-1049-1. ISSN 1471-2148. PMC 5577670. PMID 28854872.
- ^ A b C d Cong, Peiyun; Daley, Allison C .; Edgecombe, Gregory D .; Hou, Xianguang; Chen, Ailin (září 2016). "Morfologie radiodontan Lyrarapax z rané kambrijské biosy Chengjiang". Journal of Paleontology. 90 (4): 663–671. doi:10.1017 / jpa.2016.67. ISSN 0022-3360. S2CID 88742430.
- ^ A b C d E F G h i Collins, Desmond (1996). "Evoluce" Anomalocaris a jeho zařazení do třídy členovců Dinocarida (nov.) a řádu Radiodonta (nov.) ". Journal of Paleontology. 70 (2): 280–293. doi:10.1017 / S0022336000023362.
- ^ A b C d E F G h i j k l m Vinther, Jakob; Stein, Martin; Longrich, Nicholas R .; Harper, David A. T. (2014). „Anomalokarid zavěšený na krmení z raného kambriu“ (PDF). Příroda. 507 (7493): 496–499. doi:10.1038 / příroda13010. PMID 24670770. S2CID 205237459.
- ^ A b C d E F G h i Daley, Allison C .; Budd, Graham E .; Caron, Jean-Bernard; Edgecombe, Gregory D .; Collins, Desmond (2009). „Burgess Shale anomalocaridid Hurdia a jeho význam pro raný vývoj euarthropod “. Věda. 323 (5921): 1597–1600. doi:10.1126 / science.1169514. PMID 19299617. S2CID 206517995.
- ^ A b C d E F G h i j Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D .; Strausfeld, Nicholas J. (2014). "Struktura mozku řeší segmentovou afinitu anomalokaridových přídavků". Příroda. 513 (7519): 538–42. doi:10.1038 / příroda13486. PMID 25043032. S2CID 4451239.
- ^ A b C d E F G h i j k l m n Ó str q r s t u proti w X y z aa Van Roy, Peter; Daley, Allison C .; Briggs, Derek E. G. (2015). „Homologie anomalokaridových kmenových končetin byla odhalena obrovským podavačem filtrů se spárovanými klapkami“. Příroda. 522 (7554): 77–80. doi:10.1038 / příroda14256. PMID 25762145. S2CID 205242881.
- ^ A b Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A .; Shu, Degan; Paterson, John R. (01.11.2018). „Původ raptoriálního krmení u juvenilních euarthropodů odhalený kambrijským radiodontanem“. National Science Review. 5 (6): 863–869. doi:10.1093 / nsr / nwy057. ISSN 2095-5138.
- ^ A b C d E F G Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. (01.05.2017). „Původ a vývoj hlavy panartropodů - paleobiologická a vývojová perspektiva“. Struktura a vývoj členovců. Vývoj segmentace. 46 (3): 354–379. doi:10.1016 / j.asd.2016.10.011. ISSN 1467-8039. PMID 27989966.
- ^ A b C d E Pates, Stephen; Daley, Allison C .; Butterfield, Nicholas J. (06.06.2019). „První zpráva o spárovaných ventrálních enditech v hurdiidním radiodontu“. Zoologické dopisy. 5 (1): 18. doi:10.1186 / s40851-019-0132-4. ISSN 2056-306X. PMC 6560863. PMID 31210962.
- ^ A b C d Daley, Allison C .; Budd, Graham E. (2010). "Nové anomalokaridické přílohy z Burgess Shale, Kanada". Paleontologie. 53 (4): 721–738. doi:10.1111 / j.1475-4983.2010.00955.x. ISSN 1475-4983.
- ^ A b Daley, Allison C .; Bergström, leden (duben 2012). „Orální kužel Anomalocaris není klasika peytoia". Naturwissenschaften. 99 (6): 501–504. doi:10.1007 / s00114-012-0910-8. ISSN 0028-1042. PMID 22476406. S2CID 2042726.
- ^ A b Strausfeld, Nicholas J .; Ma, Xiaoya; Edgecombe, Gregory D .; Fortey, Richard A .; Land, Michael F .; Liu, Yu; Cong, Peiyun; Hou, Xianguang (srpen 2015). „Oči členovců: Časné kambrické fosilní záznamy a odlišný vývoj vizuálních systémů“. Struktura a vývoj členovců. 45 (2): 152–172. doi:10.1016 / j.asd.2015.07.005. PMID 26276096.
- ^ A b C d Xian-Guang, Hou; Bergström, Jan; Ahlberg, Per (září 1995). "Anomalocaris a další velká zvířata v dolní kambrijské fauně Chengjiang v jihozápadní Číně". GFF. 117 (3): 163–183. doi:10.1080/11035899509546213. ISSN 1103-5897.
- ^ A b C d E F G h Daley, Allison C .; Edgecombe, Gregory D. (2014). "Morfologie Anomalocaris canadensis z měšťanské břidlice “. Journal of Paleontology. 88 (1): 68–91. doi:10.1666/13-067. S2CID 86683798.
- ^ Van Roy, Peter; Daley, Allison C .; Briggs, Derek E. G. (2013). Anomalokarididy měly dvě sady bočních chlopní. 57. výroční zasedání Paleontologické asociace. Curych, Švýcarsko.
- ^ A b C Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A .; Shu, Degan; Paterson, John R. (01.11.2018). „Původ raptoriálního krmení u juvenilních euarthropodů odhalený kambrijským radiodontanem“. National Science Review. 5 (6): 863–869. doi:10.1093 / nsr / nwy057. ISSN 2095-5138.
- ^ A b Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2014-05-02). „Sofistikovaný trávicí systém u časných členovců“. Příroda komunikace. 5 (1): 3641. doi:10.1038 / ncomms4641. ISSN 2041-1723. PMID 24785191.
- ^ Usami, Yoshiyuki (01.01.2006). "Teoretická studie o formě těla a plaveckém vzoru Anomalocaris na základě hydrodynamické simulace". Journal of Theoretical Biology. 238 (1): 11–17. doi:10.1016 / j.jtbi.2005.05.008. ISSN 0022-5193. PMID 16002096.
- ^ A b Kühl, Gabriele; Briggs, Derek E. G .; Rust, Jes (06.02.2009). „Skvělý dodatek s členovcem s radiálními ústy z dolní devonské břidlice Hunsrück, Německo“. Věda. 323 (5915): 771–773. doi:10.1126 / science.1166586. ISSN 0036-8075. PMID 19197061. S2CID 47555807.
- ^ Sheppard, K. A .; Rival, D.E .; Caron, J.-B. (2018-10-01). „O hydrodynamice ocasních ploutví Anomalocaris“. Integrativní a srovnávací biologie. 58 (4): 703–711. doi:10.1093 / icb / icy014. ISSN 1540-7063. PMID 29697774.
- ^ Daley, Allison; Drage, Harriet (01.09.2015). „Fosilní záznam ekdýzy a trendy v línání trilobitů“. Struktura a vývoj členovců. 45 (2): 71–96. doi:10.1016 / j.asd.2015.09.004. PMID 26431634.
- ^ A b C d De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob (2016-12-16). „Rekonstrukce anomalokarididového krmného přívěsku obratnost vrhá světlo na ekologii radiodontanů“. Citovat deník vyžaduje
| deník =(Pomoc) - ^ Pates, S .; Daley, A. C. (2017). „Caryosyntrips: radiodontan ze kambrijského Španělska, USA a Kanady“. Články z paleontologie. 3 (3): 461–470. doi:10,1002 / spp2,1084. ISSN 2056-2802.
- ^ A b C d E F G h i Ortega-Hernández, Javier (prosinec 2014). „Cítit smysl pro„ dolní “a„ horní “kmenovou skupinu Euarthropoda, s poznámkami o přísném používání jména Arthropoda von Siebold, 1848“. Biologické recenze Cambridge Philosophical Society. 91 (1): 255–273. doi:10.1111 / brv.12168. ISSN 1469-185X. PMID 25528950. S2CID 7751936.
- ^ Ortega-Hernández, Javier (2015-06-15). „Homologie skleritů hlavy u euarthropodů břidlicových břidlic“. Aktuální biologie. 25 (12): 1625–1631. doi:10.1016 / j.cub.2015.04.034. ISSN 0960-9822. PMID 25959966.
- ^ Xian-Guang, Hou; Bergström, Jan; Ahlberg, Per (září 1995). „Anomalocaris a další velká zvířata v dolní kambrijské fauně Chengjiang v jihozápadní Číně“. GFF. 117 (3): 163–183. doi:10.1080/11035899509546213. ISSN 1103-5897.
- ^ Haug, Joachim T .; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin (březen 2012). „Funkční morfologie, ontogeneze a vývoj kudlankovitých dravců podobných v kambriu: MANTIS SHRIMP-JAKO KAMBRIÁNSKÉ PREDÁTORY“. Paleontologie. 55 (2): 369–399. doi:10.1111 / j.1475-4983.2011.01124.x.
- ^ Smith, Martin R .; Caron, Jean-Bernard (červen 2015). "Halucigenia hlava a hltanová armatura časných ekdyzozoů" (PDF). Příroda. 523 (7558): 75–78. doi:10.1038 / příroda1473. ISSN 1476-4687. PMID 26106857. S2CID 205244325.
- ^ Tanaka, Gengo; Hou, Xianguang; Ma, Xiaoya; Edgecombe, Gregory D .; Strausfeld, Nicholas J. (říjen 2013). "Chelicerate neurální pozemní vzor v kambriu velký přívěsek členovec". Příroda. 502 (7471): 364–367. doi:10.1038 / příroda12520. ISSN 1476-4687. PMID 24132294. S2CID 4456458.
- ^ Ortega-Hernández, Javier; Lerosey-Aubril, Rudy; Pates, Stephen (2019-12-18). „Sklon nervového systému v kambrijských Burgessových ložiskách břidlicového typu“. Sborník Královské společnosti B: Biologické vědy. 286 (1917): 20192370. doi:10.1098 / rspb.2019.2370. PMC 6939931. PMID 31822253.
- ^ Sun, Zhixin; Zeng, Han; Zhao, Fangchen (01.08.2020). „Nový střední kambrijský radiodont ze severní Číny: důsledky pro morfologickou disparitu a prostorové rozdělení hurdiidů“. Paleogeografie, paleoklimatologie, paleoekologie. 558: 109947. doi:10.1016 / j.palaeo.2020.109947. ISSN 0031-0182.
- ^ A b Whiteaves, J. F. (1892). „Popis nového rodu a druhu phyllocarid Crustacea ze středního kambriu z Mount Stephen, B. C.“. Kanadský záznam vědy. 5 (4).
- ^ Conway Morris, S. (1978). „Laggania cambria Walcott: Kompozitní fosilie“. Journal of Paleontology. 52 (1): 126–131. JSTOR 1303799.
- ^ Briggs, D. E. G. (1979). „Anomalocaris, největší známý kambrijský členovec“. Paleontologie. 22 (3): 631–664.