Ostrovní gigantismus - Island gigantism

Ostrovní gigantismusnebo ostrovní gigantismus, je biologický jev, při kterém se velikost živočišného druhu izolovaného na ostrově dramaticky zvyšuje ve srovnání s jeho pevninou příbuznými. Ostrovní gigantismus je jedním z aspektů obecnějších „ostrovní efekt“ nebo „Fosterovo pravidlo“, což předpokládá, že když zvířata na pevnině kolonizují ostrovy, malé druhy mají tendenci vyvíjet větší těla a velké druhy mají tendenci vyvíjet menší těla (ostrovní nanismus ). Po příchodu lidí a přidružených zavlečených predátorů (psi, kočky, krysy, prasata) se z mnoha ostrovních i jiných ostrovních endemit stalo vyhynulý. Podobné zvýšení velikosti a zvýšené dřevnatosti bylo pozorováno u některých ostrovních rostlin.
Možné příčiny

Velká savčí masožravci často na ostrovech chybějí z důvodu nedostatečného dosahu nebo obtíží rozptýlení nad vodou. V jejich nepřítomnosti ekologické výklenky pro velké dravce mohou být obsazeni ptáky, plazy nebo menšími šelmy, které pak mohou dorůst do větší než normální velikosti. Například na pravěku Ostrov Gargano v Miocén -Pliocén Středomoří na ostrovech v karibský jako Kuba a dále Madagaskar a Nový Zéland, některé nebo všechny vrcholní dravci byli ptáci jako Orli, sokoli a sovy, včetně některých z největších známých příkladů těchto skupin. Ptáci a plazi však obecně vytvářejí méně účinné velké predátory než pokročilé šelmy.
Protože malá velikost to obvykle usnadňuje býložravci k úniku nebo skrytí před predátory může snížený predační tlak na ostrovy umožnit jejich zvětšení.[1][A] Malým býložravcům může také prospět absence konkurence z důvodu chybějících druhů velkých býložravců.
Mezi výhody velkých rozměrů, které byly navrženy pro ostrovní želvy, patří snížená náchylnost k nedostatku potravy a / nebo vody, schopnost přežít bez nich delší intervaly nebo schopnost cestovat za nimi na delší vzdálenosti. Období takového nedostatku může být na oceánských ostrovech větší hrozbou než na pevnině.[4]
Ostrovní gigantismus je tedy obvykle evolučním trendem vyplývajícím z odstranění omezení velikosti malých zvířat souvisejících s predací a / nebo soutěží.[5] Taková omezení však mohou fungovat odlišně v závislosti na velikosti zvířete; například zatímco malí býložravci mohou uniknout predátorům skrýváním, velcí býložravci mohou odradit predátory zastrašováním. Jako výsledek, doplňkový jev ostrovní nanismus může být také důsledkem odstranění omezení souvisejících s predací a / nebo soutěží o velikost velkých býložravců.[6] Naproti tomu ostrovní nanismus mezi predátory častěji vyplývá z uvalení omezení souvisejících s omezenými zdroji kořisti dostupnými na ostrovech.[6] Na rozdíl od ostrovního nanismu se ostrovní gigantismus vyskytuje u většiny velkých obratlovců skupiny a v bezobratlých.
Teritorialismus může upřednostňovat vývoj ostrovního gigantismu. Studie o Ostrov Anaho v Nevadě stanoveno, že druhy plazů, které byly teritoriální, měly tendenci být na ostrově větší než ve srovnání s pevninou, zejména u menších druhů. U teritoriálních druhů větší velikost umožňuje jednotlivcům lépe soutěžit o obranu svého území. To dává další impuls evoluci směrem k větší velikosti v ostrovní populaci.[7]
Dalším prostředkem k nastolení ostrovního gigantismu může být a zakladatelský efekt funkční, když větší členové populace pevniny mají lepší schopnost kolonizovat ostrovy.[8]
Velikost ostrova hraje roli při určování rozsahu gigantismu. Menší ostrovy obecně zrychlují rychlost vývoje změn ve velikosti organismu a organismy tam vyvíjejí větší extrémy velikosti.[9]
Příklady
Mezi příklady ostrovního gigantismu patří:
Savci
Mnoho hlodavců se na ostrovech zvětšuje šelmy, proboscideans a artiodaktyly obvykle se zmenšují.
Eulipotyphlans
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Korsický obří rejsek | Asoriculus corsicanus | Korsika | Vyhynulý (před 500 př. n.l.) |  Rudé zuby |
| Baleárská obří rejska | Asoriculus hidalgo | Mallorca a Menorca | Vyhynulý | |
| Sardinská obrovská rejska | Asoriculus similis | Sardinie | Vyhynulý | |
 Deinogalerix | Deinogalerix spp. | Ostrov Gargano | Vyhynulý (Pozdní miocén ) |  Měsíční krysy |
Hlodavci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
| Obří hutie s tupými zuby | Amblyrhiza inundata | Anguilla a Svatý Martin | Vyhynulý (Pleistocén ) |  Neotropické ostnaté krysy | |
| Větší jamajská obří hutie | Clidomys osborni | Jamaica | Vyhynulý (Pozdní pleistocén) | ||
| Desková ozubená hutie | Elasmodontomys obliquus | Portoriko | Vyhynulý (c. 1 nl) | ||
| Myš se zkroucenými zuby | Quemisia gravis | Hispaniola | Vyhynulý | ||
| Stromová obří hutie[10] | Tainotherium valei | Portoriko | Vyhynulý | ||
| Malá jamajská obří hutie | Xaymaca fulvopulvis | Jamaica | Vyhynulý | ||
| Mallorský obr křečci | Apocricetus darderi Tragomys macpheei | Mallorca | Vyhynulý | Apocricetus alberti[11] Cricetus kormosi[12] | |
 Obří křeček Gargano | Hattomys gargantua | Ostrov Gargano | Vyhynulý | ||
 Polní myš St Kilda | Apodemus sylvaticus hirtensis | St Kilda | Nejméně obavy |  Dřevěná myš | MR ≈ 2 [13] |
 Henselova polní myš | Rhagamys orthodon | Korsika a Sardinie | Vyhynulý (Pozdní pleistocén ) | ||
 Obří krysa na Tenerife | Canariomys bravoi | Tenerife | Vyhynulý (Pozdní pleistocén) | Africké krysy nosorožce | |
 Krysa z Gran Canaria | Canariomys tamarani | Gran Canaria | Vyhynulý (před rokem 1500) | ||
| Formentera černý-sledoval zahradní plch | Eliomys quercinus ophiusae | Formentera | Vzácný[14] |  Plch zahradní a jiný Leithiinae dormice | |
| Minorský obří plch | Hypnomys mahonensis | Menorca | Vyhynulý | ||
 Mallorský obří plch | Hypnomys morpheus | Mallorca | Vyhynulý | ||
| Sicilský obří plch | Leithia cartei | Sicílie | Vyhynulý | ||
| Maltský obří plch | Leithia melitensis | Malta | Vyhynulý | ||
 Orkney vole | Microtus arvalis orcadensis | Orknejské ostrovy | Zranitelný |  Hraboš obecný a jiný luční hraboši | |
 Obří hraboši Gargano | Mikrotia magna M. maiuscula M. parva | Ostrov Gargano | Vyhynulý (Časný pliocén) | ||
 St Kilda dům myš | Mus musculus muralis | St Kilda | Vyhynulý (kolem r. 1930) |  Dům myš | |
 Flores obří krysa | Papagomys armandvillei | Flores | Blízko ohrožení |  Severoafrický černá krysa a další skutečné krysy | |
| Sulawesi obří krysa | Paruromys dominator | Sulawesi | Nejméně obavy | ||
| Admiralita obří krysa | Rattus detentus | Ostrov Manus | Neznámý / Pravděpodobně ohrožený[15] | ||
| Populace černé krysy Congreso[16] | Rattus rattus | Isla del Congreso | Nejméně obavy | ||
| Norské jeleni myší | Peromyscus anyapahensis P. nesodytes | Kalifornské ostrovy Severního kanálu | Vyhynulý (kolem 6000 př. n.l.) |  Severoamerická myš jelena | |
 Obří plch Gargano | Stertomys laticrestatus[17] | Ostrov Gargano | Vyhynulý |  Glirinae dormice |
Lagomorphs
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Minorský obří zajícovec | Nuralagus rex | Menorca | Vyhynulý (Střední pliocén ) | Alilepus (?) Trischizolagus (?) |
| Prolagus imperialis | Ostrov Gargano | Vyhynulý |  Pikas | |
 Sardinská pika | Prolagus sardus | Korsika, Sardinie a Tavolara | Vyhynulý (kolem r. 1800) |
Primáti
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Opice Hispaniola | Antillothrix bernensis | Hispaniola | Vyhynulý (před inzerátem 1600) |  Cheracebus |
| Haitská opice | Insulacebus toussaintiana | Jihozápadní Haiti | Vyhynulý | |
 Kubánské opice | Paralouatta marianae[18] P. varonai[18] | Kuba | Vyhynulý (Pleistocén) | |
| Jamajská opice | Xenothrix mcgregori | Jamaica | Vyhynulý | |
 Lemur gorila | Archaeoindris fontoynontii | Centrální Madagaskar | Vyhynulý (c. 350 př.nl) |  Lorisoidy |
 Paviáni lemurů | Archaeolemur spp. Hadropithecus spp. | Madagaskar | Vyhynulý (před rokem 1280) | |
 Lenost lemurů | Babakotia spp. Palaeopropithecus spp. | Západní a střední Madagaskar | Vyhynulý (asi 1500 nl) | |
 Koala lemurů | Megaladapis edwardsi M. grandidieri M. madagascariensis | Madagaskar | Vyhynulý (1280-1420 nl) |
Carnivorans
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Vydra sardinská | Megalenhydris barbaricina | Sardinie | Vyhynulý (Pozdní pleistocén) |  Vydry |
 Fossa | Cryptoprocta ferox | Madagaskar | Zranitelný |  Mongoosy |
 Obří fossa | Cryptoprocta spelaea | Madagaskar | Vyhynulý (před AD 1400) |
Ptactvo
Kmenových ptáků
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Balaur | B. bondoc | Ostrov Hateg | Vyhynulý (Pozdní křída) |  Jeholornis[19] |
 Gargantuavis | G. philohinos | Ibero -Armorican ostrov | Vyhynulý (Pozdní křída ) |  Patagopteryx (?) |
Ptáci nadřádu běžci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Kiwi | Apteryx spp. | Nový Zéland | Variabilní | Proapteryx[b] |
 Větší sloní ptáci | Aepyornis hildebrandti A. maximus Vorombe titán | Madagaskar | Vyhynulý (asi 1000 nl) | |
 Menší sloni ptáci | Mullerornis spp. | Madagaskar | Vyhynulý (kolem r. 1260) | |
 Moas | Anomalopteryx didiformis Dinornis spp. Emeus crassus Euryapteryx spp. Megalapteryx didinus Pachyornis spp. | Nový Zéland | Vyhynulý (před AD 1445) |  Tinamous |
Vodní ptáci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Novozélandská pižmová kachna | Biziura delautouri | Nový Zéland | Vyhynulý (po 1500 nl) |  Australan pižmová kachna |
 Novozélandské husy | Cnemiornis calcitrans C. gracilis | Nový Zéland | Vyhynulý |  Cape Barren husa |
 Garganornis | G. ballmanni | Gargano a Scontrone ostrovy | Vyhynulý (Pozdní miocén ) |  Husy[21] |
 Moa-nalo s želvami | Chelychelynechen quassus | Kauai | Vyhynulý (asi 1000 nl) |  Dabing kachny |
 Malý-účtoval moa-nalo | Ptaiochen pau | Maui | Vyhynulý (asi 1000 nl) | |
Velkokapacitní moa-nalo | Thambetochen chauliodous | Maui Nui | Vyhynulý (asi 1000 nl) | |
 O'ahu moa-nalo | Thambetochen xanion | O'ahu | Vyhynulý (asi 1000 nl) | |
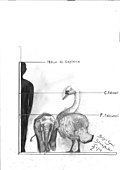 Obří labuť | Cygnus falconeri | Sicílie a Malta | Vyhynulý (Střední pleistocén ) |  Ztlumit labuť |
| Scarlettova kachna | Malacorhynchus scarletti | Nový Zéland | Vyhynulý (po 1500 nl) |  Kachna růžová |
Pangalliformes
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Hromadný stavitel megapodů | Megapodius molistructor | Nová Kaledonie a Tonga | Vyhynulý (asi 1500 př. n.l.) |  Drhnout |
| Ušlechtilý megapod | Megavitiornis altirostris | Fidži | Vyhynulý |  Hrabaví |
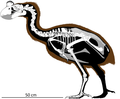 Nový kaledonský obří megapod | Sylviornis neocaledoniae | Nová Kaledonie a Isle of Pines | Vyhynulý |
Gruiformes
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Červená kolejnice | Aphanapteryx bonasia | Mauricius | Vyhynulý (c. AD 1700) |  Kolejnice |
 Hawkinsova kolejnice | Diaphanapteryx hawkinsi | Chathamské ostrovy | Vyhynulý (kolem r. 1900) | |
 Antilleanská jeskyně | Nesotrochis debooyi | Portoriko a Panenské Ostrovy | Vyhynulý | |
| Kubánská jeskynní zábradlí | Nesotrochis picapicensis | Kuba | Vyhynulý | |
| Haitská jeskynní kolejnice | Nesotrochis steganinos | Hispaniola | Vyhynulý | |
 Jižní ostrov takahē | Porphyrio hochstetteri | Jižní ostrov, Nový Zéland | Ohrožený | |
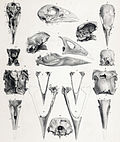 Severní ostrov takahē | Porphyrio mantelli | Severní ostrov, Nový Zéland | Vyhynulý (před rokem 1900) | |
 Adzebills | Aptornis defossor A. otidiformis | Nový Zéland | Vyhynulý |  Madagaskarský chmýří[22] |
 Chatham lyska | Fulica chathamensis | Chathamské ostrovy | Vyhynulý (po 1500 nl) |  Červeně hrbolatý lyska a další lysky |
 Mascarene lyska | Fulica newtonii | Mauricius a Shledání | Vyhynulý (c. AD 1700) | |
| Lyska novozélandská | Fulica prisca | Nový Zéland | Vyhynulý (po 1280 nl) | |
 Réunionské bažiny | Porphyrio coerulescens | Plaine des Cafres, Shledání | Vyhynulý (kolem r. 1730) |  Fialové bažiny |
Holubi
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Obří holub Viti Levu | Natunaornis gigoura | Viti Levu, Fidži | Vyhynulý |  Korunované holuby |
 Rodrigues solitaire | Pezophaps solitaria | Rodrigues | Vyhynulý (před AD 1778) |  Nicobar holub |
 Dodo | Raphus cucullatus | Mauricius | Vyhynulý (kolem r. 1662) |
Draví ptáci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Jeskyně Liko Golden Eagle | Aquila chrysaetos simurgh | Kréta | Vyhynulý (Pozdní pleistocén) |  Zlatý orel |
| Obří krab-jestřáb[23] | Buteogallus borrasi | Kuba | Vyhynulý |  Velký černý jestřáb a další jestřáby |
| Obří jestřáb | Gigantohierax sp. | Kuba | Vyhynulý | |
| Titan-jestřáb | Titanohierax gloveralleni | Kuba, Hispaniola a Bahamy | Vyhynulý | |
| Jamajský caracara | Caracara tellustris | Jamaica | Vyhynulý |  Caracaras |
| Oční harrier | Circus eylesi | Nový Zéland | Vyhynulý (asi 1000 nl) |  Bažina harrier |
| Orli na ostrově Gargano | Garganoaetus freudenthali G. murivorus | Ostrov Gargano | Vyhynulý (Pozdní miocén) | Aquila delphinensis |
 Haastův orel | Hieraaetus moorei | Nový Zéland | Vyhynulý (c. AD 1400) |  Malý orel |
 Filipínský orel | Pithecophaga jefferyi | Filipíny | Kriticky ohrožený |  Bateleur[24] |
Papoušci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Hercules papoušek | Heracles inexpectatus | Nový Zéland | Vyhynulý (Miocén ) |  Ostatní papoušci |
 Kakapo | Strigops habroptilus | Nový Zéland | Kriticky ohrožený | |
 Papoušek obecný | Lophopsittacus mauritianus | Mauricius | Vyhynulý (kolem r. 1680) |  Psittaculin papoušci |
Sovy
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Krétská sova | Athene cretensis | Kréta | Vyhynulý (Pleistocén) |  Sýček |
 Kubánské obří sovy | Ornimegalonyx spp. | Kuba | Vyhynulý (Pleistocén) |  Dřevěné sovy |
 Obří sova větší Gargano | Tyto gigantea | Ostrov Gargano | Vyhynulý (Pozdní miocén ) |  Sovy pálené |
 Sova pálená na ostrově Andros | Tyto pyly | Androsův ostrov, Bahamy | Vyhynulý (před inzerátem 1600) | |
 Riverova sova pálená | Tyto riveroi | Kuba | Vyhynulý | |
 Malá sova Gargano | Tyto robusta | Ostrov Gargano | Vyhynulý (Časný pliocén ) |
Caprimulgiformes
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Nový Zéland owlet-nightjar | Aegotheles novazelandiae | Nový Zéland | Vyhynulý (asi 1200 nl) |  Australský owlet-nightjar |
 New Caledonian owlet-nightjar | Aegotheles savesi | Nová Kaledonie | Kriticky ohrožený |
Passeriforms
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Chatham havran | Corvus moriorum | Chathamské ostrovy | Vyhynulý | Havran Nového Zélandu |
 Strnad dlouhý | Emberiza alcoveri | Tenerife | Vyhynulý (po inzerátu 1) |  Cabanis je strnad |
| Obří nukupu'u | Hemignathus vorpalis | Havaj | Vyhynulý (po 1000 nl) |  Pěnkavy |
| Tasmánský vynikající víla | Malurus cyaneus cyaneus | Tasmánie | Nejméně obavy |  Vynikající pohádky |
| Klokaní ostrov vynikající víly | Malurus cyaneus ashbyi | Klokaní ostrov | Nejméně obavy | |
| Stout nohama střízlík | Pachyplichas yaldwyni | Jižní ostrov z Nový Zéland | Vyhynulý |  Ostatní pěvci |
 Kozoroh silvereye | Zosterops lateralis chlorocephalus | Skupina Kozoroha a bunkru z Australan Velký bariérový útes | Neznámý |  Silvereye |
Plazi
Pterosaurs
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Hatzegopteryx | H. thambema | Ostrov Hateg | Vyhynulý (Pozdní křída) |  Quetzalcoatlus |
Iguanids
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
 Tonganský obří leguán[25] | Brachylophus gibbonsi | Tonga | Vyhynulý (asi 800 př. n.l.) |  jihoamerický leguáni | |
 Fidžijský obří leguán [26] | Lapitiguana impensa | Fidži | Vyhynulý (kolem 1000 př. n.l.) | ||
 Angel Island chuckwalla | Sauromalus hispidus | Isla Ángel de la Guarda, Baja California | Blízko ohrožení | Poloostrovní chuckwalla | MR ≈ 5 [27] |
 San Esteban chuckwalla | Sauromalus varius | Ostrov San Esteban, Baja California | Ohrožený | MR ≈ 5 [27] |
Gekoni
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
 Delcourtův obří gekko | Hoplodactylus delcourti | Nový Zéland | Vyhynulý (c. AD 1870) |  Diplodactylid gekoni | LR ≈ 6,75 [C] |
 Nový kaledonský obří gekon | Rhacodactylus leachianus | Nová Kaledonie | Nejméně obavy | LR ≈ 4,4 [d] MR ≈ 60 [E] | |
 Rodrigues obří den gecko | Phelsuma gigas | Rodrigues | Vyhynulý (c. AD 1850) |  Denní gekoni |
Skinks
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Vaillantova mabuya | Chioninia vaillanti | Kapverdy | Ohrožený |  Pevnina mabuyin skinks |
 Kapverdský obří skink | Macroscincus coctei | Kapverdy | Vyhynulý (po inzerátu 1900) | |
 Mauricius obří skink | Leiolopisma mauritiana | Mauricius | Vyhynulý (po inzerátu 1600) | Pevnina eugongyline skinks |
| Teror skink | Phoboscincus bocourti | Desle des Pins vypnuto Nová Kaledonie | Ohrožený |  Pevnina eugongyline skinks |
| Kishinoueův obří skink | Plestiodon kishinouyei | Miyako ostrovy a Yaeyama Islands, Japonsko | Zranitelný |  asijský Plestiodon spp. |
Ještěrky
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| La Palma obří ještěrka | Gallotia auaritae | La Palma | Kriticky ohrožený |  Ještěrky středomořské sandrunner |
 La Gomera obří ještěrka | Gallotia bravoana | Gomera | Kriticky ohrožený | |
 Tenerife obří ještěrka[31] | Gallotia goliath | Tenerife | Vyhynulý (asi 1500 nl) | |
 El Hierro obří ještěrka | Gallotia simonyi | El Hierro | Kriticky ohrožený | |
 Obří ještěrka Gran Canaria | Gallotia stehlini | Gran Canaria | Nejméně obavy |
Hadi
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
| Ostrov Angel de la Guarda skvrnitý chřestýš | Crotalus mitchellii angelensis | Isla Ángel de la Guarda vypnuto Baja California | Nejméně obavy |  Skvrnitý chřestýš |
| Populace hadích pruhovaných Tadanae-jima[32] | Elaphe quadrivirgata | Ostrov Tadanae-jima pryč Tokio | Neznámý |  Japonský pruhovaný had |
 Populace hada ostrovního tygra | Notechis scutatus | Ostrovy Mount Chappell, Williams, Hopkins, a Souostroví Nuyts (jižní Austrálie )[33] | Nejméně obavy[34] |  Tygří had |
| Isla Cerralvo had s dlouhým nosem | Rhinocheilus lecontei etheridgei | Ostrov Jacques Cousteau vypnuto Baja California Sur | Neznámý |  Had s dlouhým nosem |
Pochybné příklady


- The varan komodský z Flores a blízké ostrovy, největší existující ještěrka a podobný (zaniklý) obr monitor ještěrka z Timor byly považovány za příklady obrovských ostrovních masožravců. Vzhledem k tomu, že ostrovy mají tendenci nabízet omezené jídlo a území, jsou jejich savčí šelmy (jsou-li přítomni) obvykle menší než kontinentální. Tyto případy zahrnují ectothermic masožravci na ostrovech příliš malé na to, aby toho hodně podporovali savčí soutěž. Tyto ještěrky však nejsou tak velké jako jejich vyhynulý australský příbuzný megalania, a bylo navrženo na základě fosilních důkazů, že předkové těchto varanidů nejprve vyvinuli svou velkou velikost v Austrálii a poté se rozptýlili do Indonésie.[35] Pokud je to pravda, spíše než na ostrovní obry by byli považováni za příklady fyletický gigantismus. Podpora této interpretace je důkazem ještěrky v pliocénní Indii, Varanus sivalensis, srovnatelné co do velikosti komodoensis.[35] Vzhledem k tomu, že Austrálie je často popisována jako největší ostrov na světě a že příbuzná megalanie, největší suchozemská ještěrka známá ve fosilních záznamech, byla omezena na Austrálii, vnímání největších australských / indonéských ještěrek jako ostrovních obrů může mít stále nějakou platnost.
- Obří želvy v Galapágy a Seychely, největší existující želvy, stejně jako vyhynulé želvy Mascarenes a Kanárské ostrovy, jsou často považovány za příklady ostrovního gigantismu. Během pleistocénu se v něm však vyskytovaly srovnatelně velké nebo větší želvy Austrálie (Meiolania ), jižní Asie (Megalochelys ), Madagaskar (Aldabrachelys ), Severní Amerika[36] (Hesperotestudo ) a Jižní Amerika[37] (Chelonoidis, stejný rod, který se nyní nachází na Galapágách[38]) a na řadě dalších, přístupnějších ostrovů.[36] Na konci pliocénu byli také přítomni v Africe.[39] ("Geochelon " laetoliensis[39]). Současná situace velkých želv, které se nacházejí pouze na vzdálených ostrovech, zřejmě odráží skutečnost, že tyto ostrovy byly objeveny lidmi nedávno a nebyly hustě osídleny, takže jejich želvy méně podléhají nadměrné vykořisťování.
Obojživelníci
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný | Ostrov / pevnina poměr délky nebo hmotnosti |
|---|---|---|---|---|---|
| Obří rosnička Svatého Tomáše | Hyperolius thomensis[40] | Ostrov Svatého Tomáše | Ohrožený |  Africké rákosové žáby | |
| Palmová lesní žába | Leptopelis palmatus[40] | Princip ostrov | Zranitelný |  Červená rosnička | LR ≈ 1,2 [F] |
| Obří Fidži pozemní žába | Platymantis megabotoniviti[43] | Viti Levu, Fidži | Vyhynulý |  asijský platymantiny | |
| Obří tráva žába Svatého Tomáše | Ptychadena newtoni[40] | Ostrov Svatého Tomáše | Ohrožený |  Mascarene tráva žába |
Členovci





















Plži
| Příklad | Binomické jméno | Nativní rozsah | Aktuální stav | Kontinentální příbuzný |
|---|---|---|---|---|
 Kauri přistávají hlemýždi | Paryphanta spp. Powelliphanta spp. | Nový Zéland | Blízko ohrožení |  Jiné rhytididy |
Flóra
Kromě zvětšení velikosti, ostrov tráva rostliny mohou také vykazovat „ostrovní dřevitost“. Nejpozoruhodnější příklady jsou megaherbety z Nový Zéland je subantarktické ostrovy.[47] Zvýšená velikost listů a semen byla hlášena také u některých ostrovních druhů bez ohledu na formu růstu (bylina, keř nebo strom ).[48]




















Viz také
Poznámky
- ^ Snížení predace na ostrovech často také vede k krotitelskému chování druhů ostrovních kořistí, což je trend, který byl analyzován u ještěrek.[2][3]
- ^ Nejdříve známý novozélandský předek kiwi, předpokládaný nedávný příjezd z Austrálie.[20]
- ^ Na základě odhadované celkové délky H. delcourti, ~ 23,6 palce,[28] a průměrná délka člena Diplodactylus, nejvíce speciose rod australských diplodactylid gekonů, ~ 3,5 palce.[29]
- ^ Na základě průměrné celkové délky většího poddruhu R. l. leachianus, ~ 15,5 palce,[30] a průměrná délka člena Diplodactylus, nejvíce speciose rod australských diplodactylid gekonů, ~ 3,5 palce.[29]
- ^ Na základě průměrné hmotnosti většího poddruhu R. l. leachianus, ~ 240 g,[30] s průměrnou hmotností člena Diplodactylus, nejvíce speciose rod australských diplodactylid gekonů, ~ 4 g.[29]
- ^ Na základě průměrné délky ženského čenichu k ventilaci (SVL) L. palmatus, ~ 96 mm,[41] s průměrnou SVL žen L. rufus, ~ 80 mm.[42]
Reference
- ^ Herczeg, G. B .; Gonda, A. L .; Merilä, J. (2009-07-16). „Evoluce gigantismu u devíti ostnatých sticklebacků“. Vývoj. 63 (12): 3190–3200. doi:10.1111 / j.1558-5646.2009.00781.x. PMID 19624722. S2CID 205782326.
- ^ Cooper, W. E .; Pyron, R. A .; Garland, T. (01.01.2014). „Krotkost ostrova: Život na ostrovech snižuje počáteční vzdálenost letu“. Sborník Královské společnosti B: Biologické vědy. 281 (1777): 20133019. doi:10.1098 / rspb.2013.3019. PMC 3896029. PMID 24403345.
- ^ Yong, E. (08.01.2014). "Ostrovy krotí zvířata". Příroda. doi:10.1038 / příroda.2014.14462. S2CID 183158746.
- ^ Jaffe, A.L .; Slater, G. J .; Alfaro, M. E. (2011-01-26). „Vývoj ostrovního gigantismu a kolísání velikosti těla u želv a želv“. Biologické dopisy. 7 (4): 558–561. doi:10.1098 / rsbl.2010.1084. PMC 3130210. PMID 21270022.
- ^ Barahona, F .; Evans, S.E .; Mateo, J. A.; Garcia-Marquez, M .; Lopez-Jurado, L.F. (březen 2000). „Endemismus, gigantismus a vyhynutí u ostrovních ještěrek: rod Gallotia na Kanárských ostrovech “. Journal of Zoology. 250 (3): 373–388. doi:10.1017 / s0952836900003101. hdl:10553/19918.
- ^ A b Raia, P .; Meiri, S. (srpen 2006). „Vládne ostrov u velkých savců: paleontologie se setkává s ekologií“. Vývoj. 60 (8): 1731–1742. doi:10.1111 / j.0014-3820.2006.tb00516.x. PMID 17017072. S2CID 26853128.
- ^ Keehn, J. E.; Nieto, N. C .; Tracy, C. R .; Gienger, C. M .; Feldman, C. R. (2013-08-27). „Evoluce na pustém ostrově: Odchylka velikosti těla mezi plazy nevadského ostrova Anaho a pevninou kolem jezera Pyramid“. Journal of Zoology. 291 (4): 269–278. doi:10.1111 / jzo.12066.
- ^ Lomolino, M. V. (2005-09-05). "Vývoj velikosti těla u ostrovních obratlovců: obecnost ostrovního pravidla". Časopis biogeografie. 32 (10): 1683–1699. doi:10.1111 / j.1365-2699.2005.01314.x. hdl:2027.42/146565.
- ^ Filin, I .; Ziv, Y. (2004). „Nová teorie ostrovní evoluce: sjednocení ztráty dispergovatelnosti a změny tělesné hmotnosti“ (PDF). Evoluční ekologický výzkum. 6: 115–124.
- ^ Turvey, S. T. (2006). „Nový rod a druh obří hutie (Tainotherium valei) z kvartéru Portorika: vyhynulý stromový čtyřnohý? “. Journal of Zoology. 270 (4): 585–594. doi:10.1111 / j.1469-7998.2006.00170.x.
- ^ Torres-Roig, E .; Agustí, J .; Bover, P .; Alcover, J.A. (2017). „Nový obří cricetin z bazálního pliocénu na Mallorce (Baleárské ostrovy, západní Středomoří): biostratigrafický vztah se zónami kontinentálních savců“. Historická biologie. 31 (5): 559–573. doi:10.1080/08912963.2017.1377194. S2CID 135302585.
- ^ Freudenthal, M. (1985). Cricetidae (Rodentia) z neogenu Gargano (Prov. Foggia, Itálie). Rijksmuseum van Geologie en Mineralogie.
- ^ „Studovaly se„ super velké “polní myši sv. Kildy“. BBC. 2010-09-03. Citováno 2020-03-02.
- ^ http://www.mapama.gob.es/es/biodiversidad/temas/conservacion-de-especies-amenazadas/LIRON_tcm7-20977.pdf
- ^ Timm, R. M .; Weijola, V .; Aplin, K. P .; Donnellan, S. C .; Flannery, T. F .; Thomson, V .; Pine, R. H. (2016-04-12). "Nový druh Ratus (Rodentia: Muridae) z ostrova Manus, Papua Nová Guinea ". Journal of Mammalogy. 97 (3): 861–878. doi:10.1093 / jmammal / gyw034.
- ^ http://www.raco.cat/index.php/Orsis/article/viewFile/24434/24268
- ^ Daams, R .; Freudenthal, M. (1985). "Stertomys laticrestatus, nový kluzák (dormice, Rodentia) ze ostrovní fauny Gargano (Prov. Foggia, Itálie) ". Scripta Geologica. 77: 21–27.
- ^ A b MacPhee, R.D.E., Iturralde-Vinent, M.A. a Gaffney, E. S. (Únor 2003). „Domo de Zaza, raně miocénní lokalita obratlovců na jihu střední Kuby, s poznámkami o tektonickém vývoji Portorika a průchodu Mona“. Americké muzeum Novitates. 3394 (1): 1–42. doi:10.1206 / 0003-0082 (2003) 394 <0001: DDZAEM> 2.0.CO; 2. hdl:2246/2820.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ „Pozdně křídová zvířata na rumunském ostrově Haţeg - komplexnější pohled“.
- ^ Hoden, Trevor H .; et al. (2013). Miocénní fosilie ukazují, že kiwi (Apteryx, Apterygidae) pravděpodobně nejsou fyletičtí trpaslíci (PDF). Paleornitologický výzkum 2013, sborník z 8. mezinárodního zasedání Společnosti pro avianskou paleontologii a evoluci. Citováno 16. září 2017.
- ^ Pavia, M .; Meijer, H. J. M .; Rossi, M. A .; Göhlich, U. B. (2017-01-11). "Extrémní ostrovní adaptace Garganornis ballmanni Meijer, 2014: obří Anseriformes neogenu středomořské pánve “. Royal Society Open Science. 4 (1): 160722. Bibcode:2017RSOS .... 460722P. doi:10.1098 / rsos.160722. PMC 5319340. PMID 28280574.
- ^ https://www.canterburymuseum.com/about-us/media-releases/african-origins-for-the-enigmatic-adzebill/
- ^ Naish, Darrene (2008-01-28). „Jestřábi titáni a další super dravci“. Blog zoologie Tetrapod. ScienceBlogs LLC. Citováno 2011-03-02.
- ^ Lerner, Heather R.L .; Mindell, David P. (2005). „Fylogeneze orlů, supů ze Starého světa a dalších Accipitridae na základě nukleární a mitochondriální DNA“. Molekulární fylogenetika a evoluce. 37 (2): 327–346. doi:10.1016 / j.ympev.2005.04.010. PMID 15925523.
- ^ Pregill, G. K.; Steadman, D. W. (březen 2004). „Iguanas v jižním Pacifiku: dopady na člověka a nový druh“. Herpetology Journal. 38 (1): 15–21. doi:10.1670 / 73-03A. JSTOR 1566081. S2CID 85627049.
- ^ Pregill, G. K.; Worthy, T. H. (březen 2003). „New Iguanid Lizard (Squamata, Iguanidae) from the Lare Quaternary of Fiji, Southwest Pacific“. Herpetologica. 59 (1): 57–67. doi:10.1655 / 0018-0831 (2003) 059 [0057: ANILSI] 2.0.CO; 2. ISSN 0018-0831.
- ^ A b Petren, K .; Case, T.J. (1997). „Fylogenetická analýza vývoje velikosti těla a biogeografie u Chuckwallas (Sauromalus) a další leguáni “. Vývoj. 51 (1): 206–219. doi:10.1111 / j.1558-5646.1997.tb02402.x. PMID 28568786. S2CID 22032248.
- ^ Wilson, K.-J. (2004). Let Huia: Ekologie a ochrana novozélandských žab, plazů, ptáků a savců. Canterbury University Press. ISBN 0-908812-52-3. OCLC 937349394.
- ^ A b C Stewart, C. "Diplodactylus Gekoni Austrálie “. reptilesmagazine.com/. Citováno 2020-02-29.
- ^ A b Bergman, J .; Hamper, R. (2016). "List péče o nový gekon Gedon". reptilesmagazine.com/. Citováno 2020-02-29.
- ^ Maca-Meyer, N .; Carranza, S .; Rando, J. C .; Arnold, E. N .; Cabrera, V. M. (01.12.2003). „Stav a vztahy vyhynulého obřího ještěra na Kanárských ostrovech Gallotia goliath (Reptilia: Lacertidae), hodnoceno pomocí staré mtDNA z jejích mumifikovaných pozůstatků “ (PDF). Biologický žurnál společnosti Linnean. 80 (4): 659–670. doi:10.1111 / j.1095-8312.2003.00265.x. Citováno 2010-04-03.
- ^ https://www.jstage.jst.go.jp/article/hsj2000/21/1/21_1_43/_pdf
- ^ Keogh, J. S .; Scott, I. A. W .; Hayes, C. (leden 2005). „Rychlý a opakovaný původ ostrovního gigantismu a nanismu u australských tygřích hadů“. Vývoj. 59 (1): 226–233. doi:10.1111 / j.0014-3820.2005.tb00909.x. PMID 15792242. S2CID 58524.
- ^ Michael, D .; Clemann, N .; Robertson, P. (2018). "Notechis scutatus". Červený seznam ohrožených druhů IUCN. 2018: e.T169687A83767147. Citováno 19. prosince 2019.
- ^ A b Hocknull, S.A .; Piper, P.J .; van den Bergh, G.D .; Due, R.A .; Morwood, M. J.; Kurniawan, I. (2009). „Dragon's Paradise Lost: Palaeobiogeografie, evoluce a vyhynutí dosud největších suchozemských ještěrek (Varanidae)“. PLOS ONE. 4 (9): e7241. Bibcode:2009PLoSO ... 4.7241H. doi:10.1371 / journal.pone.0007241. PMC 2748693. PMID 19789642.
- ^ A b Hansen, D. M .; Donlan, C. J .; Griffiths, C. J .; Campbell, K. J. (duben 2010). „Ekologická historie a latentní potenciál zachování: velké a obří želvy jako model pro substituce taxonů“ (PDF). Ekografie. 33 (2): 272–284. doi:10.1111 / j.1600-0587.2010.06305.x. Citováno 2012-03-02.
- ^ Cione, A. L .; Tonni, E. P .; Soibelzon, L. (2003). „The Broken Zig-Zag: Late Cenozoic large savec and tortoise extinction in South America“ (PDF). Mus. Argentino Cienc. Nat. N.S. 5 (1): 1–19. doi:10.22179 / REVMACN.5.26. ISSN 1514-5158. Archivovány od originál (PDF) dne 06.07.2011. Citováno 2011-02-06.
- ^ Fariña, R.A., Vizcaíno, S.F. & De Iuliis, G. (2013) Megafauna: Obří zvířata Jižní Ameriky. Indiana University Press, 448 stran.
- ^ A b Harrison, T. (2011). "Želvy (Chelonii, Testudinidae)". Paleontologie a geologie Laetoli: Evoluce člověka v kontextu, sv. 2: Fosilní homininy a sdružená fauna. Paleobiologie a paleoantropologie obratlovců. Springer Science + Business Media. 479–503. doi:10.1007/978-90-481-9962-4_17. ISBN 978-90-481-9961-7.
- ^ A b C Measey, G.J .; Vences, M .; Drewes, R.C .; Chiari, Y .; Melo, M .; Bourles, B. (2006). „Sladkovodní cesty přes oceán: molekulární fylogeneze žáby Ptychadena newtoni poskytuje pohled na kolonizaci obojživelníků oceánských ostrovů ". Časopis biogeografie. 34 (1): 7–20. doi:10.1111 / j.1365-2699.2006.01589.x.
- ^ "Leptopelis palmatus". amphibiaweb.org. University of California, Berkeley. 2008. Citováno 29. února 2020.
- ^ "Leptopelis rufus". amphibiaweb.org. University of California, Berkeley. 2008. Citováno 29. února 2020.
- ^ Worthy, T.H. (2001). "Nový druh Platymantis (Anura: Ranidae) z kvartérních ložisek na Viti Levu na Fidži “. Paleontologie. 44 (4): 665–680. doi:10.1111/1475-4983.00197.
- ^ Ani kokosoví kraby, ani jejich příbuzní nemohou plavat za larvální stádium, takže dospělí v praxi přistávají se zvířaty. Krabí kokosové ořechy mohou vážit více než 4 kg (9 liber); největší poustevnický krab příbuzného rodu Coenobita, C. brevimanus pobřežní Afriky a Asie dosahuje pouze 230 gramů (0,5 liber).
- ^ „Akční plán biologické rozmanitosti na ostrově Ascension: Garypus titanius akční plán pro druhy “ (PDF). Georgetown, Ostrov Ascension: Odbor ochrany ostrova Ascension Island. 26. 02. 2015. Citováno 2019-09-11.
- ^ Keppel, Gunnar; Lowe, Andrew J .; Possingham, Hugh P. (2009). „Měnící se pohledy na biogeografii tropického jižního Pacifiku: vlivy rozptýlení, vikariátu a vyhynutí“. Časopis biogeografie. 36 (6): 1035–1054. doi:10.1111 / j.1365-2699.2009.02095.x. ISSN 0305-0270.
- ^ Bowen, Lizabeth; Vuren, Dirk Van (1997). „Ostrovní endemické rostliny postrádají obranu proti býložravcům“. Biologie ochrany. 11 (5): 1249–1254. doi:10.1046 / j.1523-1739.1997.96368.x. ISSN 0888-8892.
- ^ A b C https://www.sciencelearn.org.nz/resources/2289-small-islands-breed-big-seeds
- ^ http://www.terrain.net.nz/friends-of-te-henui-group/table-1/corokia-macrocarpa-chatham-is-korokio.html
- ^ http://www.terrain.net.nz/friends-of-te-henui-group/table-1/korokio.html
- ^ A b Burns, K.C. (Květen 2019). Evoluce v izolaci: Hledání ostrovního syndromu u rostlin. Cambridge University Press. doi:10.1017/9781108379953. ISBN 978-1108379953. OCLC 1105218367.
- ^ Proctor, J. (1984). "Vegetace žulových ostrovů na Seychelách". V Stoddart, D. R. (ed.). Biogeografie a ekologie na Seychelských ostrovech. W. Junk. ISBN 978-90-6193-881-1. OCLC 906429733.
externí odkazy
| Pravidla |
|  |
|---|---|---|
| Příbuzný |
| |