Haploskupina L3 (mtDNA) - Haplogroup L3 (mtDNA)
| Haploskupina L3 | |
|---|---|
| Možná doba vzniku | 80 000–60 000 YBP[1] |
| Možné místo původu | východní Afrika[1][2] nebo Asie[3] |
| Předek | L3'4 |
| Potomci | L3a, L3b'f, L3c'd, L3e'i'k'x, L3h, M, N |
| Definování mutací | 769, 1018, 16311[4] |
Haploskupina L3 je lidská mitochondriální DNA (mtDNA) haploskupina. Clade hrál klíčovou roli v časném rozptýlení anatomicky moderní lidé.
Je silně spojena s migrace mimo Afriku moderních lidí před asi 70–50 000 lety. Zdědí to všechny moderní neafrické populace i některé populace v Africe.[5][6]
Původ

Haploskupina L3 vznikla téměř před 70 000 lety, blízko doby nedávná událost mimo Afriku. Toto rozptýlení vzniklo v východní Afrika a rozšířila se do západní Asie a dále do jižní a jihovýchodní Asie v průběhu několika tisíciletí a některé výzkumy naznačují, že L3 se podílela na této migraci z Afriky. Odhad z roku 2007 pro věk L3 naznačoval rozsah 104–84 000 před lety.[7] Novější analýzy, včetně Soares et al. (2012) přicházejí k novějšímu datu, zhruba před 70–60 000 lety. Soares a kol. také naznačují, že L3 s největší pravděpodobností expandoval z východní Afriky do Eurasie někdy před 65–55 000 lety před lety jako součást nedávné akce mimo Afriku, stejně jako z východní Afriky do střední Afriky před 60–35 000 lety.[1]V roce 2016 Soares et al. opět naznačil, že haploskupina L3 se objevila ve východní Africe, což vedlo k migraci mimo Afriku, asi před 70-60 000 lety.[8]
Haploskupiny L6 a L4 tvoří sesterské subtypy L3, které vznikly ve východní Africe zhruba ve stejnou dobu, ale které se neúčastnily migrace mimo Afriku. Klady předků L3'4'6 byly odhadnuty na zhruba 110 kya a L3'4 clade na 95 kya.[5]


A: Výstup z předchůdce L3 do Eurasie. b: Návrat do Afriky a expanze do Asie bazálních linií L3 s následnou diferenciací na obou kontinentech.
Možnost vzniku L3 již v Asii navrhli také Cabrera et al. (2018) na základě podobných dat splynutí L3 a jeho euroasijské distribuce M a N derivátové subtypy (asi 70 kya), vzdálená lokace v Jihovýchodní Asie nejstarších subclades M a N a srovnatelný věk otcovské haploskupiny DE. Podle této hypotézy by po počáteční migraci nositelů pre-L3 (L3'4 *) kolem 125 kya mimo Afriku došlo k zpětné migraci žen nesoucích L3 z Eurasie do východní Afriky někdy po 70 kya. Navrhuje se, aby tato zpětná migrace byla v souladu s nositeli otcovské haploskupiny E, o kterém se rovněž předpokládá, že pochází z Eurasie. Tyto nové euroasijské linie se poté navrhují tak, aby do značné míry nahradily staré autochtonní mužské a ženské severovýchodní africké linie.[3]
Podle jiného výzkumu, i když došlo k dřívější migraci anatomicky moderních lidí z Afriky, současné euroasijské populace místo toho sestupují z pozdější migrace z Afriky datované před asi 65 000 až 50 000 lety (spojená s migrací z L3).[9][2][10] Vai a kol. (2019) naznačují, z nově objevené staré a hluboce zakořeněné větve mateřské haploskupiny N nalezené v počátcích neolitických severoafrických pozůstatků, že haploskupina L3 vznikla ve východní Africe před 70 000 - 60 000 lety a obě se šířily v Africe a opustily Afriku jako část migrace mimo Afriku, přičemž haploskupina N se od ní odchýlila brzy poté (před 65 000–50 000 lety) buď v Arábii nebo případně v severní Africe, a haploskupina M pocházející ze Středního východu přibližně ve stejnou dobu jako N.[2]
Studie Lipsona a kol. (2019) analyzující pozůstatky z kamerunského webu Shum Laka shledali, že jsou více podobné dnešní době Trpasličí národy než západoafričanům, a naznačuje, že několik dalších skupin (včetně společných předků západoafrických, východoafrických a předků neafričanů) běžně pocházejících z lidské populace pocházející z východní Afriky přibližně před 80 000–60 000 lety, kterou navrhnout byl také zdroj a zóna původu haploskupiny L3 asi před 70 000 lety.[11]
Rozdělení
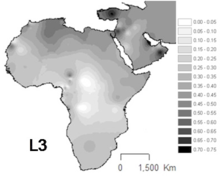
L3 je běžný v Severovýchodní Afrika a některé další části východní Afriky,[12] na rozdíl od jiných částí Afriky, kde jsou haploskupiny L1 a L2 představují přibližně dvě třetiny linií mtDNA.[13] L3 subleages jsou také časté v Arabský poloostrov.
L3 je rozdělena do několika subtypů, z nichž dva vytvořily makrohaploskupiny M a N které dnes nosí většina lidí mimo Afriku.[13] Existuje alespoň jeden relativně hluboký non-M, non-N clade L3 mimo Afriku, L3f1b6, který se vyskytuje s frekvencí 1% v Asturie, Španělsko. To se lišilo od afrických linií L3 nejméně před 10 000 lety.[14]
Podle Maca-Meyer et al. (2001), „L3 souvisí více s euroasijský haploskupiny než nejrozmanitější Afričan shluky L1 a L2 ".[15] L3 je haploskupina, z níž pocházejí všichni moderní lidé mimo Afriku.[16] Existuje však větší rozmanitost hlavních větví L3 v Africe než mimo ni, dvě hlavní neafrické větve jsou L3 odnože M a N.
Distribuce subclade

L3 má sedm potomků ve stejné vzdálenosti: L3a, L3b'f, L3c'd, L3e'i'k'x, L3h, M, N. Pět je afrických, zatímco dva jsou spojeni s Z Afriky událost.
- N – Eurasie a části Afriky kvůli zpětné migraci.[6][17]
- M – Asie, Středomořská pánev a části Afriky kvůli zpětné migraci.[6][17]
- L3a - Východní Afrika.[5][6] Střední až vysoké frekvence nalezené mezi Sanye, Samburu, Iraqw, Yaaku, El-Molo a další menší domorodé populace z Východoafrické příkopové propadliny. V Súdánu a v oblasti Sahelu se vyskytuje jen zřídka nebo vůbec.[18]
- L3a1 - Nalezeno ve východní Africe. Odhadovaný věk 35,8–39,3 ka.[6]
- L3a2 - Nalezeno ve východní Africe. Odhadovaný věk 48,3–57,7 ka.[19][Poznámka 1]
- L3b'f
- L3b - Šíření z východní Afriky v horní paleolit do západní a střední Afriky. Některé subclades se šířil ze střední Afriky do východní Afriky s migrací Bantu.[6]
- L3f - Severovýchodní Afrika, Sahel, Arabský poloostrov, Iberia. Gaalien,[21] Bejo[21]
- L3f1
- L3f2 - Primárně distribuováno ve východní Africe.[6] Nalezeno také v severní Africe a střední Africe.[20]
- L3f3 - Šíření z východní Afriky do Čadu a Sahelu kolem 8–9 ka.[6] Nalezeno v Čadská pánev.[20][22]
- L3c'd
- L3c - Extrémně vzácná linie s pouhými dvěma vzorky dosud nalezenými ve východní Africe a Blízký východ.[6]
- L3d - Šíření z východní Afriky v horní paleolit do střední Afriky. Některé subclades se rozšířily do východní Afriky s migrací Bantu.[6] Nalezeno mezi Fulani,[5] Chadians,[5] Etiopané,[23] Akanští lidé,[24] Mosambik,[23] Jemenčané,[23] Egypťané, Berbeři[25]
- L3e'i'k'x
- L3e - Šíření z východní Afriky v horní paleolit do západní a střední Afriky. Je to nejběžnější sub-clade L3 Bantu - mluvící populace.[26] L3e je spojován se středoafrickým původem a je také nejčastější subclade L3 mezi afroameričany, Afro-Brazilci a Karibik[27]
- L3e1 - Šíření ze západní a střední Afriky do jihozápadní Afriky s migrací Bantu. Nalezen v Angola (6.8%).[28] Mosambik, súdánský a Kikuyu z Keňa stejně jako v Jemenu a mezi Akanští lidé[24]
- L3e5 - Vznikl v čadské pánvi. Nalezen v Alžírsko,[29] stejně jako Burkina Faso, Nigérie, Jih Tunisko, Jih Maroko a Egypt[30]
- L3i Téměř výlučně ve východní Africe.[6]
- L3k - Vzácná haploskupina vyskytující se především v severní Africe a na Sahelu.[6][20]
- L3x - Téměř výlučně ve východní Africe.[6] Nalezeno mezi etiopskými Oromos,[23] Egypťané[Poznámka 2][31]
- L3e - Šíření z východní Afriky v horní paleolit do západní a střední Afriky. Je to nejběžnější sub-clade L3 Bantu - mluvící populace.[26] L3e je spojován se středoafrickým původem a je také nejčastější subclade L3 mezi afroameričany, Afro-Brazilci a Karibik[27]
- L3h - Téměř výlučně ve východní Africe.[6]
- L3h1 - Primárně se vyskytuje ve východní Africe s větvemi L3h1b1 sporadicky nalezenými v Sahelu a severní Africe.[19][20]
- L3h2 - Nalezeno v severovýchodní Africe a Sokotře. Rozděleny od ostatních větví L3h již v 65–69 ka během roku střední paleolit.[19][20]
Starověké a historické vzorky
Haploskupina L3 byla pozorována ve starověké fosilii patřící k Pre-Pottery Neolithic B kultura.[32] L3x2a byla pozorována u 4500 let staré lovecké skupiny vykopané v Mota, Etiopie, se starou fosílií, která byla nejvíce spojena s moderní Jihozápadní etiopský populace.[33][34] Haploskupina L3 byla také nalezena mezi staroegyptský mumie (1/90; 1%) vyhloubené v Abusir el-Meleq archeologické naleziště ve středním Egyptě, zbytek pochází z euroasijských subclades, které pocházejí z před-Ptolemaic /pozdě Nová říše a Ptolemaiova období. Staroegyptské mumie nesly genomickou složku blízkovýchodního původu, která nejvíce souvisí s moderními blízkovýchodníky.[35] Navíc byla ve starověku pozorována haploskupina L3 Guanche fosilie vytěžené v Gran Canaria a Tenerife na Kanárské ostrovy, které byly radiokarbonem do 7. až 11. století n. l. Všichni jedinci nesoucí clade byli obýváni na místě Gran Canaria, přičemž u většiny těchto vzorků bylo zjištěno, že patří do subclade L3b1a (3/4; 75%), zbytek pochází z obou ostrovů (8/11; 72%) z euroasijských subclades. Guancheovy kostry také nesly autochtonní maghrebskou genomickou složku, která vrcholí mezi moderními Berbeři, což naznačuje, že pocházejí z předků berberských populací obývajících severozápadní Affoundnat ve vysoké míře[36]
Ve starověkých pozůstatcích spojených s pastoračním neolitem a pastorační dobou železa východní Afriky byla odkryta celá řada L3.[37]
| Kultura | Genetický shluk nebo afinita | Země | Stránky | datum | Mateřská haploskupina | Otcovská haploskupina | Zdroj |
| Brzy pastorační | PN | Keňa | Prettejohnova rokle (GsJi11) | 4060–3860 | L3f1b | – | Prendergast 2019 |
| Pastorační neolit | PN | Keňa | Coleův pohřeb (GrJj5a) | 3350–3180 | L3i2 | E-V32 | Prendergast 2019 |
| Pastorační neolit nebo elmenteitán | PN | Keňa | Jeskyně Rigo (GrJh3) | 2710–2380 | L3f | E-M293 | Prendergast 2019 |
| Pastorační neolit | PN | Keňa | Naishi Rockshelter | 2750–2500 | L3x1a | E-V1515 (prob. E-M293) | Prendergast 2019 |
| Pastorační neolit | PN | Tanzanie | Jeskyně Gishimangeda | 2490–2350 | L3x1 | – | Prendergast 2019 |
| Pastorační neolit | PN | Keňa | Pohřebiště Naivasha | 2350–2210 | L3h1a1 | E-M293 | Prendergast 2019 |
| Pastorační neolit | PN | Keňa | Pohřebiště Naivasha | 2320–2150 | L3x1a | E-M293 | Prendergast 2019 |
| Pastorační neolit | PN | Tanzanie | Jeskyně Gishimangeda | 2150–2020 | L3i2 | E-M293 | Prendergast 2019 |
| Pastorační neolit nebo elmenteitán | PN | Keňa | Jeskyně Njoro River II | 2110–1930 | L3h1a2a1 | – | Prendergast 2019 |
| Pastorační neolit | N / A | Tanzanie | Jeskyně Gishimangeda | 2000–1900 | L3h1a2a1 | – | Prendergast 2019 |
| Pastorační neolit | PN | Keňa | Ol Kalou | 1810–1620 | L3d1d | E-M293 | Prendergast 2019 |
| Pastorační doba železná | PIA | Keňa | Kisima Farm, C4 | 1060–940 | L3h1a1 | E-M75 (kromě M98) | Prendergast 2019 |
| Pastorační doba železná | PIA | Keňa | Polokošile Emurua Ole (GvJh122) | 420–160 | L3h1a1 | E-M293 | Prendergast 2019 |
| Pastorační doba železná | Odlehlá hodnota PN | Keňa | Kokurmatakore | N / A | L3a2a | E-M35 (ne E-M293) | Prendergast 2019 |
Strom
Tento fylogenetický strom haploskupiny L3 subclades vychází z příspěvku Mannisa van Oven a Manfreda Kaysera Aktualizovaný komplexní fylogenetický strom globální variace lidské mitochondriální DNA[4] a následný publikovaný výzkum.[38]
Nejnovější společný předek (MRCA)
- L1-6
- L2-6
- L2'3'4'6
- L3'4'6
- L3'4
- L3
- L3a
- L3a1
- L3a1a
- L3a1b
- L3a2
- L3a2a
- L3a1
- L3b'f
- L3b
- L3b1
- L3b1a
- L3b1a1
- L3b1a2
- L3b1a3
- L3b1a4
- L3b1a5
- L3b1a5a
- L3b1a6
- L3b1a7
- L3b1a7
- L3b1a8
- L3b1a9
- L3b1a9a
- L3b1a10
- L3b1a11
- L3b1b
- L3b1b1
- L3b1a
- L3b2
- L3b2a
- L3b2a
- L3b3
- L3b1
- L3f
- L3f1
- L3f1a
- L3f1a1
- L3f1b
- L3f1b1
- L3f1b2
- L3f1b2a
- L3f1b3
- L3f1b4
- L3f1b4a
- L3f1b4a1
- L3f1b4b
- L3f1b4c
- L3f1b4a
- L3f1b5
- L3f1a
- L3f2
- L3f2a
- L3f2b
- L3f3
- L3f3a
- L3f3b
- L3f1
- L3b
- L3c'd
- L3c
- L3d
- L3d1-5
- L3d1
- L3d1a
- L3d1a1
- L3d1a1a
- L3d1a1
- L3d1b
- L3d1b1
- L3d1c
- L3d1d
- L3d1a
- 199
- L3d2
- L3d5
- L3d3
- L3d3a
- L3d4
- L3d5
- L3d1
- L3d1-5
- L3e'i'k'x
- L3e
- L3e1
- L3e1a
- L3e1a1
- L3e1a1a
- 152
- L3e1a2
- L3e1a3
- L3e1a1
- L3e1b
- L3e1c
- L3e1d
- L3e1e
- L3e1a
- L3e2
- L3e2a
- L3e2a1
- L3e2a1a
- L3e2a1b
- L3e2a1b1
- L3e2a1
- L3e2b
- L3e2b1
- L3e2b1a
- L3e2b2
- L3e2b3
- L3e2b1
- L3e2a
- L3e3'4'5
- L3e3'4
- L3e3
- L3e3a
- L3e3b
- L3e3b1
- L3e4
- L3e3
- L3e5
- L3e3'4
- L3e1
- L3i
- L3i1
- L3i1a
- L3i1b
- L3i2
- L3i1
- L3k
- L3k1
- L3x
- L3x1
- L3x1a
- L3x1a1
- L3x1a2
- L3x1b
- L3x1a
- L3x2
- L3x2a
- L3x2a1
- L3x2a1a
- L3x2a1
- L3x2b
- L3x2a
- L3x1
- L3e
- L3h
- L3h1
- L3h1a
- L3h1a1
- L3h1a2
- L3h1a2a
- L3h1a2b
- L3h1b
- L3h1b1
- L3h1b1a
- L3h1b1a1
- L3h1b1a
- L3h1b2
- L3h1b1
- L3h1a
- L3h2
- L3h1
- M
- N
- L3a
- L3
- L3'4
- L3'4'6
- L2'3'4'6
- L2-6
Viz také
Fylogenetický strom haploskupiny lidské mitochondriální DNA (mtDNA) | |||||||||||||||||||||||||||||||||||||||
| Mitochondriální Eva (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | Ó | A | S | R | Já | Ž | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | před JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K. | |||||||||||||||||||||||||||||||||||||
| H | PROTI | J | T | ||||||||||||||||||||||||||||||||||||
Reference
- ^ A b C Soares, P; Alshamali, F; Pereira, J. B; Fernandes, V; Silva, N.M .; Afonso, C; Costa, M. D; Musilova, E; MacAulay, V; Richards, M. B; Černý, V; Pereira, L (2011). „Expanze mtDNA Haplogroup L3 v Africe i mimo ni“. Molekulární biologie a evoluce. 29 (3): 915–927. doi:10,1093 / molbev / msr245. PMID 22096215.Bodový odhad 71,6 kya podle Soares et al. (2009), ze 70,2 autorů Fernandes et al. (2015).
- ^ A b C d Vai S, Sarno S, Lari M, Luiselli D, Manzi G, Gallinaro M, Mataich S, Hübner A, Modi A, Pilli E, Tafuri MA, Caramelli D, di Lernia S (březen 2019). „Rodová mitochondriální linie N z neolitické„ zelené “Sahary“. Sci Rep. 9 (1): 3530. Bibcode:2019NatSR ... 9,3530V. doi:10.1038 / s41598-019-39802-1. PMC 6401177. PMID 30837540.
- ^ A b C Cabrera VM, Marrero P, Abu-Amero KK, Larruga JM (červen 2018). „Nositelé mitochondriální DNA makrohaploskupiny L3 bazálních linií migrovali zpět do Afriky z Asie asi před 70 000 lety“. BMC Evoluční biologie. 18 (1): 98. doi:10.1186 / s12862-018-1211-4. PMC 6009813. PMID 29921229.
- ^ A b Van Oven, Mannis; Kayser, Manfred (2009). "Aktualizovaný komplexní fylogenetický strom globální variace lidské mitochondriální DNA". Lidská mutace. 30 (2): E386–94. doi:10.1002 / humu.20921. PMID 18853457. S2CID 27566749.
- ^ A b C d E Behar, Doron M .; Villems, Richard; Soodyall, Himla; Blue-Smith, Jason; Pereira, Luisa; Metspalu, Ene; Scozzari, Rosaria; Makkan, Heeran; et al. (2008). „Úsvit lidské matrilineální rozmanitosti“ (PDF). American Journal of Human Genetics. 82 (5): 1130–40. doi:10.1016 / j.ajhg.2008.04.002. PMC 2427203. PMID 18439549.
- ^ A b C d E F G h i j k l m n Ó p q r s t Soares, P .; Alshamali, F .; Pereira, J. B .; Fernandes, V .; Silva, N.M .; Afonso, C .; Costa, M. D .; Musilova, E .; Macaulay, V. (2011-11-16). "Expanze mtDNA Haplogroup L3 v Africe i mimo ni". Molekulární biologie a evoluce. 29 (3): 915–927. CiteSeerX 10.1.1.923.345. doi:10,1093 / molbev / msr245. ISSN 0737-4038. PMID 22096215.
- ^ Gonder, M. K .; Mortensen, H. M .; Reed, F. A .; De Sousa, A .; Tishkoff, S.A. (2006). „Analýza sekvence genomu celé mtDNA staroafrických linií“. Molekulární biologie a evoluce. 24 (3): 757–68. doi:10.1093 / molbev / msl209. PMID 17194802.
- ^ Soares P, Rito T, Pereira L, Richards M (březen 2016). „Genetická perspektiva afrického pravěku“ (PDF). Afrika od MIS 6-2. Paleobiologie a paleoantropologie obratlovců (1): 383–405. doi:10.1007/978-94-017-7520-5_18. ISBN 978-94-017-7519-9.
- ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, Valentin F, Thevenet C, Furtwängler A, Wißing C, Francken M, Malina M, Bolus M, Lari M, Gigli E, Capecchi G, Crevecoeur I , Beauval C, Flas D, Germonpré M, van der Plicht J, Cottiaux R, Gély B, Ronchitelli A, Wehrberger K, Grigorescu D, Svoboda J, Semal P, Caramelli D, Bocherens H, Harvati K, Conard NJ, Haak W , Powell A, Krause J (2016). „Pleistocénní mitochondriální genomy naznačují jediný zásadní rozptyl Afričanů a pozdní obrat ledové populace v Evropě“. Aktuální biologie. 26 (6): 827–833. doi:10.1016 / j.cub.2016.01.037. hdl:2440/114930. PMID 26853362. S2CID 140098861.
- ^ Haber M, Jones AL, Connel BA, Asan, Arciero E, Huanming Y, Thomas MG, Xue Y, Tyler-Smith C (červen 2019). „Vzácná hluboce zakořeněná africká Y-chromozomální haploskupina D0 a její důsledky pro expanzi moderních lidí z Afriky“. Genetika. 212 (4): 1421–1428. doi:10.1534 / genetika.119.302368. PMC 6707464. PMID 31196864.
- ^ Ancient Human DNA from Shum Laka (Cameroon) in the Context of African Population History, autorem Lipson Mark a kol., 2019
- ^ Martina Kujanová; Luisa Pereira; Veronica Fernandes; Joana B. Pereira; Viktor Černý (2009). „Blízkovýchodní neolitický genetický vstup v malé oáze egyptské západní pouště“. American Journal of Physical Anthropology. 140 (2): 336–46. doi:10.1002 / ajpa.21078. PMID 19425100.
- ^ A b Wallace, D; Brown, MD; Lott, MT (1999). "Variace mitochondriální DNA v evoluci a nemoci člověka". Gen. 238 (1): 211–30. doi:10.1016 / S0378-1119 (99) 00295-4. PMID 10570998.
- ^ A b Pardiñas, AF; Martínez, JL; Roca, A; García-Vazquez, E; López, B (2014). „Přes písek a daleko: Interpretace iberské mitochondriální linie se starodávným západoafrickým původem“. Dopoledne. J. Hum. Biol. 26 (6): 777–83. doi:10.1002 / ajhb.22601. PMID 25130626. S2CID 22184219.
- ^ Maca-Meyer, Nicole; González, Ana M; Larruga, José M; Flores, Carlos; Cabrera, Vicente M (2001). „Hlavní genomové mitochondriální linie určují časné lidské expanze“. Genetika BMC. 2: 13. doi:10.1186/1471-2156-2-13. PMC 55343. PMID 11553319.
- ^ „Archivovaná kopie“. Archivovány od originál dne 8.7.2011. Citováno 2009-03-09.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ A b Vicente M. Cabrera (2017). „Nosiče základních linií makrohaploskupiny L3 mitochondriální DNA migrovaly zpět do Afriky z Asie před asi 70 000 lety“. bioRxiv 10.1101/233502.CS1 maint: používá parametr autoři (odkaz)
- ^ Boru, Hirbo, Jibril (2011). „Komplexní genetická historie východoafrických lidských populací“ (PDF): 118. Citovat deník vyžaduje
| deník =(Pomoc) - ^ A b C d E Soares, P .; Alshamali, F .; Pereira, J. B .; Fernandes, V .; Silva, N.M .; Afonso, C .; Costa, M. D .; Musilova, E .; Macaulay, V .; Richards, M. B .; Černý, V .; Pereira, L. (16. listopadu 2011). „Expanze mtDNA Haplogroup L3 v Africe i mimo ni“. Molekulární biologie a evoluce. 29 (3): 915–927. doi:10,1093 / molbev / msr245. PMID 22096215.CS1 maint: ref = harv (odkaz) Doplňující údaje na [1]
- ^ A b C d E F G h i Hernández, Candela L; Soares, Pedro; Dugoujon, Jean M; Novelletto, Andrea; Rodríguez, Juan N; Rito, Teresa; Oliveira, Marisa; Melhaoui, Mohammed; Baali, Abdellatif; Pereira, Luisa; Calderón, Rosario (2015). „Rané holocenní a historické africké podpisy mtDNA na Pyrenejském poloostrově: Andaluský region jako paradigma“. PLOS ONE. 10 (10): e0139784. Bibcode:2015PLoSO..1039784H. doi:10.1371 / journal.pone.0139784. PMC 4624789. PMID 26509580. Doplňující údaje doi:10.1371 / journal.pone.0139784.s006.
- ^ A b Mohamed, Hisham Yousif Hassan. „Genetické vzorce variace chromozomu Y a mitochondriální DNA s důsledky pro osídlení Súdánu“ (PDF). Univerzita v Chartúmu. Citováno 14. června 2016.
- ^ Černý, Viktor; Fernandes, Verónica; Costa, Marta D; Hájek, Martin; Mulligan, Connie J; Pereira, Luísa (2009). „Migrace čadičsky mluvících pastevců v Africe na základě populační struktury čadské pánve a fylogeografie mitochondriální haploskupiny L3f“. BMC Evoluční biologie. 9: 63. doi:10.1186/1471-2148-9-63. PMC 2680838. PMID 19309521.
- ^ A b C d Kivisild, T; Reidla, M; Metspalu, E; Rosa, A; Brehm, A; Pennarun, E; Parik, J; Geberhiwot, T; et al. (2004). „Dědictví etiopské mitochondriální DNA: sledování toku genů napříč bránou slz“. American Journal of Human Genetics. 75 (5): 752–70. doi:10.1086/425161. PMC 1182106. PMID 15457403.
- ^ A b Fendt, Liane; Röck, Alexander; Zimmermann, Bettina; Bodner, Martin; Thye, Thorsten; Tschentscher, Frank; Owusu-Dabo, Ellis; Göbel, Tanja M.K .; Schneider, Peter M .; Parson, Walther (2012). „MtDNA rozmanitost Ghany: forenzní a fylogeografický pohled“. Forensic Science International: Genetics. 6 (2): 244–49. doi:10.1016 / j.fsigen.2011.05.011. PMC 3314991. PMID 21723214.
- ^ List1 - PLOS Patogeny
- ^ Anderson, S. 2006, Fylogenetická a fylogeografická analýza variace africké mitochondriální DNA. Archivováno 10.09.2011 na Wayback Machine
- ^ Bandelt, HJ; Alves-Silva, J; Guimarães, PE; Santos, MS; Brehm, A; Pereira, L; Coppa, A; Larruga, JM; et al. (2001). „Fylogeografie lidské mitochondriální haploskupiny L3e: snímek afrického prehistorie a obchodu s otroky v Atlantiku“. Annals of Human Genetics. 65 (Pt 6): 549–63. doi:10.1046 / j.1469-1809.2001.6560549.x. PMID 11851985. S2CID 221411246.
- ^ Plaza, Stéphanie; Salas, Antonio; Calafell, Francesc; Corte-Real, Francisco; Bertranpetit, Jaume; Carracedo, Ángel; Comas, David (2004). „Pohledy na západní rozptýlení Bantu: analýza linie mtDNA v Angole“. Genetika člověka. 115 (5): 439–47. doi:10.1007 / s00439-004-1164-0. PMID 15340834. S2CID 13213447.
- ^ Asmahan Bekada; Lara R. Arauna; Tahria Deba; Francesc Calafell; Soraya Benhamamouch; David Comas (24. září 2015). „Genetická heterogenita u alžírských lidských populací“. PLOS ONE. 10 (9): e0138453. Bibcode:2015PLoSO..1038453B. doi:10.1371 / journal.pone.0138453. PMC 4581715. PMID 26402429.; Tabulka S5
- ^ Fadhlaoui-Zid, K .; Plaza, S .; Calafell, F .; Ben Amor, M .; Comas, D .; Bennamar, A .; Gaaied, El (2004). "Heterogenita mitochondriální DNA v tuniských Berbers". Annals of Human Genetics. 68 (Pt 3): 222–33. doi:10.1046 / j.1529-8817.2004.00096.x. PMID 15180702. S2CID 6407058.
- ^ Stevanovitch, A .; Gilles, A .; Bouzaid, E .; Kefi, R .; Paris, F .; Gayraud, R. P .; Spadoni, J. L .; El-Chenawi, F .; Beraud-Colomb, E. (2004). „Mitochondriální DNA sekvenční rozmanitost v sedavé populaci z Egypta“. Annals of Human Genetics. 68 (Pt 1): 23–39. doi:10.1046 / j.1529-8817.2003.00057.x. PMID 14748828. S2CID 44901197.
- ^ Fernández, Eva; et al. (2014). „Starověká analýza DNA z roku 8000 př. N.l. poblíž východních zemědělců podporuje ranou neolitickou průkopnickou námořní kolonizaci kontinentální Evropy přes Kypr a Egejské ostrovy“. Genetika PLOS. 10 (6): e1004401. doi:10.1371 / journal.pgen.1004401. PMC 4046922. PMID 24901650.
- ^ Vidět doplňkové materiály z Llorente, M. Gallego; Jones, E. R .; Eriksson, A .; Siska, V .; Arthur, K. W .; Arthur, J. W .; Curtis, M. C .; Stock, J. T .; Coltorti, M .; Pieruccini, P .; Stretton, S .; Brock, F .; Higham, T .; Park, Y .; Hofreiter, M .; Bradley, D. G .; Bhak, J .; Pinhasi, R .; Manica, A. (13. listopadu 2015). „Starověký etiopský genom odhaluje rozsáhlou euroasijskou příměs ve východní Africe“. Věda. 350 (6262): 820–822. Bibcode:2015Sci ... 350..820L. doi:10.1126 / science.aad2879. PMID 26449472.
- ^ Llorente, M. Gallego; Jones, E. R .; Eriksson, A .; Siska, V .; Arthur, K. W .; Arthur, J. W .; Curtis, M. C .; Stock, J. T .; Coltorti, M. (2015-11-13). „Starověký etiopský genom odhaluje rozsáhlou euroasijskou příměs ve východní Africe“. Věda. 350 (6262): 820–822. Bibcode:2015Sci ... 350..820L. doi:10.1126 / science.aad2879. PMID 26449472.
- ^ Schuenemann, Verena J .; et al. (2017). „Staroegyptské genomy mumií naznačují nárůst předků subsaharské Afriky v post-římských obdobích“. Příroda komunikace. 8: 15694. Bibcode:2017NatCo ... 815694S. doi:10.1038 / ncomms15694. PMC 5459999. PMID 28556824.
- ^ Rodríguez-Varela; et al. (2017). „Genomické analýzy předevropského dobytí pozůstatků lidí z Kanárských ostrovů odhalují blízkou afinitu k moderním severoafrickým lidem“. Aktuální biologie. 27 (1–7): 3396–3402.e5. doi:10.1016 / j.cub.2017.09.059. PMID 29107554. Citováno 27. října 2017.
- ^ Prendergast, Mary E .; Lipson, Mark; Sawchuk, Elizabeth A .; Olalde, Iñigo; Ogola, Christine A .; Rohland, Nadin; Sirak, Kendra A .; Adamski, Nicole; Bernardos, Rebecca (2019-05-30). „Starověká DNA odhaluje vícestupňové šíření prvních pastevců do subsaharské Afriky“. Věda. 365 (6448): eaaw6275. Bibcode:2019Sci ... 365.6275P. doi:10.1126 / science.aaw6275. ISSN 0036-8075. PMC 6827346. PMID 31147405.
- ^ "PhyloTree.org | strom | L3". phylotree.org. Citováno 2018-06-25.
Poznámky
- ^ Viz Supplemental_TreeUpdatedOctober.xls nalezený pod doplňkovými daty z Soares a kol. 2011
- ^ GUR46 v tabulce 1. je mtDNA haploskupina L3x2a.
externí odkazy
- Všeobecné
- Iana Logana Stránky mitochondriální DNA
- Haploskupina L3
- Mannis van Oven's PhyloTree.org - podstrom mtDNA L3
- Šíření haploskupiny L3, z národní geografie