ESCRT - ESCRT - Wikipedia
The endozomální třídicí komplexy potřebné pro přepravu (ESCRT) strojní zařízení je tvořeno cytosolický proteinové komplexy, známé jako ESCRT-0, ESCRT-I, ESCRT-II a ESCRT-III. Spolu s řadou doplňkových proteinů umožňují tyto komplexy ESCRT jedinečný režim membrána remodelace, která má za následek ohýbání / pučení membrán od cytoplazma.[1][2] Tyto složky ESCRT byly izolovány a studovány u řady organismů včetně droždí a lidé.[3]
Strojní zařízení ESCRT hraje zásadní roli v řadě buněčných procesů včetně multivezikulární tělo (MVB) biogeneze, buněčná abscise, a virové pučení. Biogeneze multivesikulárního těla (MVB) je proces, při kterém ubikvitin označené proteiny vstupují organely nazývané endosomy tvorbou vezikuly. Tento proces je nezbytný pro to, aby buňky zničily špatně složené a poškozené proteiny.[4] Bez strojů ESCRT se tyto proteiny mohou hromadit a vést k nim neurodegenerativní choroba. Například abnormality ve složkách ESCRT-III mohou vést k neurologickým poruchám, jako je dědičná spastická paraplegie (HSP).[5] Buněčná abcise, proces, při kterém se štěpí membrána spojující dvě dceřiné buňky, je také zprostředkována strojem ESCRT. Bez komplexů ESCRT se dceřiné buňky nemohly oddělit a abnormální buňky obsahující dvojnásobné množství DNA bude vygenerováno. Tyto buňky by byly nevyhnutelně zničeny procesem známým jako apoptóza. A konečně, virové pučení nebo proces, kterým specifické typy virů opouštějí buňky, se nemusí vyskytnout bez absence mechanismu ESCRT. To by nevyhnutelně zabránilo šíření virů z buňky do buňky.
Komplexy ESCRT a doplňkové proteiny

Každý z komplexů ESCRT a doplňkových proteinů má jedinečné struktury, které umožňují odlišné biochemické funkce. Existuje řada synonym pro každou proteinovou složku stroje ESCRT, a to jak pro kvasinky, tak pro metazoans. Souhrnná tabulka všech těchto proteinů je uvedena níže.
V kvasinkách existují následující komplexy / doplňkové proteiny takto:
ESCRT-0
Komplex ESCRT-0 hraje zásadní roli při generování multivesikulárních těl tím, že váže a shlukuje ubikvitinované proteiny a / nebo receptory na povrchu buňky. Komplex je pak odpovědný za vazbu na a lipid na endosomální membráně, která rekrutuje tyto označené proteiny do endosomu.[6] Jednou správně lokalizovaný, tyto proteiny jsou poté přijímány do endosomu přes vezikuly, tvořící multivesikulární těla, a jsou nakonec dodány do lysozom kde jsou degradovány. Tento proces je nezbytný, protože je hlavní cestou degradace poškozených proteinů, které prošly Golgi.[4] Komponenty komplexu ESCRT-0 existují následovně:
Komplex je v poměru 1: 1 heterodimer z Vps27 (vakuolární třídění proteinů protein 27) a Hse1.[1][5] Vps27 a Hse1 dimerizovat přes antiparalelní svinutá cívka GAT (tak pojmenované podle proteinů GGA a Tom1) domén.[1] Vps27 i Hse1 obsahují amino-terminál Doména VHS (tak pojmenovaný, protože je obsažen v PROTIps27, HRS a STAM proteiny).[7] Tyto VHS domény váží ubikvitin na proteiny, jejichž cílem je buňka degradovat. Ubikvitin se může také spojit s motivy interagujícími s ubikvitinem, jako je motiv na Hse1 nebo oboustranná doména nacházející se na Vps27. A Doména FYVE (pojmenovaný po čtyřech proteinech, ve kterých byl původně identifikován: Fab1p, YOTB, Vac1 a EEA1) se nalézá v sendviči mezi doménami motivů Vps27 interagujících s VHS a ubikvitinem.[5][8] Fosfatidylinositol 3-fosfát, běžný endozomální lipid, se váže na tuto FYVE doménu, což vede k náboru ESCRT-0 do endosomu.[5]
ESCRT-I
Úlohou komplexu ESCRT-I je pomáhat při generování multivesikulárních tělísek shlukováním ubikvitinovaných proteinů a působit jako most mezi komplexy ESCRT-0 a ESCRT-II.[9] Hraje také roli v rozpoznávání a remodelaci membrány během úniku membrány tvorbou prstenců na obou stranách střední část těla dělících se buněk. ESCRT-I je také zodpovědný za nábor ESCRT-III, který tvoří zúženou zónu těsně před oddělením buněk.[10] Kromě toho hraje ESCRT-I roli ve virovém pučení interakcí se specifickými virovými proteiny, což vede k náboru dalšího mechanismu ESCRT do potenciálního místa uvolňování viru.[11] Podrobnosti o strojním zařízení ESCRT-I jsou popsány níže.
Komplex ESCRT-I je a heterotetramer (1: 1: 1: 1) z Vps23, Vps28, Vps37 a Mvb12.[3] Sestavený heterotetramer vypadá jako tyčová stopka složená z Vps23, Vps37 a Mvb12 s vějířovým víčkem složeným z jednoho šroubovice verzí Vps23, Vps28 a Vps37.[3][5] Vps23 obsahuje jednu variantní doménu ubiquitinu E2, která je zodpovědná za vazbu ubikvitinu, komplexu ESCRT-0 a na PTAP (strroline, threonin, ALanine, strroline) motiv viru Gag proteiny.[3][5] Těsně po této variantní doméně ubikvitinu E2, motiv bohatý na prolin (GPPX3Je přítomen Y), který směruje ESCRT-I do středního těla během abcise membrány.[5] Mvb12 může také vázat ubikvitin prostřednictvím jeho karboxy-konec. Vps28 je zodpovědný za interakci ESCRT-I a ESCRT-II přidružením k doméně GLUE (GRAM-Like Uvázání biquitinu v EAP45) Vps36 prostřednictvím svého karboxy-terminálu svazek čtyř šroubovic doména.[1]
ESCRT-II
Komplex ESCRT-II funguje primárně během biogeneze multivesikulárních tělísek a dodávání proteinů značených ubikvitinem do endosomu. Ubiquitinem značené proteiny jsou předávány z ESCRT-0 do ESCRT-I a poté do ESCRT-II. ESCRT-II se přidruží k ESCRT-III, který sevře náklad obsahující váček uzavřený.[5] Specifické aspekty ESCRT-II jsou následující:
ESCRT-II je heterotetramer (2: 1: 1) složený ze dvou Vps25 podjednotky, jedna Vps22 a jedna Vps36 podjednotka.[3] Molekuly Vps25 obsahují motivy PPXY, na které se váží okřídlená spirála (WH) motivy Vps22 a Vps36 vytvářející komplex ve tvaru Y s Vps22 a Vps36 jako základnou a Vps25 molekulami jako rameny.[3][5] Molekuly Vps25 také obsahují WH motivy, které jsou zodpovědné za interakci ESCRT-II s ESCRT-III. Vps36 obsahuje doménu GLUE, která váže fosfatidylinositol 3-fosfát a Vps28 ESCRT-I.[3][5] Dva zinkový prst domény jsou zacykleny do domény GLUE kvasinek Vps36. Jedna z těchto domén se zinkovým prstem váže karboxyterminální doménu Vps28 a druhá se asociuje s ubikvitinem.[5]
ESCRT-III
Komplex ESCRT-III je pravděpodobně nejdůležitější ze všech strojů ESCRT, protože hraje roli ve všech procesech zprostředkovaných ESCRT.[12] Během abscesu membrány a virového pučení vytváří ESCRT-III dlouhá vlákna, která se vinou kolem místa zúžení membrány těsně před membránovým štěpením.[10][13] K tomuto zprostředkování úrazu dochází prostřednictvím interakcí s centralspindlin komplex.[14] Tyto vláknité struktury jsou také přítomny během multivesikulární formace těla a fungují jako prstencový plot, který ucpává začínající váček, aby zabránil úniku nákladních proteinů do cytosolu buňky.[10] ESCRT-III existuje a funguje následovně:
Komplex ESCRT-III se liší od všech ostatních strojů ESCRT v tom, že existuje pouze přechodně a obsahuje základní i nepodstatné složky.[1][10] Základní podjednotky se musí shromažďovat ve správném pořadí (Vps20, Snf7, Vps24, pak Vps2), aby strojní zařízení fungovalo.[5] Mezi nepodstatné podjednotky patří Vps60, Did2 a Ist1.[10] Vps20 iniciuje sestavu ESCRT-III tím, že působí jako nukleator sestavy polymeru Snf7. Vps24 se poté přidruží k Snf7, aby komplex zakryl a získal Vps2.[1][3] Vps2 pak přináší Vps4 do komplexu.[15] Všechny „bezplatné“ cytosolické formy každé podjednotky jsou považovány za uzavřené. To znamená, že se karboxyterminální část každé podjednotky složí na sebe autoinhibičním způsobem stabilizujícím monomerní podjednotky.[1][3] Karboxy-konec většiny podjednotek ESCRT-III, základní i neesenciální, obsahuje MIM (MTO (mikrotubul interakční a transportní doména) iinterakce motif) motivy.[16] Tyto motivy jsou odpovědné za vazbu Vps4 a AAA-ATPáza spastin.[3]
Vps4-Vta1
Proteiny Vps4-Vta1 jsou vyžadovány pro stripování dalších složek ESCRT (obvykle ESCRT-III) z membrán, jakmile je konkrétní proces dokončen. Tam je nějaká debata o tom, zda Vps4 štěpí komplex ESCRT-III pryč nebo remodeluje komplex tak, aby se jedna složka vrhla v určitou dobu.[11] Předpokládá se, že Vta1 působí jako aktivátor Vps4, napomáhá jeho sestavení a zvyšuje jeho aktivitu AAA-ATPázy.[12][17] Způsob, jakým tyto proteiny fungují, je následující:
Podjednotky Vps4 mají dvě funkční domény, aminoterminální doménu MIT a centrální doménu AAA-ATPázy.[3] Doména MIT je zodpovědná za interakci Vps4 s doménou MIM Vps2.[1] Doména AAA-ATPázy hydrolyzuje ATP k energetické demontáži komplexu ESCRT-III.[10] Toto „odstranění“ ESCRT-III umožňuje recyklaci všech souvisejících podjednotek pro další použití.[10][11] Vta1 je dimerní protein obsahující jednu doménu VSL (tak pojmenovanou, protože se nachází v proteinech) PROTIps4, SBP1 a LIP5), který umožňuje vazbu na Vps4, a doménu MIT pro přidružení k podjednotce ESCRT-III Vps60. Ačkoli to není nezbytné, ukázalo se, že Vta1 pomáhá při sestavování prstenů Vps4, urychluje aktivitu ATPázy Vsp4 a podporuje demontáž ESCRT-III.[5]
Bro1
Hlavní funkcí Bro1 je nábor deubikvitinázy do komplexu ESCRT-III.[18] To má za následek odstranění ubikvitinových značek z proteinů zaměřených na degradaci v lysozomu těsně před vytvořením multivesikulárních těl. Rovněž se spekulovalo, že Bro1 pomáhá stabilizovat ESCRT-III, zatímco ubiquitinové značky jsou štěpeny z nákladových proteinů.[18]
Bro1 obsahuje Bro1 amino-terminální doménu, která se váže na Snf7 ESCRT-III.[19] Tato vazba přináší Bro1 na místo abcise membrány. Bro1 také váže katalytické doména Doa4, ubikvitin hydroláza (deubiquitináza), která ji přivádí na místo abscise. Doa4 odstraňuje ubikvitin z proteinů nákladu, které jsou cíleny na lysozom.[19]
Multivesikulární biogeneze těla a pendlování
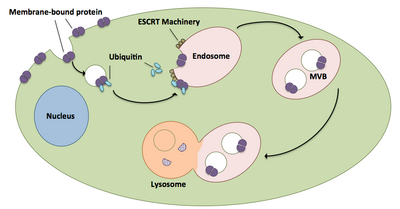
Multivesikulární tělesa hrají velkou roli v transportu ubikvitinovaných proteinů a receptorů do lysozomu.[20] Komplexy ESCRT transportují ubikvitinovaný náklad do buněčných vezikul, které se objevují přímo v endosomálním kompartmentu buňky a vytvářejí multivesikulární těla.[20] Tato multivesikulární tělesa se nakonec spojí s lysozomem a způsobí degradaci nákladu.[15] Podrobnější popis procesu, včetně souvisejících strojů, existuje takto:
- Jednotlivé komponenty ESCRT-0 Vps27 a Hse1 se vážou na ubikvitinovaný náklad.[1][20]
- Vps27 se váže na fosfatidylinositol-3-fosfát, endozomální lipid, který pak rekrutuje celý komplex do endosomu.[1][20]
- Vps27 váže podjednotku Vps23 ESCRT-I, čímž přivádí ESCRT-I do endosomu. ESCRT-I může také vázat ubikvitinované proteiny.[1][20]
- Vps36 se sdružuje s podjednotkou ESCRT-I Vps28, což vede k náboru komplexu ESCRT-II.[1]
- Podjednotka Vps25 ESCRT-II se váže na a aktivuje Vps20 komplexu ESCRT-III.[1][15][20]
- Vps20 nukleauje tvorbu řetězců Snf7, které jsou poté omezeny Vps24.[15]
- Vps24 rekrutuje Vps2, což přináší Vps4 do komplexu.[15]
- Vps4 tvoří póry ze dvou hexamerické kroužky, na které se váže Vta1.[1] Tento komplex Vps4-Vta1 spouští demontáž ESCRT-III a znamená konec multivezikulární formace těla.[2]
Absorpce membrány

Absorpce membrány v době cytokineze je proces, při kterém je membrána spojující dvě dceřiné buňky štěpena během buněčné dělení. Protože je konzervován v řadě Archaea „Absorpce membrány je považována za nejranější roli pro ESCRT stroje.[5] Proces začíná, když centrosomální protein CEP55 je přijímán do středního těla dělících se buněk ve spojení s MKLP1, a mitotický kinesin -jako protein, který se asociuje s mikrotubuly.[5][22] CEP55 poté rekrutuje podjednotku Vps23 z ESCRT-I a doplňkového proteinu ALIX, které se formují do kroužků na obou stranách středního těla.[5][10][11] ESCRT-I a ALIX získávají ESCRT-III prostřednictvím své podjednotky Snf7.[5] ESCRT-III podjednotky Vps20, Snf7, Vps24, Vps2 a Did2 se formují do spirálovitého vlákna sousedícího s prstenci tvořenými Vps23.[1][11][18] Tvorba této spirálovité struktury deformuje membránu a AAA-ATPázový spastin je přiveden pomocí Did2 a Ist1, aby štěpily mikrotubuly vytvořené ve středu těla.[11][18] Vps4 tedy katalyzuje demontáž komplexu ESCRT-III vedoucí ke dvěma nově odděleným dceřiným buňkám.[18] Proces membránové abcise byl popsán pomocí metazoanových proteinů, protože tento proces byl studován ve větší míře u metazoanů.
Virové pučení

Uvolňování virových částic, také známé jako virové pučení, je proces, kterým zdarma viriony jsou uvolňovány z buněk prostřednictvím únosu strojů ESCRT hostitelské buňky.[1][13] Retroviry, jako HIV-1 a lidský T-lymfotropní virus, stejně jako řada obalené viry, včetně Virus ebola, vyžadovat, aby ESCRT stroje opustily hostitelskou buňku.[1] Proces je zahájen virovými proteiny Gag, hlavními strukturálními proteiny retrovirových obalů, s nimiž interagují TSG101 komplexu ESCRT-I a doplňkového proteinu ALIX.[11][12] Podjednotky ESCRT-III (nezbytné jsou pouze CHMP4 a CHMP2 [9]) jsou rekrutováni do místa virového pučení, aby zúžili a oddělili krk pupíku podobným způsobem, jaký je popsán pro abscesy membrány během cytokineze.[1][5][11] Vps4 poté recykluje složky ESCRT-III do cytosolu a virus se uvolní z buňky.[5] Zde popsaný mechanismus využívá metazoanové proteiny, protože virové pučení bylo u metazoanů studováno extenzivněji.
Reference
- ^ A b C d E F G h i j k l m n Ó str q r s Schmidt O, Teis D (únor 2012). „Strojní zařízení ESCRT“. Curr. Biol. 22 (4): R116–20. doi:10.1016 / j.cub.2012.01.028. PMC 3314914. PMID 22361144.
- ^ A b Babst M (srpen 2011). „Tvorba vezikul MVB: závislá na ESCRT, nezávislá na ESCRT a vše mezi tím“. Curr. Opin. Cell Biol. 23 (4): 452–7. doi:10.1016 / j.ceb.2011.04.008. PMC 3148405. PMID 21570275.
- ^ A b C d E F G h i j k l Hurley JH, Hanson PI (srpen 2010). „Membránové pučení a štěpení strojním zařízením ESCRT: je to všechno na krku“. Nat. Rev. Mol. Cell Biol. 11 (8): 556–66. doi:10.1038 / nrm2937. PMC 2922035. PMID 20588296.
- ^ A b Piper RC, Katzmann DJ (2007). "Biogeneze a funkce multivesikulárních těles". Annu. Rev. Cell Dev. Biol. 23: 519–47. doi:10.1146 / annurev.cellbio.23.090506.123319. PMC 2911632. PMID 17506697.
- ^ A b C d E F G h i j k l m n Ó str q r s t Hurley JH (prosinec 2010). „Komplexy ESCRT“. Krit. Biochem. Mol. Biol. 45 (6): 463–87. doi:10.3109/10409238.2010.502516. PMC 2988974. PMID 20653365.
- ^ Wollert T, Hurley JH (duben 2010). "Molekulární mechanismus multivesikulární biogeneze těla pomocí komplexů ESCRT". Příroda. 464 (7290): 864–9. Bibcode:2010Natur.464..864W. doi:10.1038 / nature08849. PMC 2851844. PMID 20305637.
- ^ Ren X, Hurley JH (březen 2010). „VHS domény ESCRT-0 spolupracují při vazbě vysoké avidity na polyubiquitinovaný náklad“. EMBO J.. 29 (6): 1045–54. doi:10.1038 / emboj.2010.6. PMC 2845278. PMID 20150893.
- ^ Banerjee S, Basu S, Sarkar S (2010). „Srovnávací genomika odhaluje selektivní distribuci a doménovou organizaci proteinů domény FYVE a PX napříč eukaryotickými liniemi“. BMC Genomics. 11: 83. doi:10.1186/1471-2164-11-83. PMC 2837644. PMID 20122178.
- ^ A b Morita E, Sandrin V, McCullough J, Katsuyama A, Baci Hamilton I, Sundquist WI (březen 2011). „Požadavky na protein ESCRT-III pro začínající HIV-1“. Buněčný hostitelský mikrob. 9 (3): 235–42. doi:10.1016 / j.chom.2011.02.004. PMC 3070458. PMID 21396898.
- ^ A b C d E F G h Adell MA, Teis D (říjen 2011). „Montáž a demontáž komplexu štěpení membrány ESCRT-III“. FEBS Lett. 585 (20): 3191–6. doi:10.1016 / j.febslet.2011.09.001. PMC 3192940. PMID 21924267.
- ^ A b C d E F G h Mueller M, Adell MA, Teis D (srpen 2012). „Membránová abscise: první pohled na dynamické ESCRT“. Curr. Biol. 22 (15): R603–5. doi:10.1016 / j.cub.2012.06.063. PMC 3414845. PMID 22877781.
- ^ A b C McDonald B, Martin-Serrano J (červenec 2009). „Nejsou připojeny žádné řetězce: strojní zařízení ESCRT ve virovém pučení a cytokinéze“. J. Cell Sci. 122 (Pt 13): 2167–77. doi:10.1242 / jcs.028308. PMC 2723143. PMID 19535732.
- ^ A b Jouvenet N, Zhadina M, Bieniasz PD, Simon SM (duben 2011). „Dynamika náboru proteinů ESCRT během retrovirové montáže“. Nat. Cell Biol. 13 (4): 394–401. doi:10.1038 / ncb2207. PMC 3245320. PMID 21394083.
- ^ Glotzer, Michael. „Cytokinesis: Centralspindlin Moonlight as a Membrane Anchor“, Aktuální biologie, 18. února 2013
- ^ A b C d E Teis D, Saksena S, Judson BL, Emr SD (březen 2010). „ESCRT-II koordinuje sestavu vláken ESCRT-III pro třídění nákladu a tvorbu vezikul těla vícečetných těl“. EMBO J.. 29 (5): 871–83. doi:10.1038 / emboj.2009.408. PMC 2837172. PMID 20134403.
- ^ Scott A, Gaspar J, Stuchell-Brereton MD, Alam SL, Skalicky JJ, Sundquist WI (září 2005). "Struktura a ESCRT-III proteinové interakce MIT domény lidského VPS4A". Proc. Natl. Acad. Sci. USA. 102 (39): 13813–8. Bibcode:2005PNAS..10213813S. doi:10.1073 / pnas.0502165102. PMC 1236530. PMID 16174732.
- ^ Azmi I, Davies B, Dimaano C, Payne J, Eckert D, Babst M, Katzmann DJ (únor 2006). „Recyklace ESCRT pomocí AAA-ATPázy Vps4 je regulována konzervovanou oblastí VSL ve Vta1“. J. Cell Biol. 172 (5): 705–17. doi:10.1083 / jcb.200508166. PMC 2063703. PMID 16505166.
- ^ A b C d E Babst M, Davies BA, Katzmann DJ (říjen 2011). "Regulace Vps4 během třídění MVB a cytokineze". Provoz. 12 (10): 1298–305. doi:10.1111 / j.1600-0854.2011.01230.x. PMC 3171586. PMID 21658171.
- ^ A b Wemmer M, Azmi I, West M, Davies B, Katzmann D, Odorizzi G (leden 2011). „Vazba Bro1 na Snf7 reguluje aktivitu štěpení membrány ESCRT-III v kvasinkách“. J. Cell Biol. 192 (2): 295–306. doi:10.1083 / jcb.201007018. PMC 3172170. PMID 21263029.
- ^ A b C d E F Hurley JH, Emr SD (2006). „Komplexy ESCRT: struktura a mechanismus sítě pro obchodování s membránami“. Annu Rev Biophys Biomol Struct. 35: 277–98. doi:10.1146 / annurev.biophys.35.040405.102126. PMC 1648078. PMID 16689637.
- ^ Carmena M (červenec 2012). „Kontrola kontrolního bodu úniku: zaseknutá uprostřed s Aurorou B“. Otevřená Biol. 2 (7): 120095. doi:10.1098 / rsob.120095. PMC 3411112. PMID 22870391.
- ^ Zhu C, Bossy-Wetzel E, Jiang W (červenec 2005). „Nábor MKLP1 do vřetenové midzony / midbody pomocí INCENP je nezbytný pro tvorbu midbody a dokončení cytokineze v lidských buňkách“. Biochem. J. 389 (Pt 2): 373–81. doi:10.1042 / BJ20050097. PMC 1175114. PMID 15796717.