Vize u ryb - Vision in fishes

Vidění je důležité senzorický systém pro většinu druhů Ryba. Ryba oči jsou podobné očím pozemní obratlovců jako ptactvo a savci, ale mají více sférický objektiv. Ptáci a savci (včetně lidí) se obvykle přizpůsobují soustředit se změnou tvaru jejich čočky, ale ryby obvykle upravují zaostření pohybem čočky blíže nebo dále od sítnice. Rybí sítnice mají obvykle obojí tyčové buňky a kuželové buňky (pro skotopický a fotopické vidění ) a většina druhů má barevné vidění. Některé ryby vidí ultrafialový a některé jsou citlivé polarizované světlo.
Mezi ryby bez čelistí, mihule [1] má dobře vyvinuté oči, zatímco hagfish má pouze primitivní oční skvrny.[2] Předchůdci moderních hagfishů, považovaných za protovrchovce[3] byly evidentně tlačeny do velmi hlubokých, temných vod, kde byly méně náchylné k vidícím predátorům a kde je výhodné mít konvexní oční bod, který shromažďuje více světla než plochý nebo konkávní. Vize ryb ukazuje evoluční přizpůsobování například do jejich vizuálního prostředí hlubinné ryby mají oči vhodné do tmavého prostředí.
Voda jako vizuální prostředí


Ryby a další vodní živočichové žijí v jiném světelném prostředí než žijí suchozemské druhy. Voda absorbuje světlo takže s rostoucí hloubkou množství dostupného světla rychle klesá. Optické vlastnosti vody také vedou k různým vlnové délky světla absorbovaného v různých stupních. Například viditelné světlo dlouhých vlnových délek (např. Červená, oranžová) je absorbováno více ve vodě než světlo kratších vlnových délek (zelená, modrá). Ultrafialové světlo (dokonce kratší vlnová délka než fialové) může proniknout hlouběji než vizuální spektra [5] Kromě těchto univerzálních vlastností vody mohou různé vodní útvary absorbovat světlo různých vlnových délek v důsledku měnící se soli a / nebo chemické přítomnosti ve vodě.
Voda velmi účinně pohlcuje přicházející světlo, takže množství světla pronikajícího do oceánu s hloubkou rychle klesá (je oslabeno). V čisté mořské vodě zůstává v hloubce jednoho metru pouze 45% sluneční energie, která dopadá na povrch oceánu. V hloubce 10 metrů je stále přítomno pouze 16% světla a pouze 1% původního světla je ponecháno na 100 metrů. Za 1 000 metrů nepronikne žádné světlo.[6]
Kromě celkového útlumu oceány absorbují různé vlnové délky světla různými rychlostmi. Vlnové délky na extrémních koncích viditelného spektra jsou oslabeny rychleji než vlnové délky uprostřed. Nejprve jsou absorbovány delší vlnové délky. V čistých vodách oceánu je červená absorbována v horních 10 metrech, oranžová asi 40 metrů a žlutá zmizí před 100 metry. Kratší vlnové délky pronikají dále a modré a zelené světlo zasahuje do nejhlubších hloubek.[6]
Proto se věci pod vodou zdají modré. To, jak jsou barvy vnímány okem, závisí na vlnových délkách světla, které oko přijímá. Objekt vypadá na oko červeně, protože odráží červené světlo a pohlcuje jiné barvy. Jedinou barvou, která dosáhne do oka, je tedy červená. Modrá je jediná barva světla dostupná v hloubce pod vodou, takže je to jediná barva, která se může odrážet zpět do očí, a vše má pod vodou modrý nádech. Červený objekt v hloubce se neobjeví červeně, protože není k dispozici žádné červené světlo, které by odráželo od objektu. Objekty ve vodě se objeví jako jejich skutečné barvy poblíž povrchu, kde jsou stále k dispozici všechny vlnové délky světla, nebo pokud jsou ostatní vlnové délky světla poskytovány uměle, například osvětlením objektu potápěčským světlem.[6]
Struktura a funkce


Rybí oči jsou velmi podobné očím ostatních obratlovců - zejména pak tetrapody (obojživelníci, plazi, ptáci a savci - všichni se vyvinuli z předka ryb). Světlo vstupuje do oka na rohovka, procházející přes žák dosáhnout objektiv. Zdá se, že většina druhů ryb má pevnou velikost zornice, ale elasmobranches (jako žraloci a paprsky) mají svalstvo duhovka což umožňuje upravit průměr zornice. Tvar zornice se liší a může být např. kruhové nebo štěrbinové.[5]
Čočky jsou obvykle sférické, ale u některých druhů mohou být mírně eliptické. Ve srovnání s suchozemskými obratlovci jsou čočky ryb obecně hustší a sférické. Ve vodním prostředí není zásadní rozdíl v index lomu rohovky a okolní vody (ve srovnání se vzduchem na zemi), takže čočku musí udělat většinu lomu.[7] Kvůli index lomu gradient uvnitř čočky - přesně tak, jak by se dalo očekávat od optické teorie ",[8] sférické čočky ryb jsou schopné vytvářet ostré obrazy bez sférická aberace.[7]
Jakmile světlo prochází čočkou, je přenášeno průhledným kapalným médiem, dokud nedosáhne sítnice, která obsahuje fotoreceptory. Stejně jako ostatní obratlovci jsou fotoreceptory ve vnitřní vrstvě, takže světlo musí procházet vrstvami jiných neurony než k nim dorazí. Sítnice obsahuje prutové buňky a kuželové buňky.[5]Mezi očima ryb a jinými obratlovci jsou podobnosti. Světlo obvykle vstupuje rybím okem do rohovky a prochází zornicí, aby dosáhlo čočky. Většina druhů ryb má pevnou velikost zornice, zatímco několik druhů má svalovou duhovku, která umožňuje upravit průměr zornice.
Rybí oči mají sférickější čočku než ostatní suchozemští obratlovci. Úprava zaostření u savců a ptáků se obvykle provádí změnou tvaru oční čočky, zatímco u ryb se to provádí pohybem čočky dále od nebo blíže k sítnici. Sítnice ryby má obecně jak prutové buňky, tak kuželové buňky, které jsou zodpovědné za skotopické a fotopické vidění. Většina druhů ryb má obvykle barevné vidění. Existují druhy, které jsou schopné vidět ultrafialové záření, zatímco jiné jsou citlivé na polarizované světlo.[9]
Rybí sítnice má prutové buňky, které poskytují vysokou vizuální citlivost za špatných světelných podmínek, a kuželové buňky, které poskytují vyšší časové a prostorové rozlišení, než jaké jsou tyčinkové buňky schopné. Umožňují možnost barevného vidění porovnáním absorbance mezi různými typy kuželů.[10] Podle Marshalla et al., většina zvířat v mořském prostředí nemá žádné nebo relativně jednoduché barevné vidění. V oceánu však existuje větší rozmanitost barevného vidění, než je tomu na souši. To je způsobeno zejména extrémy fotického prostředí a barevného chování.[11]
Sítnice
V sítnici tyčové buňky poskytují vysokou vizuální citlivost (za cenu ostrost ), který se používá za špatných světelných podmínek. Kónické buňky poskytují vyšší prostorové a časové rozlišení, než mohou tyčinky, a umožňují možnost barevného vidění porovnáním absorbance napříč různými typy kuželů, které jsou citlivější na různé vlnové délky. Poměr prutů a šišek závisí na ekologii příslušných druhů ryb, např.ti, kteří jsou během dne aktivní hlavně v čisté vodě, budou mít více kuželů než ti, kteří žijí ve špatném osvětlení. Barevné vidění je užitečnější v prostředích se širší dostupnou vlnovou délkou, např., blízko povrchu v čistých vodách spíše než v hlubších vodách, kde přetrvává jen úzký pás vlnových délek.[5]
Distribuce fotoreceptorů po sítnici není jednotná. Některé oblasti mají například vyšší hustotu kuželových buněk (viz fovea ). Ryby mohou mít dvě nebo tři oblasti specializované na vysokou ostrost (např. K zachycení kořisti) nebo citlivost (např. Ze slabého světla přicházejícího zespodu). Distribuce fotoreceptorů se může v průběhu vývoje jedince také měnit. To platí zejména v případě, kdy se druh během svého životního cyklu obvykle pohybuje mezi různými světelnými prostředími (např. Mělká do hlubokých vod nebo sladká voda do oceánu).[5] nebo když změny potravinového spektra doprovázejí růst ryb, jak je vidět na antarktické ledovce Champsocephalus gunnari.[12]
Některé druhy mají a tapetum, reflexní vrstva, která odráží světlo, které prochází sítnicí zpět skrz ni. To zvyšuje citlivost za špatných světelných podmínek, jako jsou noční a hlubinné druhy, protože dává fotonům druhou šanci být zachycen fotoreceptory.[7] To však přichází za cenu sníženého rozlišení. Některé druhy jsou schopny účinně vypínat své tapetum za jasných podmínek a podle potřeby jej pokrývá tmavá pigmentová vrstva.[5]
Sítnice používá ve srovnání s většinou ostatních tkání hodně kyslíku a je zásobována dostatečným množstvím okysličené krve, aby byl zajištěn optimální výkon.[5]
Ubytování
Ubytování je proces, kterým se oko obratlovců přizpůsobuje soustředit se na objekt, když se pohybuje blíže nebo dál. Zatímco ptáci a savci dosahují akomodace deformací oční čočky, ryb a obojživelníci normálně upravit soustředit se pohybem čočky blíže nebo dále od sítnice.[5] Používají speciální sval, který mění vzdálenost čočky od sítnice. v kostnaté ryby sval se nazývá navíječ lentis, a je uvolněný pro vidění na blízko, zatímco pro paryby sval se nazývá úhloměr lentis, a je uvolněný pro vidění do dálky. Kostnaté ryby se tak přizpůsobí vidění na dálku pohybem čočky blíže k sítnici, zatímco chrupavčité ryby se přizpůsobí vidění na blízko pohybem čočky dále od sítnice.[13][14][15]Akomodace označuje proces, kterým oko obratlovců upravuje své zaměření na jakýkoli konkrétní objekt, když se pohybuje dále od oka nebo blíže k oku. Savci a ptáci obvykle dosáhnou akomodace prostřednictvím deformace oční čočky. Na druhou stranu ryby obvykle dosáhnou akomodace pohybem čočky blíže nebo dále od sítnice.
Stabilizace obrazu

Je potřeba nějaký mechanismus, který stabilizuje obrázky během rychlých pohybů hlavy. Toho je dosaženo vestibulo-oční reflex, což je reflex pohyb očí který stabilizuje obrázky na sítnice produkcí pohybů očí ve směru opačném k pohybům hlavy, čímž se zachová obraz ve středu zorného pole. Například když se hlava pohybuje doprava, oči se pohybují doleva a naopak. Lidský vestibulo-oční reflex je a reflex pohyb očí že stabilizuje obrázky na sítnice během pohybu hlavy vytvořením pohybu očí ve směru opačném k pohybu hlavy, čímž se zachová obraz ve středu zorného pole. Podobným způsobem mají ryby vestibulo-oční reflex, který stabilizuje vizuální obrazy na sítnici, když pohybuje ocasem.[16] U mnoha zvířat, včetně lidí, se vnitřní ucho funguje jako biologický analog an akcelerometr v systémech stabilizace obrazu fotoaparátu stabilizovat obraz pohybem oči. Je-li detekována rotace hlavy, je odeslán inhibiční signál do extraokulární svaly na jedné straně a excitační signál do svalů na druhé straně. Výsledkem je kompenzační pohyb očí. Typické pohyby očí člověka zaostávají za pohyby hlavy o méně než 10 ms.[17]
Diagram vpravo ukazuje horizontálu vestibulo-oční reflex obvody v kostnatý a paryby.
- "Zlatá rybka" ukazuje hlavní tří neuronový vestibulo-oční reflex spojující vodorovný půlkruhový kanál s kontralaterální únosci (ABD) a ipsilaterální PAN motoneurony.[18]
- „Platýz“ ukazuje, že po 90 ° posunutí vestibulu vzhledem k vizuální ose (metamorfóza) jsou kompenzační pohyby očí produkovány přesměrováním signálů vodorovného kanálu na vertikální a šikmé motoneurony.[19][20]
- V „žraločím“ vodorovném kanálu / druhém pořadí neurony projekt do kontralaterální ABD a MR motoneurony včetně ipsilaterální AI neurony. 1 °, první objednávka vestibulární neuron; ATD, Vzestupná trakt Deiterova.[20]
Ultrafialový
Rybí vidění je zprostředkováno čtyřmi vizuálními pigmenty, které absorbují různé vlnové délky světla. Každý pigment je vyroben z a chromofor a transmembránový protein, známý jako opsin. Mutace v opsinu umožnily vizuální rozmanitost, včetně variací absorpce vlnových délek.[21] Mutace opsinu na pigmentu SWS-1 umožňuje některým obratlovcům absorbovat UV světlo (≈360 nm), takže mohou vidět objekty odrážet UV světlo.[22] Tuto vizuální vlastnost si během evoluce vyvinula a udržovala široká škála druhů ryb, což naznačuje, že je to výhodné. UV vidění může souviset s hledáním potravy, komunikací a výběrem partnera.
Hlavní teorie týkající se evoluční selekce UV vidění u vybraných druhů ryb je dána její silnou rolí při výběru partnera. Experimenty s chováním ukazují, že africké cichlidy používají při výběru partnera vizuální podněty. Jejich hnízdiště jsou typicky v mělkých vodách s vysokou čistotou a UV zářením. Mužské africké cichlidy mají převážně modrou barvu, která je reflexní v UV světle. Ženy jsou schopny správně zvolit partnera svého druhu, pokud jsou přítomny tyto reflexní vizuální narážky. To naznačuje, že detekce UV světla je zásadní pro správný výběr partnera.[23] UV reflexní barevné vzory také zvyšují mužskou atraktivitu u guppies a třízubých sticklebacků. V experimentálních podmínkách strávily samičky guppies podstatně více času kontrolou mužů s UV-reflexním zbarvením než u těch, které měly UV odraz zablokovány.[24] Podobně samice tříprstých lipňáků upřednostňovaly muže pozorované v celém spektru před těmi pozorovanými v UV blokujících filtrech.[25] Tyto výsledky silně naznačují roli UV detekce v sexuálním výběru, a tedy v reprodukční zdatnosti. Prominentní role detekce UV světla při volbě kamarádů ryb umožnila, aby byla vlastnost zachována v průběhu času. Vidění UV může také souviset s hledáním potravy a dalším komunikačním chováním.
Mnoho druhů ryb může vidět ultrafialový konec spektra, za fialovou.[26]
Ultrafialové vidění se někdy používá pouze během části životního cyklu ryby. Například mladistvý pstruh potoční žijí v mělké vodě, kde používají ultrafialové vidění ke zvýšení své schopnosti detekce zooplankton. Jak stárnou, pohybují se do hlubších vod, kde je málo ultrafialového světla.[27]
The dva pruhy motýlice, Dascyllus reticulatus, má ultrafialové záření barvení které podle všeho používají jako výstražný signál jiným rybám svého druhu.[28] Dravý druhy to nemohou vidět, pokud jejich vidění není citlivé na ultrafialové záření. Pro tento názor existují další důkazy, že některé ryby používají ultrafialové záření jako „vysoce věrný tajný komunikační kanál skrytý před predátory“, zatímco jiné druhy používají ultrafialové záření k vytváření sociálních nebo sexuálních signálů.[5][29]
Polarizované světlo
Není snadné zjistit, zda je ryba citlivá polarizované světlo, i když se jeví jako pravděpodobné u řady taxonů. Bylo to jednoznačně prokázáno v ančovičky.[30]Schopnost detekovat polarizované světlo může poskytnout migrujícím druhům lepší kontrast a / nebo směrové informace. Polarizované světlo je nejhojnější za úsvitu a soumraku.[5] Polarizované světlo odražené od šupin ryby může ostatním rybám umožnit lepší detekci na rozptýleném pozadí,[31] a mohou poskytnout užitečné informace pro školní ryby o jejich blízkosti a orientaci ve vztahu k sousedním rybám.[32] Některé experimenty naznačují, že pomocí polarizace mohou některé ryby vyladit své vidění tak, aby jim bylo zdvojnásobeno jejich normální pozorovací vzdálenosti.[9]
Dvojité kužely
Většina ryb má dvojité kužely, pár kuželových buněk spojených k sobě navzájem. Každý člen dvojitého kužele může mít jinou špičkovou absorbanci a důkazy o chování podporují myšlenku, že každý typ jednotlivého kužele ve dvojitém kužele může poskytnout samostatnou informaci (tj. Signál od jednotlivých členů dvojitého kužele nemusí být nutně sčítán dohromady) .[33]
Přizpůsobení stanovišti

The čtyřnohá ryba živí se na povrchu vody očima, které jí umožňují vidět současně nad i pod hladinu.

1) Podvodní sítnice 2) Objektiv 3) Vzduchová pupila
4) Tkáňový pás 5) Iris 6) Podvodní zornice
7) Vzduchová sítnice 8) Optický nerv
Ryby, které žijí v povrchových vodách až do asi 200 metrů, epipelagické ryby žijí v zóně osvětlené sluncem, kde vizuální predátoři používají vizuální systémy, které jsou navrženy tak, jak by se dalo očekávat. Ale i tak mohou nastat neobvyklé úpravy. Čtyřoká ryba mají oči zvednuté nad temeno hlavy a rozdělené na dvě různé části, takže vidí současně pod i nad vodní hladinu. Ryby se čtyřmi očima mají ve skutečnosti jen dvě oči, ale jejich oči jsou speciálně přizpůsobeny jejich životnímu stylu obydlí. Oči jsou umístěny na temeni hlavy a ryba se vznáší na vodní hladině, přičemž pod vodou je pouze spodní polovina každého oka. Obě poloviny jsou rozděleny páskem tkáně a oko má dvě žáci, propojený částí duhovka. Horní polovina oka je přizpůsobena pro vidění ve vzduchu, dolní polovina pro vidění ve vodě.[34] Čočka oka se mění v tloušťce shora dolů, aby odpovídala rozdílu v indexy lomu vzduchu proti vodě. Tyto ryby tráví většinu času na hladině vody. Jejich strava se většinou skládá ze suchozemského hmyzu, který je k dispozici na povrchu.[35]
Mesopelagické ryby žít v hlubších vodách, v zóně soumraku až do hloubky 1 000 metrů, kde množství dostupného slunečního světla nestačí na podporu fotosyntéza. Tyto ryby jsou přizpůsobeny pro aktivní život za špatných světelných podmínek. Většina z nich jsou vizuální predátoři s velkýma očima. Některé ryby s hlubší vodou mají tubulární oči s velkými čočkami a pouze tyčové buňky které vypadají vzhůru. Ty poskytují binokulární vidění a velkou citlivost na malé světelné signály.[36] Tato adaptace poskytuje vylepšené terminální vidění na úkor laterálního vidění a umožňuje dravci vyzvednout se oliheň, sépie obecná a menší ryby, které jsou siluety proti temnotě nad nimi. Pro citlivější vidění při slabém osvětlení, některé ryby mají odrazka za sítnice. Svítilna ryby mít toto plus fotofóry, které používají v kombinaci k detekci oči u jiných ryb.[37][38][39]
Stále hlouběji vodní sloup, pod 1000 metrů, se nacházejí bathypelagické ryby. V této hloubce je oceán černý a ryby jsou sedavé, přizpůsobené k produkci minimální energie v prostředí s velmi malým množstvím potravy a bez slunečního záření. Bioluminiscence je jediné světlo dostupné v těchto hloubkách. Tento nedostatek světla znamená, že se organismy musí spoléhat na jiné smysly než na vidění. Jejich oči jsou malé a nemusí vůbec fungovat.[40][41]

Hlubinné ryby, jako je tento Antarktický zubáček, často mají velké, vzhůru vyhlížející oči, přizpůsobené k detekci kořisti, která je siluetována proti temnotě výše.[42]

The dalekohled má velké, dopředu směřující teleskopické oči s velkými čočkami.[43]

The mezopelagický šavlozubý je přepadení dravce s teleskopickýma očima směřujícím nahoru.


Na samém dně oceánu platýs Může být nalezeno. Platýsy jsou bentické ryby s negativním vztlakem, aby mohli odpočívat na mořském dně. Ačkoli platýsi žijí při dně, obvykle nejde o hlubinné ryby, ale vyskytují se hlavně v ústí řek a na kontinentálním šelfu. Když platýs larvy poklop mají protáhlý a symetrický tvar typického kostnatá ryba. Larvy nebývají na dně, ale plavou v moři jako plankton. Nakonec se začnou proměňovat v dospělou formu. Jedno z očí migruje přes temeno hlavy a na druhou stranu těla, přičemž na jedné straně nechává ryby slepé. Larva ztrácí své plynový měchýř a trny a klesají ke dnu a položí svou slepou stranu na podkladový povrch.[44] Richard Dawkins vysvětluje to jako příklad evoluční adaptace
... kostnaté ryby mají zpravidla výraznou tendenci být zploštěny ve svislém směru .... Bylo proto přirozené, že když se předkové [platýse] dostali na mořské dno, měli ležet na jednom boční.... Ale to vyvolalo problém, že jedno oko se vždy dívalo dolů do písku a bylo skutečně k ničemu. V evoluci byl tento problém vyřešen dolním okem „pohybujícím se“ kolem na horní stranu.[45]

Většina hlubinných ryb nevidí červené světlo. Hluboká voda semafor loosejaw produkuje červenou bioluminiscenci, takže může lovit s efektivně neviditelným paprskem světla.[46]

Když larvy a platýs roste, oko na jedné straně se otáčí na druhou stranu, takže ryby mohou odpočívat na mořském dně

The Platýs evropský je platýs se zvednutýma očima, takže když se zahrabává do písku pro maskování, stále vidí



Kořist má obvykle oči po stranách hlavy, takže má velké zorné pole, ze kterého se vyhne predátorům. Predátoři mají obvykle oči před hlavou, takže se mají lépe vnímání hloubky.[47][48] Bentičtí dravci, jako platýs, mají oči uspořádané tak, aby měli binokulární pohled na to, co je nad nimi, když leží na dně.
Barvení
Ryby si vyvinuly sofistikované způsoby použití barvení. Například, kořist ryby mít způsoby, jak pomocí zbarvení ztížit vizuální dravce, aby je viděli. v pelagické ryby, tyto úpravy se zabývají hlavně zmenšením siluety, formou maskovat. Jedním ze způsobů, jak toho dosáhnout, je zmenšit plochu jejich stínu boční kompresí těla. Další metodou, také formou maskování, je protisvětlo v případě epipelagické ryby a tím protisvětlo v případě mezopelagické ryby. Protichůdného odstínu je dosaženo zbarvením ryb tmavšími pigmenty nahoře a světlejšími pigmenty na dně tak, aby zbarvení odpovídalo pozadí. Při pohledu shora se tmavší hřbetní oblast zvířete mísí s temnotou vody pod vodou a při pohledu zespodu se světlejší ventrální oblast mísí se slunečním světlem z povrchu. Protisvětlo je dosaženo pomocí bioluminiscence produkcí světla z ventrálního fotofóry, zaměřené na přizpůsobení intenzity světla ze spodní strany ryby intenzitě světla z pozadí.[49]
Bentická ryba, kteří spočívají na mořském dně, fyzicky se skrývají zahrabáním do písku nebo ústupem do zákoutí nebo se maskují splynutím s pozadím nebo vypadáním jako skála nebo kousek mořských řas.[50]
I když tyto nástroje mohou být účinné jako mechanismy vyhýbání se predátorům, slouží také jako stejně účinné nástroje pro samotné predátory. Například hlubinná samet břicho lucerna žralok používá protisvětlo, aby se skryl před svou kořistí.[51]

Epipelagické ryby, jako je tato Tuňák obecný v Atlantiku, jsou typicky zahloubený se stříbrnými barvami

The foureye butterflyfish má falešné oči na svém zadním konci, matoucí predátory, o nichž je přední konec ryby

The John Dory má velké oční místo uprostřed těla, matoucí kořist



Zobrazují se také některé druhy ryb falešné oči. The foureye butterflyfish dostává své jméno podle velké tmavé skvrny na zadní části každé strany těla. Toto místo je obklopeno zářivě bílým prstencem, který připomíná oční skvrnu. Pravým okem protéká černá svislá čára na hlavě, takže je těžké ji vidět.[52] To může mít za následek a dravec myslet si, že ryba je větší, než je, a zaměňovat zadní konec s předním koncem. Prvním instinktem motýlí ryby, když je ohrožen, je útěk, čímž se falešná skvrna očí přiblíží dravci než hlavě. Většina dravců míří na oči a tento falešný skvrn na oči přiměje dravce, aby věřil, že ryba prchne ocasem jako první.
The John Dory je benthopelagický pobřežní ryby s vysokým bočně stlačeným tělem. Jeho tělo je tak tenké, že je zepředu jen stěží vidět. Má také velkou tmavou skvrnu na obou stranách, která se používá k blikání „zlého oka“, pokud se blíží nebezpečí. Velké oči v přední části hlavy jí poskytují bifokální vidění a vnímání hloubky, které potřebuje k ulovení kořisti. The John Dory's oční skvrna na boku těla také plete kořist, která je poté nasávána do úst.[53]
Barreleyes

obvykle směřují nahoru, ale lze je také otočit dopředu
---------------------------------------------------------------------
Vpravo: Spookfish hnědý je jediným známým obratlovcem
použít zrcadlové oko (stejně jako čočku):
(1) divertikl (2) hlavní oko
(a) sítnice (b) reflexní krystaly (c) čočka (d) sítnice
Barreleyes jsou rodina malých, neobvykle vypadajících mezopelagických ryb, pojmenovaných pro své trubkovité oči ve tvaru sudu, které jsou obecně směrovány vzhůru, aby detekovaly siluety dostupné kořisti.[54][55] Barreleye mají velké, teleskopický oči, které dominují a vyčnívají z lebka. Tyto oči obvykle hledí vzhůru, ale u některých druhů je lze také otočit dopředu. Jejich oči mají velkou čočku a sítnici s výjimečným počtem tyčové buňky a vysoká hustota rhodopsin („vizuální fialový“ pigment); nejsou k dispozici žádné kuželové buňky.[54]
Barreleye druhy, Mikropomů Macropinna, má na vrcholu hlavy průhlednou ochrannou kopuli, něco jako kopuli nad kokpitem letadla, skrz kterou lze vidět čočky jejích očí. Kopule je tvrdá a pružná a pravděpodobně chrání oči před nematocysty (bodavé buňky) sifonofory ze kterého se věří, že barreleye krade jídlo.[54][55][56]
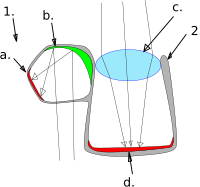
Další druh barreleye, brownsnout spookfish, je jediným obratlovcem, o kterém je známo, že používá zrcadlo, na rozdíl od čočky, aby zaostřil obraz v očích.[57][58] Je neobvyklé, že využívá obojí refrakční a reflexní optika vidět. Hlavní tubulární oko obsahuje laterální vejčitý otok zvaný a divertikl, do značné míry oddělené od oka a septum. Sítnice lemuje většinu vnitřku oka a existují dva rohovka otvory, jeden směřující nahoru a druhý dolů, které umožňují světlo do hlavního oka a do divertikula. Hlavní oko zaměstnává a objektiv zaostřit svůj obraz, jako u jiných ryb. Uvnitř divertikula se však světlo odráží a zaostřuje na sítnici zakřiveným složeným zrcadlem odvozeným od tapetum sítnice, složený z mnoha vrstev malých reflexních desek, případně vyrobených z guanin krystaly. Rozštěpená struktura oka spookfish brownsnout umožňuje rybám vidět současně nahoru i dolů. Kromě toho je zrcadlový systém při shromažďování světla lepší než čočka. Je pravděpodobné, že hlavní oko slouží k detekci objektů siluetou proti slunečnímu světlu, zatímco divertikulum slouží k detekci bioluminiscenční bliká ze stran a zespodu.[57]
Žraloci

Žralok oči jsou podobné očím ostatních obratlovců, včetně podobných čočky, rohovky a sítnice, i když jejich zrak je dobře přizpůsoben námořní prostředí pomocí tkáně zvané tapetum lucidum. Tato tkáň je za sítnice a odráží zpět světlo, čímž zvyšuje viditelnost v temných vodách. Účinnost tkáně se liší, přičemž někteří žraloci mají silnější účinek noční úpravy. Mnoho žraloků se může smrštit a rozšířit je žáci, jako lidé, něco ne teleost ryby může udělat. Žraloci mají víčka, ale nemrkají, protože okolní voda oči čistí. Kvůli ochraně očí některé druhy mají nictitující membrány. Tato membrána pokrývá oči při lovu a při útoku na žraloka. Některé druhy, včetně velký bílý žralok (Carcharodon carcharias), nemají tuto membránu, ale místo toho otočí oči dozadu, aby je ochránily při úderu na kořist. Důležitost zraku v chování lovu žraloků je diskutována. Někteří tomu věří elektro- a chemorecepce jsou významnější, zatímco jiné poukazují na nictující membránu jako na důkaz, že zrak je důležitý. Pravděpodobně by žralok neochránil oči, kdyby byly nedůležité. Použití zraku se pravděpodobně liší podle druhů a vodních podmínek. Žraločí zorné pole se může střídat monokulární a stereoskopický kdykoliv.[59] A mikrospektrofotometrie studie 17 druhů žraloků našel pouze 10 tyčové fotoreceptory a žádné kuželové buňky v jejich sítnice dávat jim dobré noční vidění a přitom je vyrábět barvoslepý. Zbývajících sedm druhů mělo kromě prutů jeden druh kuželový fotoreceptor citliví na zelenou a viděni pouze v odstínech šedé a zelené jsou považováni za účinně barvoslepé. Studie naznačuje, že pro detekci objektu může být důležitější kontrast objektu než pozadí, nikoli barva.[60][61][62]
Další příklady



Malé ryby často škola společně pro bezpečnost. To může mít vizuální výhody, jak vizuálně matoucí dravé ryby, a poskytnutím mnoha očí pro školu považovanou za tělo. „Efekt zmatení predátora“ je založen na myšlence, že pro predátory je obtížné vybrat ze skupin jednotlivou kořist, protože mnoho pohyblivých cílů vytváří senzorické přetížení vizuálního kanálu predátora.[63] „Hejnové ryby jsou stejně velké a stříbřité, takže pro vizuálně orientovaného predátora je těžké vybrat jednotlivce z masy kroucených a blikajících ryb a poté mít dostatek času na uchopení kořisti, než zmizí v hejnu.“[64] „Efekt mnoha očí“ je založen na myšlence, že s rostoucí velikostí skupiny může být úkol skenování prostředí pro predátory rozložen na mnoho jedinců, hromadná spolupráce pravděpodobně poskytuje vyšší úroveň bdělosti.[65][66]
Ryby jsou obvykle chladnokrevné a jejich tělesná teplota je stejná jako okolní voda. Některé však oceánské dravé ryby, jako mečoun a nějaký žralok a tuňák druhy, mohou zahřívat části těla, když loví kořist v hluboké a studené vodě. Vysoce vizuální mečoun používá topný systém zahrnující jeho svaly, který zvyšuje teplotu v očích a mozku až o 15 ° C. Zahřátí sítnice zlepšuje rychlost, jakou oči reagují na změny rychlého pohybu způsobeného kořistí, a to až desetkrát.[67][68][69]
Některé ryby mají oči.[70] Eyeshine je výsledkem vrstvy shromažďující světlo v očích zvané tapetum lucidum, který odráží bílé světlo. Nevyskytuje se u lidí, ale lze jej pozorovat u jiných druhů, například u jelenů ve světlometu. Eyeshine umožňuje rybám dobře vidět za špatných světelných podmínek i ve zakalených (špinavých nebo drsných, lámajících se) vodách, což jim dává výhodu nad jejich kořistí. Toto vylepšené vidění umožňuje rybám osídlit hlubší oblasti oceánu nebo jezera. Zejména sladkovodní Walleye jsou tak pojmenovaní, protože jejich oči svítí.[71]
Mnoho druhů Loricariidae, rodina sumec, mít upravený duhovka volal omega iris. Horní část duhovky sestupuje a vytváří smyčku, která se může rozpínat a smršťovat a která se nazývá iris operculum; když je úroveň světla vysoká, zornice se zmenšuje v průměru a smyčka se rozšiřuje tak, aby zakryla střed zornice, což vede k půlměsíc tvarovaná část přenášející světlo.[72] Název této funkce je odvozen od podobnosti s obrácenou Řecký dopis omega (Ω). Původy této struktury nejsou známy, ale předpokládá se, že prolomení obrysu vysoce viditelných očních pomůcek maskovat u často velmi strakatých zvířat.[72]
Distanční senzorické systémy

Vizuální systémy jsou systémy senzoriky vzdálenosti, které poskytují rybám údaje o poloze nebo objektech na dálku, aniž by se jich ryby musely přímo dotýkat. Takové systémy snímání vzdálenosti jsou důležité, protože umožňují komunikaci s jinými rybami a poskytují informace o umístění potravy a predátorů a vyhýbání se překážkám nebo udržování polohy v rybí školy. Například některé školní druhy mají po stranách „školní značky“, například vizuálně výrazné pruhy, které poskytují referenční značky a pomáhají sousedním rybám posoudit jejich relativní polohy.[74] Vizuální systém však není jediný, který může tyto funkce provádět. Některé školní ryby mají také boční čára po délce jejich těl. Tato boční linie umožňuje rybám snímat změny tlaku vody a turbulence sousedící s jejím tělem. S využitím těchto informací může školní ryba upravit svou vzdálenost od sousedních ryb, pokud se přiblíží nebo zbloudí příliš daleko.[74]
Vizuální systém u ryb je rozšířen o další snímací systémy se srovnatelnými nebo doplňkovými funkcemi. Některé ryby jsou slepí a musí se plně spoléhat na alternativní systémy snímání.[75] Mezi další smysly, které mohou také poskytovat údaje o poloze nebo vzdálených objektech, patří sluch a echolokace, elektrorecepce, magnetoception a chemorecepce (čich a chuť ). Například, sumec mají chemoreceptory napříč celým tělem, což znamená, že „ochutnají“ vše, čeho se dotknou, a „ucítí“ jakékoli chemikálie ve vodě. „Na sumci, degustace hraje hlavní roli v orientaci a umístění jídla “.[76]
Paryby (žraloci, rejnoky a chiméry) používají magnetocepci. Mají speciální elektroreceptory volal ampule Lorenzini které detekují mírnou změnu elektrického potenciálu. Tyto receptory umístěné podél tlamy a nosu ryby fungují podle principu, že časově proměnné magnetické pole pohybující se vodičem indukuje elektrický potenciál přes konce vodiče. Ampule mohou také umožnit rybám detekovat změny teploty vody.[77][78] Stejně jako u ptáků může magnetoception poskytnout informace, které pomohou migračním trasám ryb.[79]
Viz také
Poznámky
- ^ Meyer-Rochow, VB; Stewart, Duncan (1996). "Recenze ultrastruktury očních a postlarálních očí u mihule (Cyclostomata) se zvláštním důrazem na Geotria australis (šedá)". Mikroskopický výzkum a technika. 35 (6): 431–444. doi:10.1002 / (sici) 1097-0029 (19961215) 35: 6 <431 :: aid-jemt3> 3.0.co; 2-l. PMID 9016447.
- ^ N. A. Campbell a J. B. Reece (2005). Biologie, Sedmé vydání. Benjamin Cummings, San Francisco, Kalifornie.
- ^ Trevor D. Lamb, Shaun P. Collin a Edward N. Pugh, Jr. (2007). „Vývoj oka obratlovců: opsiny, fotoreceptory, sítnice a oční kalíšek“. Recenze přírody Neurovědy. 8 (12): 960–976. doi:10.1038 / nrn2283. PMC 3143066. PMID 18026166.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Ocean Explorer NOAA. Aktualizováno: 26. srpna 2010.
- ^ A b C d E F G h i j k Helfman a kol., 2009, s. 84-87.
- ^ A b C Webb, Paul (2019) Úvod do oceánografie, kapitola 6.5 Light, Komunita Rebus, Univerzita Rogera Williamse, otevřená učebnice.
 Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0.
Materiál byl zkopírován z tohoto zdroje, který je k dispozici pod a Mezinárodní licence Creative Commons Attribution 4.0. - ^ A b C d Land, M. F .; Nilsson, D. (2012). Zvířecí oči. Oxford University Press. ISBN 9780199581146.
- ^ Wehner, R (2005). "Senzorická fyziologie: oči bez mozku" (PDF). Příroda. 435 (7039): 157–159. Bibcode:2005 Natur.435..157W. doi:10.1038 / 435157a. PMID 15889076. S2CID 4408533.
- ^ A b Novales Flamarique, Iñigo (2019). "Swimming behaviour tunes fish polarization vision to double prey sighting distance". Vědecké zprávy. 9 (1): 944. Bibcode:2019NatSR...9..944N. doi:10.1038/s41598-018-37632-1. PMC 6353921. PMID 30700806.
- ^ Ingram, Norianne T.; Sampath, Alapakkam P.; Fain, Gordon L. (2016). "Why are rods more sensitive than cones?". The Journal of Physiology. 594 (19): 5415–5426. doi:10.1113/JP272556. PMC 5043029. PMID 27218707.
- ^ Marshall, Justin; Carleton, Karen L.; Cronin, Thomas (2015). "Colour vision in marine organisms". Aktuální názor v neurobiologii. 34: 86–94. doi:10.1016/j.conb.2015.02.002. PMID 25725325. S2CID 20978931.
- ^ Miyazaki, T; Iwamu, T; Meyer-Rochow, VB (2011). "The position of the retinal area centralis changes with age in Champsocephalus gunnari (Channichthyidae), a predatory fish from coastal Antarctic waters". Polární biologie. 34 (8): 1117–1123. doi:10.1007/s00300-011-0969-2. S2CID 19066809.
- ^ Schwab, IR; Hart, N (2006). "More than black and white". British Journal of Ophthalmology. 90 (4): 406. doi:10.1136/bjo.2005.085571. PMC 1857009. PMID 16572506.
- ^ Schwab, Ivan R. (2012) Evolution's Witness: How Eyes Evolved Page 82. Oxford University Press. ISBN 9780195369748.
- ^ Khorramshahia, O; Schartaua, JM; Krögera, RHH (2008). "A complex system of ligaments and a muscle keep the crystalline lens in place in the eyes of bony fishes (teleosts)". Vision Vision. 48 (13): 1503–1508. doi:10.1016/j.visres.2008.03.017. PMID 18471852. S2CID 17757889.
- ^ Singh H.R. and Khanna S.S. (1994) Advances in fish biology, str. 235, Hindustan Pub. ISBN 978-81-7075-029-1.
- ^ Barnes, GR (February 1979). „Vestibulo-oční funkce během koordinovaných pohybů hlavy a očí pro získání vizuálních cílů“. The Journal of Physiology. 287: 127–47. doi:10.1113 / jphysiol.1979.sp012650. PMC 1281486. PMID 311828.
- ^ Graf, W; Spencer, R; Baker, H; Baker, R (May 1997). "Excitatory and inhibitory vestibular pathways to the extraocular motor nuclei in goldfish". Journal of Neurophysiology. 77 (5): 2765–79. doi:10.1152/jn.1997.77.5.2765. PMID 9163391. S2CID 13004673.

- ^ Graf, W; Baker, R (October 1985). "The vestibuloocular reflex of the adult flatfish. II. Vestibulooculomotor connectivity". Journal of Neurophysiology. 54 (4): 900–16. doi:10.1152/jn.1985.54.4.900. PMID 4067626.
- ^ A b Graf, W; Spencer, R; Baker, H; Baker, R (September 2001). "Vestibuloocular reflex of the adult flatfish. III. A species-specific reciprocal pattern of excitation and inhibition". Journal of Neurophysiology. 86 (3): 1376–88. doi:10.1152/jn.2001.86.3.1376. PMID 11535684.
- ^ Yokoyama, S.; Yokoyama, R. (1996). "Adaptive evolution of photoreceptors and visual pigments in vertebrates". Výroční přehled ekologie a systematiky. 27: 543–567. doi:10.1146/annurev.ecolsys.27.1.543.
- ^ Shi, Y .; Yokoyama, S. (2003). "Molecular analysis of the evolutionary significance of ultraviolet vision in vertebrates". Sborník Národní akademie věd. 100 (14): 8308–8313. Bibcode:2003PNAS..100.8308S. doi:10.1073/pnas.1532535100. PMC 166225. PMID 12824471.
- ^ Carleton, K.L., Hárosi, F.I., & Kocher, T.D. (2000), Visual pigments of African cichlid fishes: Evidence for ultraviolet vision from microspectrophotometry and DNA sequences, Vision Research, 40(8), 879-890.
- ^ Kodric-Brown, A., & Johnson, S.C. (2002). Ultraviolet reflectance patterns of male guppies enhance their attractiveness to females, Animal Behaviour, 63(2), 391-396.
- ^ Rick, I.P., Modarressie, R., & Bakker, T.C.M. (2006). UV wavelengths affect female mate choice in three-spined sticklebacks, Animal Behaviour, 71(2), 307-313.
- ^ Jacobs, GH (1992). "Ultraviolet Vision in Vertebrates". Dopoledne. Zool. 32 (4): 544–554. doi:10.1093/icb/32.4.544.
- ^ Shi, Y; Yokoyama, S (2003). "Molecular analysis of the evolutionary significance of ultraviolet vision in vertebrates". PNAS. 100 (14): 8308–8313. Bibcode:2003PNAS..100.8308S. doi:10.1073/pnas.1532535100. PMC 166225. PMID 12824471.
- ^ Losey, G. S. Jr (2003). "Crypsis and communication functions of UV-visible coloration in two coral reef damselfish, Dascyllus aruanus a D. reticulatus". Chování zvířat. 66 (2): 299–307. doi:10.1006/anbe.2003.2214. S2CID 140204848.
- ^ Siebeck, UE; Parker, AN; Sprenger, D; Mäthger, LM; Wallis, G (2010). "A Species of Reef Fish that Uses Ultraviolet Patterns for Covert Face Recognition" (PDF). Aktuální biologie. 20 (5): 407–410. doi:10.1016/j.cub.2009.12.047. PMID 20188557. S2CID 3743161.
- ^ Horváth G and Varjú D (2004)Polarized light in animal vision: polarization patterns in nature str. 294, Springer. ISBN 978-3-540-40457-6.
- ^ Denton, EJ; Nichol, JAC (1965). "Polarization of light reflected from the silvery exterior of the bleak Alburnus alburnus" (PDF). J. Mar. Biol. Doc. U. K. 150: 78–94.
- ^ Rowe, DM; Denton, EJ (1997). "The physical basis of reflective communication between fish, with special reference to the horse mackerel, Trachurus trachurus". Phil. Trans. R. Soc. Lond. B. 352 (1353): 531–549. Bibcode:1997RSPTB.352..531R. doi:10.1098/rstb.1997.0037. PMC 1691948.
- ^ Pignatelli, V .; Champ, C .; Marshall, J .; Vorobyev, M. (2010). „Dvojité kužely se používají k rozlišování barev u útesových ryb, Rhinecanthus aculeatus". Biologické dopisy. 6 (4): 537–539. doi:10.1098 / rsbl.2009.1010. PMC 2936199. PMID 20129950.
- ^ Nelson, Joseph, S. (2006). Ryby světa. John Wiley & Sons, Inc. ISBN 978-0-471-25031-9.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2007). "Anableps anableps" v FishBase. Verze z března 2007.
- ^ Moyle and Cech, 2004, p. 585
- ^ Morin, James G.; Harrington, Anne; Nealson, Kenneth; Krieger, Neil; Baldwin, Thomas O .; Hastings, J. W. (1975). "Light for All Reasons: Versatility in the Behavioral Repertoire of the Flashlight Fish". Věda. 190 (4209): 74–76. Bibcode:1975Sci...190...74M. doi:10.1126/science.190.4209.74. S2CID 83905458.
- ^ McCosker JE (1977) "Flashlight fishes" Archivováno 2012-05-02 na Wayback Machine Scientific American, 236: 106–115.
- ^ Paxton, John R. (1998). Paxton, J.R .; Eschmeyer, W.N. (eds.). Encyklopedie ryb. San Diego: Academic Press. str. 162. ISBN 978-0-12-547665-2.
- ^ Ryan P "Deep-sea creatures: The bathypelagic zone" Te Ara - encyklopedie Nového Zélandu. Aktualizováno 21. září 2007.
- ^ Moyle and Cech, 2004, p. 587
- ^ Froese, Rainer a Pauly, Daniel, eds. (2009). "Dissostichus mawsoni" v FishBase. Verze ze srpna 2009.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2010). "Gigantura chuni" v FishBase. Verze z října 2010.
- ^ Chapleau, Francois & Amaoka, Kunio (1998). Paxton, J. R. a Eschmeyer, W. N. (eds.). Encyklopedie ryb. San Diego: Academic Press. xxx. ISBN 978-0-12-547665-2.
- ^ Dawkins, Richarde (1991). Slepý hodinář. London: Penguin Books. str. 92. ISBN 978-0-14-014481-9.
- ^ Kenaley, C.P (2007). "Revision of the Stoplight Loosejaw Genus Malacosteus (Teleostei: Stomiidae: Malacosteinae), with Description of a New Species from the Temperate Southern Hemisphere and Indian Ocean". Copeia. 2007 (4): 886–900. doi:10.1643 / 0045-8511 (2007) 7 [886: ROTSLG] 2.0.CO; 2.
- ^ "Masožravci". Americké ministerstvo vnitra, Úřad pro správu půdy. 14. 12. 2009. Archivovány od originál dne 14.06.2011. Citováno 2011-03-28.
- ^ Boroditsky, Lera (24 June 1999). "Light & Eyes: Lecture Notes". Poznámky z přednášky. Stanford. Archivovány od originál dne 5. července 2010. Citováno 11. května 2010.
- ^ Protisvětlo BBC: Science and Nature. Vyvolány 28 September 2011.
- ^ Fishy friends and fishy foes Preparation manual, Long Beach Marine Institute.
- ^ Claes, Julien M., Dag L. Aksnes & Jérôme Mallefet (2010). "Phantom hunter of the fjords: camouflage by counterillumination in a shark (Etmopterus spinax)" (PDF). Journal of Experimental Marine Biology and Ecology. 388 (1–2): 28–32. doi:10.1016/j.jembe.2010.03.009. Archivovány od originál (PDF) dne 2011-09-27. Citováno 2011-09-25.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ FishBase Froese, Rainer a Pauly, Daniel, eds. (2009). "Chaetodon capistratus" v FishBase. July 2009 version.
- ^ Walrond, Carl (2006) Coastal fish - Fish of the open sea floor, Te Ara: Encyklopedie Nového Zélandu. Accessed 28 May 2019.
- ^ A b C Robison, BH; Reisenbichler, KR (2008). "Macropinna microstoma and the Paradox of Its Tubular Eyes". Copeia. 2008 (4): 780–784. doi:10.1643/CG-07-082. S2CID 85768623.
- ^ A b Researchers solve mystery of deep-sea fish with tubular eyes and transparent head Monterey Bay Aquarium Research Institute, 23. února 2009.
- ^ Froese, Rainer a Pauly, Daniel, eds. (2011). "Macropinna microstoma" v FishBase. Verze ze září 2011.
- ^ A b Wagner, H.J., Douglas, R.H., Frank, T.M., Roberts, N.W., and Partridge, J.C. (Jan 27, 2009). "A Novel Vertebrate Eye Using Both Refractive and Reflective Optics". Aktuální biologie. 19 (2): 108–114. doi:10.1016/j.cub.2008.11.061. PMID 19110427. S2CID 18680315.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Smith, L. (Jan. 8, 2009). "Fish with four eyes can see through the deep sea gloom". Times Online. Times Newspapers Ltd. Retrieved on March 14, 2009.
- ^ Martin, R. Aidan. "Vision and a Carpet of Light". Centrum ReefQuest pro výzkum žraloků. Citováno 2009-08-22.
- ^ "Sharks are colour-blind, new study finds". Archivovány od originál dne 24.01.2011. Citováno 2011-02-03.
- ^ Gill, Victoria (2011-01-18). "Sharks are probably colour-blind". BBC novinky. Citováno 2011-01-19.
- ^ Nathan Scott Hart, Susan Michelle Theiss, Blake Kristin Harahush and Shaun Patrick Collin (2011). "Microspectrophotometric evidence for cone monochromacy in sharks". Naturwissenschaften. 98 (3): 193–201. Bibcode:2011NW.....98..193H. doi:10.1007/s00114-010-0758-8. PMID 21212930. S2CID 30148811.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Milinski, H.; Heller, R. (1978). "Influence of a predator on the optimal foraging behavior of sticklebacks". Příroda. 275 (5681): 642–644. Bibcode:1978Natur.275..642M. doi:10.1038/275642a0. S2CID 4184043.
- ^ Moyle and Cech, 2004.
- ^ Roberts, G (1996). "Why individual vigilance increases as group size increases". Anim Behav. 51: 1077–1086. doi:10.1006/anbe.1996.0109. S2CID 53202810.
- ^ Lima, S (1995). "Back to the basics of anti-predatory vigilance: the group-size effect". Chování zvířat. 49 (1): 11–20. doi:10.1016/0003-3472(95)80149-9. S2CID 53205760.
- ^ Fritsches, KA; Brill, RW; Warrant, EJ (2005). "Warm Eyes Provide Superior Vision in Swordfishes" (PDF). Aktuální biologie. 15 (1): 55–58. doi:10.1016/j.cub.2004.12.064. PMID 15649365. S2CID 14070646.
- ^ Hopkin, Michael (2005). "Swordfish heat their eyes for better vision". Zprávy o přírodě. doi:10.1038/news050110-2.
- ^ Helfman et al, 2009, pp. 95–97.
- ^ Somiya, H (1980). "Fishes with Eye Shine: Functional Morphology of Guanine Type Tapetum Lucidum". Mar. Ecol. Prog. Ser. 2: 9–26. doi:10.3354/meps002009.
- ^ Johnson JA and Esser R (2009) "http://www.fishculturesection.org/Aquanotes/pdf/Aq_App_Note_1_April_2009.pdf Walleye Culture – Habituation to Feed in the Dark" Americká rybářská společnost, Aquaculture Application Note.
- ^ A b Douglas, Ron H.; Collin, Shaun P .; Corrigan, Julie (2002-11-15). "The eyes of suckermouth armoured catfish (Loricariidae, subfamily Hypostomus): pupil response, lenticular longitudinal spherical aberration and retinal topography" (PDF). Journal of Experimental Biology. The Journal of Experimental Biology. 205 (22): 3425–3433. PMID 12364396.
- ^ Yoshizawa, M.; Yamamoto, Y .; O'Quin, K. E.; Jeffery, W. R. (December 2012). "Evolution of an adaptive behavior and its sensory receptors promotes eye regression in blind cavefish". Biologie BMC. 10: 108. doi:10.1186/1741-7007-10-108. PMC 3565949. PMID 23270452.
- ^ A b Bone & Moore, 2008, pp. 418–422.
- ^ Bone & Moore, 2008, p. 311.
- ^ Atema, Jelle (1980) "Chemical senses, chemical signals, and feeding behavior in fishes" str. 57–101. In: Bardach, JE Fish behavior and its use in the capture and culture of fishes', The WorldFish Center, ISBN 978-971-02-0003-0.
- ^ Fields, RD, Fields, KD, Fields, MC (2007). "Semiconductor gel in shark sense organs?". Neurosci. Lett. 426 (3): 166–170. doi:10.1016/j.neulet.2007.08.064. PMC 2211453. PMID 17904741.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Brown BR (2010). "Temperature response in electrosensors and thermal voltages in electrolytes". J Biol Phys. 36 (2): 121–134. doi:10.1007/s10867-009-9174-8. PMC 2825305. PMID 19760113.
- ^ Johnsen, S (2005). "The physics and neurobiology of magnetoreception" (PDF). Recenze přírody Neurovědy. 6 (9): 703–712. doi:10.1038/nrn1745. PMID 16100517. S2CID 13996233. Archivovány od originál (PDF) dne 30.06.2007. Citováno 2011-12-18.
Reference
- Bone Q and Moore RH (2008) Biologie ryb Taylor & Francis Group. ISBN 978-0-415-37562-7.
- Helfman, G. S.; Collette, B. B .; Facey, D. E.; Bowen, B. W. (2009). The Diversity of Fishes: Biology, Evolution and Ecology. Wiley-Blackwell. ISBN 9781444311907.
- Moyle, PB a Cech, JJ (2004) Ryby, Úvod do ichtyologie. 5. vydání, Benjamin Cummings. ISBN 978-0-13-100847-2
Další čtení
- Arthur, Joseph; Nicol, Colin; Somiya, Hiroaki (1989). The eyes of fishes. Clarendon Press. ISBN 978-0-19-857195-7.
- Douglas, R. H. & Djamgoz, M. (eds) (1990) Vizuální systém ryb. Chapman and Hall, 526 pp.
- Lamb TD, Collin SP, Pugh EN (December 2007). "Evolution of the vertebrate eye: opsins, photoreceptors, retina and eye cup". Nat. Rev. Neurosci. 8 (12): 960–76. doi:10.1038/nrn2283. PMC 3143066. PMID 18026166. Ilustrace. Posouzení
- Lamb, TD (2011). "Evolution of the Eye" (PDF). Scientific American. 305 (1): 64–69. Bibcode:2011SciAm.305f..64L. doi:10.1038/scientificamerican0711-64. Archivovány od originál (PDF) dne 12. 12. 2013. Citováno 2013-04-28.
- Land, Michael F and Nilsson, Dan-Eric (2012) Zvířecí oči Oxford University Press. ISBN 9780199581146.
- Hagfish research has found the “missing link” in the evolution of the eye. Vidět: Nature Reviews Neuroscience.
- Nilsson, DE; Pelger, S (1994). "A pessimistic estimate of the time required for an eye to evolve" (PDF). Sborník Královské společnosti v Londýně B. 256 (1345): 53–58. Bibcode:1994RSPSB.256...53N. doi:10.1098/rspb.1994.0048. PMID 8008757. S2CID 13061351.
- Berlinski, David (2002) Has Darwin Met His Match? Page 34, The Vexing Eye (Letter). Komentář, 1 December 2002.
- Nilsson, Dan-E. "Beware of Pseudo-science: a response to David Berlinski's attack on my calculation of how long it takes for an eye to evolve". Citovat deník vyžaduje
| deník =(Pomoc) - Meyer-Rochow, VB; Coddington, PE (2003). "Eyes and vision of the New Zealand torrentfish Cheimarrhichthys fosteri von Haast (1874): histology, photochemistry and electrophysiology". In Val AL; Kapoor BG (eds.). Fish Adaptations. Oxford and IBH Publ. & M/s Sci. Publ., Enfield, New Hampshire (USA) & Plymouth (UK). pp. 337–383.</ref>
- "Evolution of the Eye" – video on Nilsson-Pelger model (scroll down)
Bibliografie
- Justin, Marshall, Karen L Carelton, Thomas Cronin , "Vision in marine ogranisms" , sciencedirect
- Makiko Kamijo, Mayuko Kawamura, Shoji Fukamachi, "Loss of red opsin genes relaxes sexual isolation between skin-colour variants of medaka", sciencedirect
externí odkazy

| O Ryba |  | ||||
|---|---|---|---|---|---|
| Anatomie a fyziologie | |||||
| Smyslové systémy | |||||
| Reprodukce | |||||
| Pohyb | |||||
| jiný chování | |||||
| Podle místo výskytu | |||||
| Jiné typy | |||||
| Komerční |
| ||||
| Hlavní skupiny | |||||
| Seznamy |
| ||||
| |||||