Tetratrikopeptid se opakuje - Tetratricopeptide repeat
| Tetratrikopeptid se opakuje | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||
| Identifikátory | |||||||||||
| Symbol | TPR_1 | ||||||||||
| Pfam | PF00515 | ||||||||||
| Pfam klan | CL0020 | ||||||||||
| InterPro | IPR001440 | ||||||||||
| SCOP2 | 1a17 / Rozsah / SUPFAM | ||||||||||
| CDD | cd00189 | ||||||||||
| |||||||||||
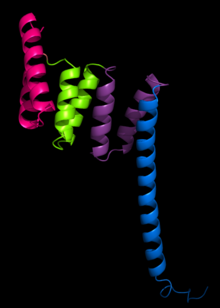
The opakování tetratrikopeptidu (TPR) je strukturální motiv. Skládá se z a degenerovat 34 aminokyselin tandemové opakování identifikovány v široké škále bílkoviny. Nachází se v tandemových polích 3–16 motivů,[1] které tvoří lešení zprostředkující interakce protein-protein a často sestavu multiproteinových komplexů. Tyto alfa-šroubovice pár se obvykle opakuje složit společně vytvořit jedinou, lineární solenoidová doména volal a TPR doména. Mezi proteiny s takovými doménami patří komplex podporující anafázi (APC) podjednotky cdc16, cdc23 a cdc27, NADPH oxidáza podjednotka p67-phox, vazba hsp90 imunofiliny, transkripční faktory, protein kináza R. (PKR), hlavní receptor pro import proteinu peroxisomální matrice PEX5, protein arginin methyltransferáza 9 (PRMT9) a mitochondriální importní proteiny.

Struktura
Struktura PP5 protein byla první struktura, která se měla určit. Struktura vyřešena Rentgenová krystalografie Das a kolegové ukázali, že motiv sekvence TPR byl složen z dvojice antiparalelních alfa šroubovic.[2] Struktura PP5 obsahovala 3 tandemové opakování TPR, které ukázaly, že postupná opakování TPR vytvořila alfa-šroubovici solenoid struktura.
Typická struktura TPR je charakterizována interakcemi mezi šroubovicemi A a B prvního motivu a šroubovice A 'dalšího TPR. Ačkoli se povaha takových interakcí může lišit, první dvě šroubovice motivu TPR mají obvykle úhel balení ~ 24 stupňů v rámci jediného motivu. Opakování více než tří motivů TPR generují pravoruký superhelix charakterizovaný jak konkávním, tak konvexním obličejem, jehož konkávní obličej je obvykle zapojen do vazby ligandu.[1] [3]

Pokud jde o sekvenci, TPR obsahuje směs malých a velkých hydrofobních zbytků, nicméně žádné polohy nejsou zcela neměnné. Existují však určité zbytky, které jsou obvykle konzervovány, včetně Tryptofanu 4, Leucinu 7, Glycinu 8, Tyrosinu 11, Alaninu 20, Fenylalaninu 24, Alaninu 27 a Prolinu 32. Z těchto 8 má alanin v pozicích 8, 20 a 27 tendenci být konzervovaný. Ostatní pozice mají silnější preference pro malé, velké nebo aromatické aminokyseliny než pro konkrétní zbytek. Mezi helixy hraje ochrana zbytků spíše strukturální roli s přítomnými zbytky štěpení šroubovice. Mezi sousedním TPR mají zbytky role se strukturálními i funkčními důsledky.[1]
PPR obsahující peptidy
Poskok
The Poskok adaptorový protein zprostředkovává asociaci molekulárních chaperonů Hsp70 a Hsp90. Obsahuje tři 3-TPR opakování, z nichž každé má vlastní peptidovou vazebnou specificitu. Je známo, že jeho doména TPR1 rozpoznává C-terminál Hsp70, zatímco TPR2 se váže na C-terminál Hsp90. Obě C-koncové sekvence končí motivem EEVD a povaha interakce je elektrostatická i hydrofobní.[1][4]
PEX5
The PEX5 protein je receptorem pro PTS1 (peroxisomální cílový signální tripeptid, který směruje proteiny do peroxisomů). Interaguje se signálem pomocí TPR motivů. Většina jeho kontaktů s C-terminálním tripeptidem PTS1 je v konkávní ploše TPR 1, 2 a 3.[5]
Neutrofilní cytosolický faktor 2
Neutrofilní cytosolický faktor 2 je nezbytný pro komplex NADPH oxidázy, který zase produkuje superoxidy v reakci na mikrobiální infekci. Vazba Rac GTPázy je klíčovým krokem do sestavení komplexu a TPR v jednotce phox zprostředkovávají sestavení multiproteinového komplexu působením vazebného lešení.[6]
Příklady
Lidské geny kódující proteiny obsahující tento motiv zahrnují:
- AAG2, ANAPC7
- BBS4
- CABIN1, CDC16, CDC23, CDC27, CNOT10, CTR9
- DNAJC3, DNAJC7, DYX1C1
- FAM10A4, FAM10A5, FKBP4, FKBP5, FKBP8, FKBPL
- GPSM1, GPSM2, GTF3C3
- IFIT1, IFIT1L, IFIT2, IFIT3, IFIT5, IFT140, IFT88
- KLC1, KLC2, KLC3, KLC4, KNS2
- LONRF2
- NARG1, NARG1L, NASP, NCF2, NFKBIL2, NOXA1, NPHP3
- OGT
- PEX5, PEX5L, PPID, PPP5C, PRPF6
- RANBP2, RANBP2L2, RANBP2L6, RAPSN, RGPD5, RGPD7, RPAP3
- SGTA, SGTB, SH3TC1, SH3TC2, SPAG1, SRP72, ST13, KROK1, STUB1, SUGT1
- TMTC1, TMTC2, TMTC3, TMTC4, TOMM34, TOMM70A
- TTC1, TTC3, TTC4, TTC5, TTC6, TTC7A, TTC7B, TTC8, TTC9C, TTC12, TTC13, TTC14, TTC15, TTC16, TTC17, TTC18, TTC21A, TTC21B, TTC22, TTC24, TTC25, TTC27, TTC28, TTC29, TTC30A, TTC30B, TTC31, TTC33, TTC37
- UNC45A, UNC45B, UTX, UTY
- WDTC1
- ZC3H7B
Reference
- ^ A b C d Blatch GL, Lässle M (listopad 1999). „Tetratrikopeptidová repetice: strukturální motiv zprostředkující interakce protein-protein“. BioEssays. 21 (11): 932–9. doi:10.1002 / (SICI) 1521-1878 (199911) 21:11 <932 :: AID-BIES5> 3.0.CO; 2-N. PMID 10517866.
- ^ Das AK, Cohen PW, Barford D (březen 1998). „Struktura tetratrikopeptidových opakování proteinové fosfatázy 5: důsledky pro interakce protein-protein zprostředkované TPR“. Časopis EMBO. 17 (5): 1192–9. doi:10.1093 / emboj / 17.5.1192. PMC 1170467. PMID 9482716.
- ^ Wilson CG, Kajander T, Regan L (leden 2005). "Krystalová struktura NlpI. Prokaryotický tetratrikopeptidový opakující se protein s globulárním záhybem". FEBS Journal. 272 (1): 166–79. doi:10.1111 / j.1432-1033.2004.04397.x. PMID 15634341.
- ^ Scheufler C, Brinker A, Bourenkov G, Pegoraro S, Moroder L, Bartunik H, Hartl FU, Moarefi I (duben 2000). "Struktura komplexů TPR doména-peptid: kritické prvky při sestavování multichaperonového stroje Hsp70-Hsp90". Buňka. 101 (2): 199–210. doi:10.1016 / S0092-8674 (00) 80830-2. PMID 10786835. S2CID 18200460.
- ^ Gatto GJ, Geisbrecht BV, Gould SJ, Berg JM (prosinec 2000). "Peroxisomální cílení rozpoznávání signálu 1 pomocí TPR domén lidského PEX5". Přírodní strukturní biologie. 7 (12): 1091–5. doi:10.1038/81930. PMID 11101887. S2CID 35168630.
- ^ Lapouge K, Smith SJ, Walker PA, Gamblin SJ, Smerdon SJ, Rittinger K (říjen 2000). "Struktura TPR domény p67phox v komplexu s Rac.GTP". Molekulární buňka. 6 (4): 899–907. doi:10.1016 / S1097-2765 (05) 00091-2. PMID 11090627.
Další čtení
- Lima Mde F, Eloy NB, Pegoraro C, Sagit R, Rojas C, Bretz T, Vargas L, Elofsson A, de Oliveira AC, Hemerly AS, Ferreira PC (18. listopadu 2010). „Genomický vývoj a složitost komplexu podporujícího anafázu (APC) v suchozemských rostlinách“. Biologie rostlin BMC. 10: 254. doi:10.1186/1471-2229-10-254. PMC 3095333. PMID 21087491.
- Das AK, Cohen PW, Barford D (březen 1998). „Struktura tetratrikopeptidových opakování proteinové fosfatázy 5: důsledky pro interakce protein-protein zprostředkované TPR“. Časopis EMBO. 17 (5): 1192–9. doi:10.1093 / emboj / 17.5.1192. PMC 1170467. PMID 9482716.
- Whitfield C, Mainprize IL (únor 2010). "TPR motivy: charakteristické znaky nového polysacharidového exportního lešení". Struktura. 18 (2): 151–3. doi:10.1016 / j.str.2010.01.006. PMID 20159460.
- Krachler AM, Sharma A, Kleanthous C (červenec 2010). „Samo-asociace TPR domén: Poučení z navrženého konsenzuálního TPR oligomeru“. Proteiny. 78 (9): 2131–43. doi:10,1002 / prot.22726. PMID 20455268.
- Schapire AL, Valpuesta V, Botella MA (září 2006). "TPR proteiny v signalizaci rostlinných hormonů". Signalizace a chování zařízení. 1 (5): 229–30. doi:10,4161 / psb.1.5.3491. PMC 2634123. PMID 19704665.
- Cortajarena AL, Regan L (květen 2006). „Vazba ligandu pomocí TPR domén“. Věda o bílkovinách. 15 (5): 1193–8. doi:10.1110 / ps.062092506. PMC 2242500. PMID 16641492.
- D'Andrea LD, Regan L (prosinec 2003). "TPR proteiny: univerzální šroubovice". Trendy v biochemických vědách. 28 (12): 655–62. CiteSeerX 10.1.1.313.8712. doi:10.1016 / j.tibs.2003.10.007. PMID 14659697.
- Goebl M, Yanagida M (květen 1991). "TPR snap helix: nový motiv proteinového opakování od mitózy po transkripci". Trendy v biochemických vědách. 16 (5): 173–7. doi:10.1016 / 0968-0004 (91) 90070-C. PMID 1882418.
externí odkazy
- Zdroj eukaryotického lineárního motivu třída motivů LIG_TPR
- Zdroj eukaryotického lineárního motivu třída motivů TRG_PTS1