Stria terminalis - Stria terminalis
| Stria terminalis | |
|---|---|
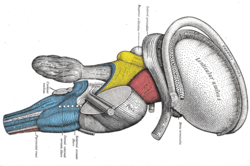 Pitva mozkového kmene. Boční pohled. (Stria terminalis označeno vpravo nahoře.) | |
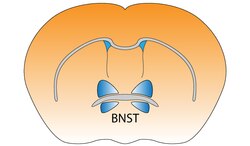 Jádro lůžka stria terminalis v myší mozek | |
| Detaily | |
| Identifikátory | |
| latinský | stria terminalis |
| NeuroNames | 286 |
| NeuroLex ID | birnlex_937 |
| TA98 | A14.1.09.275 |
| TA2 | 5592 |
| FMA | 61974 |
| Anatomické pojmy neuroanatomie | |
The stria terminalis (nebo terminální stria) je struktura v mozek sestávající z pásu vláken probíhajících podél postranního okraje komorového povrchu thalamus. Slouží jako hlavní výstupní cesta systému amygdala, stria terminalis probíhá od jeho centromediálního rozdělení k ventromediální jádro hypotalamu.
Anatomie
Stria terminalis pokrývá vynikající thalamostriate žíla, označující linii oddělení mezi thalamem a kádové jádro jak je vidět při hrubé pitvě komory mozku, při pohledu z lepšího aspektu.
Stria terminalis sahá od oblasti mezikomorová foramina do dočasný roh z boční komora, nesoucí vlákna z amygdala do septální jádra, hypotalamus a thalamické oblasti mozku. Také nese vlákna vyčnívající z těchto oblastí zpět do amygdaly.
Lůžkové jádro stria terminalis (BNST)
Činnost jádro lůžka stria terminalis koreluje s úzkostí v reakci na monitorování hrozeb.[1] Předpokládá se, že funguje jako předávací místo v rámci osa hypotalamus-hypofýza-nadledviny a regulovat jeho aktivitu v reakci na akutní stres.[2] Stresová reakce však souvisí s časem a BNST se neaktivuje pro kontextuální strach. To znamená, že náhlá děsivá situace, která je kratší než deset minut, neaktivuje BNST.[3] Rovněž se předpokládá, že podporuje inhibici chování v reakci na neznámé jedince, a to na základě vstupu od orbitofrontální kůra.[4] Ukázalo se, že dvoustranné narušení této cesty zmírňuje obnovení chování při hledání drog u hlodavců.[5]
Je známo, že toto jádro promítá inhibiční vlákna do laterálního hypotalamu a podílí se na kontrole krmení hlodavců. Optogenetický aktivace této inhibiční dráhy rychle vyvolala nenasytné stravovací chování u dobře krmených myší a optogenetická inhibice této dráhy snižuje příjem potravy i u hladovějících zvířat.[6]
Sexuální dimorfismus
Centrální dělení jádra lůžka stria terminalis (BSTc) je sexuálně dimorfní. V průměru je BSTc dvakrát větší u mužů než u žen a obsahuje dvojnásobný počet somatostatin neurony.[7] Vzorek šesti post mortem dlouhodobě léčených HRT trans ženy (muž-žena) bylo zjištěno, že mají v BSTc typický počet buněk pro ženu, zatímco a trans muž (žena-muž) bylo zjištěno, že mají mužské typické číslo.[8][9] Autoři (Jiang-Ning Zhou, Frank PM Kruijver, Dick Swaab ) také zkoumali subjekty s poruchami souvisejícími s hormony a nezjistili žádný vzorec mezi těmito poruchami a BSTc, zatímco jediný neléčený muž-žena transsexuál měl typický počet buněk pro ženy. Došli k závěru, že BSTc poskytuje důkaz neurobiologického základu genderová identita a navrhl, aby to bylo určeno před narozením.
Hormonální substituční léčba bylo prokázáno, že ovlivňuje velikost hypotalamu,[10] i když se to studie pokusila provést zahrnutím netranssexuálních mužských a ženských kontrol, které z různých lékařských důvodů zaznamenaly obrácení hormonů.[8] Tvrzení o neurobiologickém základě od narození bylo později zpochybněno, i když nebylo vyvráceno, následnou studií stejné skupiny, která zjistila, že sexuální dimorfismus BSTc není přítomen před dospělostí (přibližně 22 let) ačkoli transsexuálové uvádějí, že jsou si vědomi své genderové identity od dětství.[11]
Vzhledem k tomu, že neurony exprimující somatostatin typicky blokují dendritické vstupy do postsynaptického neuronu, čímž inhibují signály procházející souvisejícími strukturami, předpokládá se, že větší jádro lůžka stria terminalis nalezené u mužů (včetně transgender mužů) překvapivá odpověď u mužů a může být zodpovědný za vyšší výskyt specifických fóbií u žen a za možný zdroj stereotypu, že se ženy bojí myší.[12]
Aktivita receptoru oxytocinu v BNST je důležitá pro sociální uznání u potkanů. Samci i samice potkanů, kteří dostali mikroinjekci antagonisty oxytocinového receptoru, měli nižší skóre sociálního rozpoznávání než krysy, kterým byla podána injekce vehikula, a mikroinjekce oxytocinu do BNST zlepšily sociální paměť u samců, ale ne samic potkanů.[13]
Snížení velikosti jádra lůžka stria terminalis bylo pozorováno u pedofilní mužských pachatelů, kromě snížení pravé amygdaly, hypotalamu a abnormalit v souvisejících strukturách. Autoři navrhují, že dětské deficity u BNST a mediální amygdaly mohou způsobit inhibici sexuální dospělosti.[14]
Reference
- ^ Somerville L, Whalen P, Kelley W (2010). „Nucleus lidského lůžka Stria Terminalis indexuje monitorování hypervigilantních hrozeb“. Biol psychiatrie. 68 (5): 416–424. doi:10.1016 / j.biopsych.2010.04.002. PMC 2921460. PMID 20497902.
- ^ Choi D, Furay A, Evanson N, Ostrander M, Ulrich-Lai Y, Herman J (2007). „Nukleové jádro subregionů Stria Terminalis rozdílně reguluje činnost osy hypotalamus – hypofýza – nadledvin: důsledky pro integraci limbických vstupů.“. J. Neurosci. 27 (8): 2025–34. doi:10.1523 / JNEUROSCI.4301-06.2007. PMC 6673539. PMID 17314298.
- ^ Hammack SE, Todd TP, Kocho-Schellenberg M, Bouton ME (2015). „Role základního jádra Stria Terminalis při získávání kontextuálního strachu při dlouhých nebo krátkých intervalech kontextového šoku“. Behaviorální neurovědy. 129 (5): 673–678. doi:10.1037 / bne0000088. PMC 4586907. PMID 26348716.
- ^ Fox A, Shelton S, Oakes T, Converse A, DavidsonR, Kalin N (2010). „Orbitofrontální léze mozkové kůry mění aktivitu související s úzkostí v jádru primátového lůžka Stria Terminalis“. J. Neurosci. 30 (20): 7023–27. doi:10.1523 / JNEUROSCI.5952-09.2010. PMC 2915894. PMID 20484644.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Suzanne Erb · Natalina Salmaso · Demetra Rodaros Jane Stewart (2001). „Role dráhy obsahující CRF z centrálního jádra amygdaly do jádra stria terminalis při stresem navozeném obnovení kokainu u potkanů“. Psychofarmakologie. 158 (4): 360–65. doi:10,1007 / s002130000642. PMID 11797056. S2CID 23284158.
- ^ Jennings, Joshua H .; Rizzi, Giorgio; Stamatakis, Alice M .; Ung, Randall L .; Stuber, Garret D. (2013-09-27). „Inhibiční obvodová architektura krmení laterálních hypotalamů orchestruje“. Věda. 341 (6153): 1517–1521. doi:10.1126 / science.1241812. ISSN 0036-8075. PMC 4131546. PMID 24072922.
- ^ Swaab D (2007). "Sexuální diferenciace mozku a chování". Best Pract Res Clin Endocrinol Metab. 21 (3): 431–44. doi:10.1016 / j.beem.2007.04.003. PMID 17875490.
- ^ A b Zhou J, Hofman M, Gooren L, Swaab D (1995). „Rozdíl pohlaví v lidském mozku a jeho vztah k transsexualitě“. Příroda. 378 (6552): 68–70. Bibcode:1995 Natur.378 ... 68Z. doi:10.1038 / 378068a0. hdl:20.500.11755 / 9da6a0a1-f622-44f3-ac4f-fec297a7c6c2. PMID 7477289. S2CID 4344570.
- ^ Kruijver F, Zhou J, Pool C, Hofman M, Gooren L, Swaab D (2000). „Transsexuálové muž-žena mají počet ženských neuronů v limbickém jádru“. J. Clin. Endokrinol. Metab. 85 (5): 2034–41. doi:10.1210 / jcem.85.5.6564. PMID 10843193.
- ^ Hulshoff Pol HE, Cohen-Kettenis PT, Van Haren NE, Peper JS, Brans RG, Cahn W a kol. (2006). „Změna pohlaví mění váš mozek: Vliv testosteronu a estrogenu na strukturu mozku dospělého člověka“. Evropský žurnál endokrinologie. 155 (Příloha 1): S107 – S114. doi:10.1530 / eje.1.02248.
- ^ Chung W, De Vries G, Swaab D (2002). „Sexuální diferenciace jádra lůžka stria terminalis u lidí může trvat až do dospělosti“. J. Neurosci. 22 (3): 1027–33. doi:10.1523 / JNEUROSCI.22-03-01027.2002. PMC 6758506. PMID 11826131.
- ^ Cameron, Alasdair (2004). Crash Course Psychiatry. Elsevier Ltd. ISBN 978-0-7234-3340-8.
- ^ Dumais, KM; Alonso, AG; Immormino, MA; Bredewold, R; Veenema, AH (únor 2016). „Zapojení oxytocinového systému do jádra postele stria terminalis do regulace sociálního uznání specifické pro pohlaví“. Psychoneuroendokrinologie. 64: 79–88. doi:10.1016 / j.psyneuen.2015.11.007. PMC 4698213. PMID 26630388.
- ^ Schiltz K, Witzel J, Northoff G, Zierhut K, Gubka U, Fellman H, Kaufmann J, Tempelmann C, Wiebking C, Bogerts B (2007). „Mozková patologie u pedofilních pachatelů: Důkazy o zmenšení objemu v pravé amygdale a souvisejících diencefalických strukturách“. Archiv obecné psychiatrie. 64 (6): 737–746. doi:10.1001 / archpsyc.64.6.737. PMID 17548755.
externí odkazy
{kind=link}
| Kontrolní úřad |
|---|