Putamen - Putamen - Wikipedia
| Putamen | |
|---|---|
 Putamen (červeně) zobrazený v mozku | |
 Koronální část mozku prostřednictvím mezilehlé hmoty třetí komora. (Putamen označen nahoře.) | |
| Detaily | |
| Část | Hřbetní striatum |
| Identifikátory | |
| Pletivo | D011699 |
| NeuroNames | 230 |
| NeuroLex ID | birnlex_809 |
| TA98 | A14.1.09.507 |
| TA2 | 5566 |
| FMA | 61834 |
| Anatomické pojmy neuroanatomie | |
The putamen (/strjutˈeɪmən/; z latinský, což znamená "kostka") je kulatá struktura umístěná na základně přední mozek (telencephalon ). Putamen a kádové jádro společně tvoří hřbetní striatum. Je to také jedna ze struktur, které tvoří bazální jádra. Putamen je různými cestami spojen s substantia nigra, globus pallidus, klaustrum a thalamus, kromě mnoha oblastí mozkové kůry. Primární funkcí putamenu je regulace pohybů v různých fázích (např. Příprava a provedení) a ovlivňování různých typů učení se. Zaměstnává to GABA, acetylcholin, a enkefalin vykonávat své funkce. Putamen také hraje roli v degenerativní neurologické poruchy, jako Parkinsonova choroba.
Dějiny
Slovo „putamen“ je z latinský, s odkazem na to, co „odpadá při prořezávání“, z „putare“, což znamená „prořezávat, přemýšlet nebo uvažovat“.[1]
Až donedávna se většina výzkumů MRI zaměřovala široce na bazální ganglia jako celek, a to z různých důvodů (např. Rozlišení obrazu, vzácnost izolovaného infarktu nebo krvácení v putamenu atd.). Bylo však provedeno mnoho studií o bazálních gangliích a příslušných vztazích chování a mozku. V 70. letech první záznamy jednotlivých jednotek byly hotové s opice monitorování bledý neuron činnost související s pohybem. Od té doby byly zahájeny rozsáhlejší výzkumné metody sledování neuronů, stimulace a zobrazování (např. fMRI, DWI ), které umožňují vyšetřování putamenů.
Anatomie

Putamen je struktura v přední mozek. Spolu s kádové jádro tvoří hřbetní striatum. Kaudát a putamen obsahují stejné typy neuronů a obvodů - mnoho neuroanatomů považuje dorzální striatum za jedinou strukturu, rozdělenou na dvě části velkým vláknitým traktem, vnitřní kapsle, procházející středem. Putamen spolu s globus pallidus, tvoří čočkovité jádro. Putamen je nejvzdálenější část bazální ganglia. Jedná se o skupinu jader v mozku, která jsou propojena s mozková kůra, thalamus, a mozkový kmen. Bazální ganglia zahrnují hřbetní striatum, substantia nigra, nucleus accumbens a subtalamické jádro.
v savci, bazální ganglia jsou spojena s ovládání motoru, poznání, emoce, učení se a obecné funkce domény důležité pro výkonné fungování i podpora jazyků specifických pro doménu. Bazální ganglia jsou umístěna bilaterálně a mají rostrální a ocasní divize. Putamen se nachází v rostrální divizi jako součást striatum. Bazální ganglia přijímají vstup z mozkové kůry prostřednictvím striata.
Putamen je propojen s následujícími strukturami:

Tento popis je primitivní a téměř nevyčerpává ani základní zavedené obvody putamenu. Kortiko-subkortikokortikální obvody s putaminálním postižením jsou husté a komplikované, skládající se z široké škály axonálních, dendritických, chemických, aferentních a eferentních substrátů. Výstupy putamenu jsou vysoce arborizované napříč výstupními strukturami a kortikální eferenty vycházejí z vrstev III-VI kůry, v závislosti na gyri a umístění v putamenu. Topografická organizace putamenu kombinuje následující prvky: přední a zadní funkční a somatotopické přechody, laterální až střední funkční a somatotopické přechody, difúzní koncový výstup, nepravidelný lokalizovaný koncový výstup, oddělené terminály od sousedních oblastí, jemně interdigiované terminály od distálních kortikální oblasti zdánlivě překrývajícím se způsobem[Citace je zapotřebí ].
Kádové jádro
Caudate pracuje s putamenem, aby přijímal vstup z mozková kůra. Souhrnně je lze považovat za „vchod“ do bazálních ganglií. Projekce putamenu se dostávají do kaudátu přímo přes kaudolentikulární šedé můstky. Putamen a caudate jsou společně spojeny s substantia nigra, nicméně caudate vystupuje hustěji k substantia nigra pars reticulata, zatímco putamen posílá více aferentů do interní globus pallidus.
Substantia nigra
Substantia nigra obsahuje dvě části: substantia nigra pars compacta (SNpc) a substantia nigra pars reticulata (SNpr). SNpc získává vstup z putamenu a caudate a odesílá informace zpět. SNpr také získává vstupy od putamenu a caudate. Posílá však vstup mimo bazální ganglia k ovládání hlavy a pohyby očí. SNpc produkuje dopamin, který je zásadní pro pohyby. SNpc je část, která se zdegeneruje Parkinsonova choroba.[2]
Globus pallidus
Globus pallidus obsahuje dvě části: globus pallidus pars externa (GPe) a globus pallidus pars interna (GPi). Obě oblasti získávají vstup od putamenu a caudátu a komunikují s subthalamickým jádrem. Většinou však GPi odesílá GABAergní inhibiční výstup do thalamu. GPi také odesílá projekce do částí středního mozku, u nichž se předpokládá, že ovlivňují kontrolu držení těla.[2]
Fyziologie
Typy cest
Putamen (a striatum obecně) má četné paralelní obvody, které umožňují komunikační kortiko-subkortiko-kortikální smyčky. Ty byly obecně popsány jako přímé, nepřímé a hyper přímé cesty. GABAergické projekce putamenu mají inhibiční účinek na thalamus. Thalamické projekce z centromediánových a parafascikulárních jader mají excitační účinek na putamen. Na rozdíl od thalamu, který má širokou vzájemnou konektivitu, jsou kortikální projekce s putamenem aferentní, a tak posílají informace na rozdíl od jejich přijímání. Kortikální komunikace se provádí pomocí vícevláknových drah, jak je uvedeno výše (tj. Prostřednictvím jiných subkortikálních struktur).
Dopamin
Dopamin je neurotransmiter, který má dominantní roli v putamenu, většina je dodávána z substantia nigra. Když buněčné tělo a neuron (v jádrech putamen nebo caudate) vystřelí an akční potenciál, dopamin se uvolňuje z presynaptický terminál. Vzhledem k tomu, projekce z putamen a caudate jádra modulovat the dendrity substantia nigra ovlivňuje dopamin substantia nigra, která ovlivňuje plánování motorů. Stejný mechanismus je součástí drogová závislost. Za účelem kontroly množství dopaminu v synaptická štěrbina a množství vazby dopaminu na post synaptické terminály, fungují presynaptické dopaminergní neurony zpětné vychytávání přebytek dopaminu.
Jiné neurotransmitery
Putamen také hraje roli v modulace jiných neurotransmiterů. Uvolňuje GABA, enkefalin, látka P, a acetylcholin. Přijímá serotonin a glutamát.
Funkce: motorika
Putamen je propojen s mnoha dalšími strukturami a ve spojení s nimi ovlivňuje mnoho typů motorického chování. Patří mezi ně plánování motorů, učení a provádění,[3] příprava motoru,[4] specifikování amplitud pohybu,[5] a pohybové sekvence.[6]
Někteří neurologové předpokládají, že putamen také hraje roli při výběru pohybu (např. Tourettův syndrom ) a „automatický“ výkon dříve naučených pohybů (např. Parkinsonova choroba ).[7]
V jedné studii bylo zjištěno, že putamen řídí pohyb končetin. Cílem této studie bylo zjistit, zda konkrétní aktivita buněk putamenu primátů souvisí se směrem pohybu končetin nebo se základním vzorcem svalové aktivity. Dvě opice byly vycvičeny k provádění úkolů, které zahrnovaly pohyb břemen. Úkoly byly vytvořeny tak, aby bylo možné odlišit pohyb od svalové aktivity. Neurony v putamenu byly vybrány pro monitorování, pouze pokud souvisely jak s úkolem, tak s pohyby paží mimo úkol. Ukázalo se, že 50% neuronů, které byly sledovány, souviselo se směrem pohybu, nezávisle na zátěži.[8]
Byla provedena další studie, která zkoumala rozsah a rychlost pohybu pomocí PET mapování regionálního průtoku krve mozkem u 13 lidí. Pohybové úkoly byly prováděny pomocí a joystick - řízené kurzor. Statistické testy byly provedeny pro výpočet rozsahu pohybů a toho, s kterými oblastmi mozku pohyby souvisejí. Bylo zjištěno, že „rostoucí rozsah pohybu byl spojen s paralelním nárůstem rCBF v bilaterálních bazálních gangliích (BG; putamen a globus pallidus) a ipsilaterálním mozečku.“ To nejen ukazuje, že putamen ovlivňuje pohyb, ale také ukazuje, že putamen se integruje do jiných struktur za účelem plnění úkolů.[9]
Byla provedena jedna studie s cílem konkrétně prozkoumat, jak bazální ganglia ovlivňují učení sekvenčních pohybů. Dvě opice byly trénovány, aby postupně stiskly řadu knoflíků. Použité metody byly navrženy tak, aby dokázaly sledovat dobře naučené úkoly versus nové úkoly. Muscimol byl injikován do různých částí bazálních ganglií a bylo zjištěno, že „učení nových sekvencí se stalo nedostatečným po injekcích do předního kaudátu a putamenu, ale nikoli do středního-zadního putamenu“. To ukazuje, že různé oblasti EU striatum jsou využívány při provádění různých aspektů učení sekvenčních pohybů.[10]
Role v učení
V mnoha studiích se ukázalo, že putamen hraje roli v mnoha druzích učení. Některé příklady jsou uvedeny níže:
Posílení a implicitní učení
Spolu s různými druhy pohybu ovlivňuje putamen také posilování učení a implicitní učení.[11]
Posílení učení interaguje s prostředím a stravovacími akcemi, aby maximalizoval výsledek. Implicitní učení je pasivní proces, při kterém jsou lidé vystaveni informacím a získávají znalosti prostřednictvím expozice. Ačkoli přesné mechanismy nejsou známy, je zřejmé, že zde hraje klíčovou roli dopamin a tonicky aktivní neurony. Tonicky aktivní neurony jsou cholinergní interneurony že oheň po celou dobu trvání stimulu a oheň asi 0,5–3 impulsy za sekundu. Fázové neurony jsou opakem a akční potenciál aktivují pouze při pohybu.[12]
Kategorie učení
Jedna konkrétní studie používala pacienty s fokálními lézemi na bazálních gangliích (konkrétně putamen) kvůli mrtvice abych mohl studovat kategorie učení. Výhodou použití těchto typů pacientů je, že dopaminergní projekce do prefrontální kůra je větší pravděpodobnost, že budou neporušené. U těchto pacientů je také snazší spojit fungování specifických mozkových struktur, protože léze se vyskytuje pouze na určitém místě. Cílem této studie bylo zjistit, zda tyto léze ovlivňují nebo ne ovlivňují učení úkolů založené na pravidlech a integraci informací. Úkoly založené na pravidlech se učí prostřednictvím testování hypotéz v závislosti na pracovní paměti. Úkoly integrace informací jsou úkoly, u nichž je přesnost maximalizována, když jsou informace ze dvou zdrojů integrovány v předrozhodovací fázi, která se řídí procedurálním systémem.
Sedm účastníků s bazálními gangliemi léze v experimentu byly použity spolu s devíti účastníky kontroly. Je důležité si uvědomit, že kaudát nebyl ovlivněn. Účastníci byli testováni na každý typ učení během samostatných sezení, aby si informační procesy navzájem nerušily. Během každé relace seděli účastníci před obrazovkou počítače a zobrazovaly se různé řádky. Tyto linie byly vytvořeny pomocí randomizační techniky, kdy byly odebrány náhodné vzorky z jedné ze čtyř kategorií. Pro testování na základě pravidel byly tyto vzorky použity ke konstrukci linií různé délky a orientace, které spadaly do těchto čtyř samostatných kategorií. Poté, co byl stimul zobrazen, byly subjekty vyzvány, aby stiskly 1 ze 4 tlačítek, aby označily, do které kategorie linka patřila. Stejný proces byl opakován pro úkoly integrace informací a byly použity stejné podněty, kromě toho, že hranice kategorií byly otočeny o 45 °. Tato rotace způsobí, že subjekt integruje kvantitativní informace o řádku před určením, v jaké kategorii se nachází.
Bylo zjištěno, že subjekty v experimentální skupině byly narušeny při plnění úkolů založených na pravidlech, ale nikoli v oblasti integrace informací. Po statistickém testování se také předpokládalo, že mozek začal používat techniky integrace informací k řešení učebních úkolů založených na pravidlech. Jelikož úlohy založené na pravidlech používají systém mozku pro testování hypotéz, lze vyvodit závěr, že systém mozku pro testování hypotéz byl poškozen / oslaben. Je známo, že caudate a pracovní paměti jsou součástí tohoto systému. Proto se potvrdilo, že putamen je zapojen do učení kategorie, konkurence mezi systémy, zpracování zpětné vazby v úlohách založených na pravidlech a je zapojen do zpracování pre-frontálních regionů (které se týkají pracovní paměti a výkonného fungování) . Nyní je známo, že nejen bazální ganglia a kaudát ovlivňují učení kategorií.[13]
Role v „nenávistném okruhu“
Předběžné studie naznačují, že putamen může hrát roli v tzv.nenávist obvod "mozku. Nedávná studie byla provedena v Londýn katedrou buněčné a vývojové biologie v Brně University College v Londýně. An fMRI bylo provedeno u pacientů, zatímco si prohlíželi obraz lidí, které nenáviděli, a lidí, kteří byli „neutrální“. Během experimentu bylo pro každý obrázek zaznamenáno „skóre nenávisti“. Aktivita v subkortikálních oblastech mozku znamenala, že „nenávistný okruh“ zahrnuje putamen a ostrov. Předpokládalo se, že „putamen hraje roli při vnímání opovržení a hnus, a mohou být součástí motorový systém to je mobilizováno k přijetí opatření. “Bylo také zjištěno, že míra aktivity v nenávistném okruhu koreluje s mírou nenávisti, kterou osoba prohlásí, což by mohlo mít právní důsledky týkající se škodlivých trestných činů.[14]
Role u transgenderových jedinců
Bylo zjištěno, že putamen má významně větší množství šedé hmoty u transgenderových jedinců mužů než žen ve srovnání s putamenem typického muže s cisgenderem. To možná naznačuje, že mezi nimi může existovat zásadní rozdíl ve složení mozku trans ženy a cis muži.[15]
Patologie
Parkinsonova choroba
Po objevení funkce putamenu bylo neurologům zřejmé, že putamen a další části bazálních ganglií hrají důležitou roli v Parkinsonova choroba a další nemoci, které zahrnují degeneraci neurony.[16]
Parkinsonova choroba je pomalá a stálá ztráta dopaminergních neuronů u substantia nigra pars compacta. v Parkinsonova choroba putamen hraje klíčovou roli, protože jeho vstupy a výstupy jsou propojeny s substantia nigra a globus pallidus. U Parkinsonovy nemoci aktivita v přímých cestách k vnitřnímu globus pallidus klesá a aktivita v nepřímých cestách k vnějšímu globus pallidus se zvyšuje. Proto mají Parkinsonovi pacienti třes a mají potíže s prováděním dobrovolných pohybů. Rovněž bylo poznamenáno, že Parkinsonovi pacienti mají s plánováním motorů obtížné období.
Další nemoci a poruchy
S putamenem jsou spojeny následující nemoci a poruchy:
- Kognitivní pokles v Alzheimerova choroba[17]
- Huntingtonova choroba
- Wilsonova nemoc
- Demence s Lewyho těly
- Kortikobazální degenerace
- Tourettův syndrom
- Schizofrenie
- Deprese
- Porucha pozornosti s hyperaktivitou[18]
- Chorea
- Obsedantně kompulzivní porucha[19][20]
- Kernicterus
- jiný úzkostné poruchy[20]
U jiných zvířat
Putamen u lidí je strukturou a funkcí relativně podobný ostatním zvířatům. Proto bylo mnoho studií putamenu provedeno na zvířatech (opice, krysy, kočky atd.), stejně jako lidé. Mezidruhové variace jsou však skutečně pozorovány u savců a byly zdokumentovány pro putaminální konektivitu bílé hmoty. Variace souvisí primárně se strukturálními vzory konektivity, zatímco principy somatotopické organizace jsou zachovány. Výzkum primátů od 80. let do současnosti prokázal, že kortikální oblasti ve vztahu k poznání vyššího řádu primárně posílají aferentní neurony do rostální části putamenu, zatímco zbytek této struktury u primátů slouží primárně senzoromotorickým funkcím a je hustě propojené s primární a doplňkovou motorickou oblastí.
Další obrázky

Koronální část mozku skrz přední komisura.

Horizontální část pravé mozkové hemisféry.

Mozek

Frontální (koronální) část lidského mozku

Vodorovný řez obrazem MRI zobrazujícím putamen. Lze také vidět další jádra bazálních ganglií (jádro caudate a globus pallidus).

Putamen

Putamen
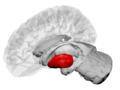
Putamen spolu s dalšími subkortikálními strukturami








Viz také
Reference
- ^ http://latin-dictionary.net/search/latin/putare
- ^ A b Alexander GE; Crutcher MD (červenec 1990). "Funkční architektura obvodů bazálních ganglií: neurální substráty paralelního zpracování". Trendy v neurovědách. 13 (7): 266–71. doi:10.1016 / 0166-2236 (90) 90107-L. PMID 1695401.
- ^ DeLong MR; Alexander GE; Georgopoulos AP; Crutcher MD; Mitchell SJ; Richardson RT (1984). "Role bazálních ganglií při pohybech končetin". Neurobiologie člověka. 2 (4): 235–44. PMID 6715208.
- ^ Alexander GE; Crutcher MD (červenec 1990). "Příprava na pohyb: neurální reprezentace zamýšleného směru ve třech motorických oblastech opice". Journal of Neurophysiology. 64 (1): 133–50. doi:10.1152 / jn.1990.64.1.133. PMID 2388061.
- ^ Delong MR; Georgopoulos AP; Crutcher MD; Mitchell SJ; Richardson RT; Alexander GE (1984). „Funkční organizace bazálních ganglií: příspěvky studií záznamu jednotlivých buněk“. Ciba Found Symp. 107: 64–82. PMID 6389041.
- ^ Marchand WR; Lee JN; Thatcher JW; Hsu EW; Rashkin E; Suchy Y; Chelune G; Starr J; Barbera SS (11. června 2008). "Putamenova koaktivace během provádění motorického úkolu". NeuroReport. 19 (9): 957–960. doi:10.1097 / WNR.0b013e328302c873. PMID 18521000.
- ^ Griffiths PD; Perry RH; Crossman AR (14. března 1994). „Podrobná anatomická analýza neurotransmiterových receptorů v putamenu a caudátu u Parkinsonovy choroby a Alzheimerovy choroby“. Neurovědy Dopisy. 169 (1–2): 68–72. doi:10.1016/0304-3940(94)90358-1. PMID 8047295.
- ^ Crutcher MD; DeLong MR (1984). „Jednobuněčné studie putamenu primátů. II. Vztahy ke směru pohybu a vzorci svalové aktivity“. Exp. Brain Res. 53 (2): 244–58. doi:10.1007 / bf00238154. PMID 6705862.
- ^ Turner RS; Desmurget M; Grethe J; Crutcher MD; Grafton ST (prosinec 2003). „Motorové obvody zprostředkující řízení rozsahu a rychlosti pohybu“. Journal of Neurophysiology. 90 (6): 3958–66. doi:10.1152 / jn.00323.2003. PMID 12954606.
- ^ Miyachi S; Hikosaka O; Miyashita K; Kárádi Z; Rand MK (červen 1997). "Diferenciální role opičího striata v učení postupného pohybu rukou". Exp. Brain Res. 115 (1): 1–5. doi:10.1007 / PL00005669. PMID 9224828.
- ^ Packard MG; Knowlton BJ (2002). "Učení a paměťové funkce bazální ganglií". Annu Rev Neurosci. 25 (1): 563–93. doi:10.1146 / annurev.neuro.25.112701.142937. PMID 12052921.
- ^ Yamada H; Matsumoto N; Kimura M (7. dubna 2004). „Tonicky aktivní neurony v jádru primátu caudate a putamen odlišně kódují instruované motivační výsledky akce“. Journal of Neuroscience. 24 (14): 3500–10. doi:10.1523 / JNEUROSCI.0068-04.2004. PMID 15071097.
- ^ Ell SW; Marchant NL; Ivry RB (2006). „Ohniskové léze putamen narušují učení v pravidlech založených na pravidlech, ale nikoli v úlohách kategorizace integrace informací“. Neuropsychologie. 44 (10): 1737–51. doi:10.1016 / j.neuropsychologia.2006.03.018. PMID 16635498.
- ^ Zeki S; Romaya JP (2008). Lauwereyns, Jan (ed.). „Neural Corelates of Hate“. PLOS ONE. 3 (10): e3556. doi:10,1371 / journal.pone.0003556. PMC 2569212. PMID 18958169.
- ^ Luders E; Sánchez FJ; Gaser C; et al. (Červenec 2009). „Regionální variace šedé hmoty v transsexualismu muž-žena“. NeuroImage. 46 (4): 904–7. doi:10.1016 / j.neuroimage.2009.03.048. PMC 2754583. PMID 19341803.
- ^ DeLong MR; Wichmann T (leden 2007). "Obvody a poruchy bazálních ganglií". Oblouk. Neurol. 64 (1): 20–4. doi:10.1001 / archneur.64.1.20. PMID 17210805.
- ^ de Jong LW; van der Hiele K; Veer IM; Houwing JJ; Westendorp RG; Bollen EL; de Bruin PW; Middelkoop HA; van Buchem MA; van der Grond J (prosinec 2008). „Silně snížené objemy putamenu a thalamu u Alzheimerovy choroby: studie MRI“. Mozek. 131 (12): 3277–85. doi:10.1093 / mozek / awn278. PMC 2639208. PMID 19022861.
- ^ Martin H. Teicher; Carl M. Anderson; Ann Polcari; Carol A. Glod; Luis C. Maas; Perry F. Renshaw (2000). „Funkční deficity v bazálních gangliích u dětí s poruchou pozornosti / hyperaktivity ukázané pomocí funkční relaxometrie pomocí magnetické rezonance“. Přírodní medicína. 6 (12): 470–3. doi:10.1038/74737. PMID 10742158.
- ^ Radua, Joaquim; Mataix-Cols, David (listopad 2009). „Voxelová metaanalýza změn šedé hmoty u obsedantně-kompulzivní poruchy“. British Journal of Psychiatry. 195 (5): 393–402. doi:10,1192 / bjp.bp.108,055046. PMID 19880927.
- ^ A b Radua, Joaquim; van den Heuvel, Odile A .; Surguladze, Simon; Mataix-Cols, David (5. července 2010). „Metaanalytické srovnání studií morfometrie založených na voxelech u obsedantně-kompulzivní poruchy vs. jiných úzkostných poruch“. Archiv obecné psychiatrie. 67 (7): 701–711. doi:10.1001 / archgenpsychiatry.2010.70. PMID 20603451.
externí odkazy
- Obarvené obrázky mozkových řezů, které obsahují „putamen“ na Projekt BrainMaps
- „Anatomické schéma: 13048 000-2“. Roche Lexicon - ilustrovaný navigátor. Elsevier. Archivovány od originál dne 22. 7. 2012.
- Schéma na uni-tuebingen.de
- Atlas obrázek: eye_38 na zdravotnickém systému University of Michigan - „Vizuální cesta zdola“
{kind=link}
| Kontrolní úřad |
|---|