Pretektální oblast - Pretectal area
| Pretektální oblast | |
|---|---|
| Detaily | |
| Část | Střední mozek |
| Díly | přední pretektální jádro, mediální pretektální jádro, jádro optického traktu, olivové pretektální jádro, zadní pretektální jádro, posterior limitans, komisurální pretektální oblast |
| Identifikátory | |
| latinský | oblast praetectalis |
| Pletivo | D066250 |
| NeuroNames | 467 |
| NeuroLex ID | nlx_59721 |
| TA98 | A14.1.08.505 A14.1.08.506 |
| TA2 | 5739 |
| FMA | 62402 |
| Anatomické pojmy neuroanatomie | |
The pretektální oblastnebo pretectum, je střední mozek struktura složená ze sedmi jader a zahrnuje část subkortikálního vizuálního systému. Prostřednictvím vzájemných bilaterálních projekcí z EU sítnice, podílí se primárně na zprostředkování behaviorálních reakcí na akutní změny okolního světla, jako je pupilární světelný reflex, optokinetický reflex a dočasné změny cirkadiánní rytmus.[1][2][3][4][5] Kromě role pretekta ve vizuálním systému bylo zjištěno, že přední pretektální jádro zprostředkovává somatosenzorický a nociceptivní informace.[6][7]
Umístění a struktura
Pretectum je dvoustranná skupina vysoce propojených jader umístěných poblíž křižovatky střední mozek a přední mozek.[8] Pretectum je obecně klasifikován jako struktura středního mozku, ačkoli kvůli jeho blízkosti předního mozku je někdy klasifikován jako součást kaudálního diencephalonu (předního mozku).[9] V rámci obratlovců, pretektum je umístěno přímo před superior colliculus a za thalamus. Nachází se nad periaqueductal šedá a jádro zadní komisury.[10]
V rámci pretectum bylo identifikováno několik jader, i když je obtížné definovat jejich hranice a debata o tom, které regiony by měly být zahrnuty, a jejich přesná jména.[1][10][11] Pět primárních jader je: olivární pretektální jádro (ON), jádro optického traktu (NOT) a přední (NPA), mediální (NPM) a zadní (NPP) pretektální jádra. NOT se skládá z relativně velkých buněk a je umístěn mezi superior colliculi. ON je umístěn mediálně k NOT a má ocas, který sahá mezi NOT a NPP, který je ventrální k ON.[10] Byly také identifikovány dvě další jádra: posterior limitans (PLi) a commisural pretectal area (CPA).[12] Zatímco tyto dvě oblasti nebyly zkoumány ve stejném rozsahu jako pět primárních jader, výzkum ukázal, že jak PLi, tak CPA přijímají sítnicový vstup, což naznačuje roli při zpracování vizuálních informací.[13]
Vstupy
Prefektum je významné binokulární vstup od fotocitlivé gangliové buňky v sítnici. U primátů jsou tyto aferenty dvoustranné[14] zatímco u hlodavců vyčnívají z kontralaterální sítnice. Většina těchto projekcí retino-pretectal jde do stavu ON a NOT[14] zatímco ostatní pretektální jádra dostávají menší retinální vstup u savců, včetně zadních, středních a předních pretektálních jader.[1][10][15][16]
NOT přijímá vstup z několika oblastí. Z thalamu NE přijímá inhibiční projekce GABA -produkující neurony v ipsilaterálním boční geniculate jádro a dvoustranné intergeniculate letáky. Ipsilaterální povrchní suprachiasmatické jádro a mediální, dorzální a laterální terminální jádra ve středním mozku vyčnívají do NOT. Vlákna také promítají do NOT z nucleus prepositus hypoglossi v míše, kontralaterální NENÍ a z různých kortikálních oblastí.[1][17]
Výstupy
Mnoho pretektálních jader sdílí cíle eferentních projekcí. Všechna pretektální jádra, s výjimkou ON, vyčnívají do jader v thalamu, subthalamu, superior colliculus, retikulární formace, pons, a nižší oliva.[10] Jak ON, tak CPA mají eferentní projekce k Edinger-Westphalovo jádro. NPP i NPA vyčnívají do pulvinaru, postranního zadního jádra thalamu a několika precerebelárních jader.[1]
NOT má eferentní projekce do zonální incerty subthalamu, několik jader pons, dřeně, intralaminární jádra, střední mozek a hřbetní a ventrální thalamová jádra. Jeho dvoustranné inhibiční projekce k doplňkovému optickému systému zahrnují připojení k laterálním a mediálním terminálním jádrům. Projekce do subthalamu jsou zaměřeny na laterální geniculární jádro a pulvinar. NEprojektuje bilaterálně s superior colliculus, i když se ipsilaterální spojení jeví jako dominantnější. Kromě těchto projekcí NENÍ promítá do vestibulární a vestibulocerebellar předat jádra.[1]
Funkce
Jako součást subkortikálního vizuálního systému reagují neurony v pretektálních jádrech na různé intenzity osvětlení a jsou primárně zapojeny do zprostředkování reakcí chování bez vědomí na akutní změny světla. Obecně tyto reakce zahrnují iniciaci optokinetických reflexů, i když pretektum může také regulovat nocicepci a REM spánek.[12]
Pupilární světelný reflex
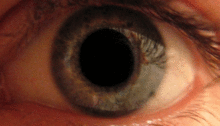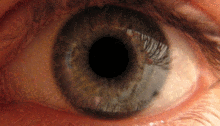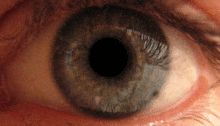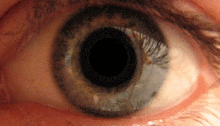
Pupilární světelný reflex je zprostředkován pretektem.[2] Tento reflex je zodpovědný za zúžení zornic při vstupu světla do oka. Několik pretektálních jader, zejména ON, přijímá informace o osvětlení z ipsilaterální sítnice optickým traktem. Je známo, že jádra v zapnutém stavu se postupně zvyšují v aktivaci v reakci na zvyšující se úrovně osvětlení. Tato informace je poté předána přímo do Edinger-Westphalova jádra, které pokračuje předáním příkazu k zúžení žáků na pupilární svěrač prostřednictvím řasnatého ganglia.[4][18]
Hladké pronásledování
Pretektální jádra, zejména NOT, se podílejí na koordinaci pohybů očí během plynulého pronásledování. Tyto pohyby umožňují oku pozorně sledovat pohybující se objekt a dohnat objekt po neočekávané změně směru nebo rychlosti. Směrově citlivé neurony sítnicového skluzu v rámci NOT neposkytují ipsiversivní horizontální informace o chybě sítnice do kůry skrz nižší oliva. Během dne jsou tyto informace snímány a předávány neurony s velkými receptivními poli, zatímco parafoveální neurony s malými receptivními poli to dělají ve tmě. Touto cestou je NOT schopen poskytnout informace o chybě sítnice k vedení pohybů očí.[1][17][19] Kromě své role v udržování plynulého pronásledování se pretektum aktivuje během optokinetického nystagmu, při kterém se oko vrací do centrální polohy směřující dopředu poté, co předmět, který sledoval, projde mimo zorné pole.[20]
Akomodační reflex
Část pretectum, zejména NOT a NPP, jsou zapleteny do akomodační reflex kterým oko udržuje zaostření.[21] Proprioceptivní informace ze sítnice se dostanou do pretektu přes okculomotorický nerv a trigeminální nerv. Od tohoto bodu je mechanismus, kterým oko udržuje zaostření prostřednictvím svalových kontrakcí sítnice, podobný mechanismu pupilárního světelného reflexu.[4]
Antinocicepce
NPA se podílí na aktivním snižování vnímání bolestivých podnětů (antinocicepce).[7] Ačkoli mechanismus, kterým NPA mění reakci organismu na bolestivé podněty, není zcela znám, výzkum ukázal, že aktivita ve ventrální NPA spouští cholinergní a serotonergní neurony. Tyto neurony aktivují sestupné dráhy, které se synchronizují v míše a inhibují nociceptivní buňky v hřbetní roh.[22] Kromě svého přímého antionociceptivního mechanismu se NPA promítá do oblastí mozku, které prostřednictvím připojení k somatosenzorické kůře regulují vnímání bolestivých podnětů. Dva z těchto regionů, o nichž je známo, že NPA promítají, jsou zona incerta a zadní thalamické jádro. Regiony NPA mohou být specializované na reakci na různé typy bolesti. Výzkum zjistil, že dorzální NPA nejlépe snížilo vnímání krátké bolesti, zatímco ventrální NPA snížilo vnímání chronické bolesti.[23] Vzhledem ke své roli při snižování chronické bolesti se předpokládá, že se na ní podílí abnormální aktivita NPA syndrom centrální bolesti.[24]
REM spánek
Na regulaci REM spánku a spánkového chování se může podílet více pretektálních jader. Výzkum ukázal, že pretectum ve spojení s superior colliculus může být zodpovědné za způsobení necikadiánních změn v chování REM spánku.[25] Ukázalo se, že pretektální jádra přijímající retinální vstup, zejména NOT a NPP, jsou částečně zodpovědná za zahájení spánku REM u albínských krys.[5] Objev projekcí z pretekta do několika thalamových jader zapojených do kortikální aktivace během spánku REM, konkrétně projekce do superchiasmatického jádra, které je součástí známého regulačního mechanismu spánku REM, tuto hypotézu podporuje.[12]
Viz také
Reference
- ^ A b C d E F G Gamlin PD (2006). Preekt: spojení a role související s okulomotorem. Pokrok ve výzkumu mozku. 151. 379–405. doi:10.1016 / S0079-6123 (05) 51012-4. ISBN 9780444516961. PMID 16221595.
- ^ A b Magoun HW, Ranson SW (květen 1935). "Centrální dráha světelného reflexu: studie účinku lézí". Archiv oftalmologie. 13 (5): 791–811. doi:10.1001 / archopht.1935,00840050069006.
- ^ Neuhuber W, Schrödl F (listopad 2011). "Autonomní ovládání oka a duhovky". Autonomní neurovědy. 165 (1): 67–79. doi:10.1016 / j.autneu.2010.10.004. PMID 21071284.
- ^ A b C Donkelaar H (2012). Klinické neuroanatomické mozkové obvody a jejich poruchy (1. vyd.). Berlín: Springer. p. 343. ISBN 978-3642191336.
- ^ A b Miller AM, Miller RB, Obermeyer WH, Behan M, Benca RM (srpen 1999). „Prefektum zprostředkovává rychlou regulaci spánku spánkovou regulací světlem“. Behaviorální neurovědy. 113 (4): 755–65. doi:10.1037/0735-7044.113.4.755. PMID 10495083.
- ^ Bosman LW, Houweling AR, Owens CB, Tanke N, Shevchouk OT, Rahmati N, Teunissen WH, Ju C, Gong W, Koekkoek SK, De Zeeuw CI (1. ledna 2011). „Anatomické dráhy zapojené do generování a snímání rytmických pohybů vousů“. Frontiers in Integrative Neuroscience. 5: 53. doi:10.3389 / fnint.2011.00053. PMC 3207327. PMID 22065951.
- ^ A b Reis GM, Rossaneis AC, Silveira JW, Prado WA (červen 2012). „Mechanismy závislé na μ1 a 5-HT1 v předním pretektálním jádru zprostředkovávají antinociceptivní účinky stimulace retrospleniální kůry u potkanů“. Humanitní vědy. 90 (23–24): 950–5. doi:10.1016 / j.lfs.2012.04.023. PMID 22575824.
- ^ Millodot M (2009). Slovník optometrie a vizuální vědy (7. vydání). Edinburgh: Elsevier / Butterworth-Heinemann. ISBN 978-0-7020-2958-5.
- ^ Ramachandran VS (2002). Encyklopedie lidského mozku. Amsterdam: Acad. Lis. ISBN 978-0122272103.
- ^ A b C d E Nieuwenhuys R, ten Donkelaar HJ, Nicholson C (1998). Centrální nervový systém obratlovců. Berlin [u.a.]: Springer. 1812–1817. ISBN 978-3540560135.
- ^ Borostyánkoi-Baldauf Z, Herczeg L (1. března 2002). „Parcelace lidského pretektálního komplexu: chemoarchitektonické přehodnocení“. Neurovědy. 110 (3): 527–40. doi:10.1016 / S0306-4522 (01) 00462-6. PMID 11906791.
- ^ A b C Prichard JR, Stoffel RT, Quimby DL, Obermeyer WH, Benca RM, Behan M (1. října 2002). "Fos imunoreaktivita v subkortikálním vizuálním plášti krysy v reakci na změny osvětlení". Neurovědy. 114 (3): 781–93. doi:10.1016 / S0306-4522 (02) 00293-2. PMID 12220578.
- ^ Morin LP, Blanchard JH (červenec – srpen 1997). "Neuropeptid Y a enkefalin imunoreaktivita v retinorecipientních jádrech křečka pretectum a thalamu". Vizuální neurověda. 14 (4): 765–77. doi:10.1017 / s0952523800012712. PMID 9279004.
- ^ A b Hutchins B, Weber JT (únor 1985). „Pretektální komplex opice: opětovné prozkoumání morfologie a zakončení sítnice“. The Journal of Comparative Neurology. 232 (4): 425–42. doi:10,1002 / k.902320402. PMID 3980762.
- ^ Hutchins B (1991). „Důkazy pro přímou projekci sítnice do předního pretekálního jádra u kočky“. Výzkum mozku. 561: 169–173. doi:10.1016 / 0006-8993 (91) 90764-m.
- ^ Weber JT, Hutchins B (1982). „Demonstrace projekce sítnice do mediálního pretektálního jádra u kočky domácí a opice veverky: anautoradiografická analýza“. Výzkum mozku. 232: 181–186. doi:10.1016/0006-8993(82)90622-9.
- ^ A b Ono S, Mustari MJ (květen 2010). „Signály vizuální chyby z pretektálního jádra optického traktu pro vedení motorického učení pro plynulé pronásledování“. Journal of Neurophysiology. 103 (5): 2889–99. doi:10.1152 / jn.01024.2009. PMC 2867559. PMID 20457849.
- ^ Gamlin PD, Zhang H, Clarke RJ (1995). „Světelné neurony v pretektálním olivárním jádru zprostředkovávají pupilární světelný reflex u opice rhesus“. Experimentální výzkum mozku. 106 (1): 169–76. doi:10.1007 / bf00241367. PMID 8542972.
- ^ Collewijn H (leden 1975). "Okulomotorické oblasti ve středním mozku a pretektu králíků". Journal of Neurobiology. 6 (1): 3–22. doi:10,1002 / neu.480060106. PMID 1185174.
- ^ Dieterich M, Schlindwein P, Janusch B, Bauermann T, Stoeter P, Bense S (1. prosince 2007). "Aktivace mozkového kmene a mozečku během optokinetické stimulace". Klinická neurofyziologie. 118 (12): 2811–2812. doi:10.1016 / j.clinph.2007.09.019.
- ^ Konno S, Ohtsuka K (leden – únor 1997). "Plochy ubytování a zúžení pupilů v kočičím středním mozku". Japonský oftalmologický žurnál. 41 (1): 43–8. doi:10.1016 / s0021-5155 (96) 00010-x. PMID 9147188.
- ^ Villarreal CF, Del Bel EA, Prado WA (květen 2003). „Zapojení předního pretektálního jádra do kontroly přetrvávající bolesti: studie chování a exprese c-Fos u krysy“. Bolest. 103 (1–2): 163–74. doi:10.1016 / S0304-3959 (02) 00449-9. PMID 12749971.
- ^ Villarreal CF, Kina VA, Prado WA (září 2004). "Antinocicepce indukovaná stimulací předního pretektálního jádra u dvou modelů bolesti u krys". Klinická a experimentální farmakologie a fyziologie. 31 (9): 608–13. doi:10.1111 / j.1440-1681.2004.04057.x. PMID 15479168.
- ^ Murray PD, Masri R, Keller A (červen 2010). „Abnormální aktivita předního pretektálního jádra přispívá k syndromu centrální bolesti“. Journal of Neurophysiology. 103 (6): 3044–53. doi:10.1152 / jn.01070.2009. PMC 2888237. PMID 20357063.
- ^ Miller AM, Obermeyer WH, Behan M, Benca RM (červenec 1998). „Superior colliculus-pretectum zprostředkovává přímé účinky světla na spánek“. Sborník Národní akademie věd Spojených států amerických. 95 (15): 8957–62. Bibcode:1998PNAS ... 95,8957M. doi:10.1073 / pnas.95.15.8957. PMC 21184. PMID 9671786.
externí odkazy
- Obarvené obrázky mozkových řezů, které obsahují „Pretectum“ na Projekt BrainMaps
- Dílčí atlas pretectum
| Kontrolní úřad |
|---|