Sonic ježek - Sonic hedgehog
tento článek chybí informace o vývoji číslic končetin, polydaktylie. (Červen 2016) |




Sonic ježek je protein, který je u lidí kódován SHH gen.[5] Je to nejlépe studované ligand z ježková signální dráha; ostatní jsou pouštní ježek (DHH) a Indický ježek (IHH). Hraje klíčovou roli v rozvoj zvířat od hmyzu po savce. v obratlovců, podílí se na organogeneze, včetně růstu číslice na končetinách a organizaci mozku. Sonic hedgehog je archetypálním příkladem a morfogen jak je definováno Lewis Wolpert je Model francouzské vlajky - tj. Molekula, která šíří vytvořit a koncentrační gradient a má různé účinky na vyvíjející se buňky embryo, v závislosti na jeho koncentraci. Sonic ježek je také aktivní u dospělých; například ovládá proliferace z dospělé kmenové buňky a byl zapleten do vývoj některých druhů rakoviny.
Objev a jméno
Gen ježka (hh ) byl poprvé identifikován u ovocné mušky Drosophila melanogaster na klasických obrazovkách Heidelbergu Christiane Nüsslein-Volhard a Eric Wieschaus, jak byla zveřejněna v roce 1980.[6] Tyto obrazovky - což vedlo k tomu, že vyhráli Nobelova cena v roce 1995 spolu s vývojovým genetikem Edward B. Lewis - identifikované geny, které řídí segmentace vzor Drosophila embrya. The hh ztráta funkce mutant fenotyp způsobí, že embrya budou pokryta denticly - tj. malými špičatými výstupky připomínajícími hroty a ježek. Vyšetřování zaměřená na zjištění a ježek ekvivalent u obratlovců do Philip Ingham, Andrew P. McMahon a Clifford Tabin odhalil tři homologní geny.[7][8][9][10]
Dva z těchto genů - tj. pouštní ježek a Indický ježek - byly pojmenovány pro druhy ježků, zatímco zvukový ježek byl pojmenován po Sonic the Hedgehog, protagonista postavy stejnojmenná série videoher.[11][12] Gen byl jako takový pojmenován Robertem Riddleem, který byl postdoktorandem v USA Tabin Lab, poté, co se jeho manželka Betsy Wilder vrátila domů s herním časopisem obsahujícím reklamu na videohru Sonic the Hedgehog.[13][14][15] V zebrafish, dva ze tří obratlovců hh geny jsou duplikovány: SHH a [16] a SHH b[17] (dříve popsáno jako ježek tiggywinkle, pojmenovaný pro Paní Tiggy-Winkle, postava z Beatrix Potterová knihy pro děti) a ihha a ihhb[18] (dříve popsáno jako echidna ježek, pojmenovaný pro ostnatý mravenečník a ne pro postavu Klouby Echidna v Zvukový franšíza).
Funkce
Z hh homologové, SHH bylo zjištěno, že má nejkritičtější role ve vývoji a jedná jako morfogen podílí se na vytváření vzorů mnoha systémů - včetně přední hypofýza[19], pallium mozku[20], mícha[21], plíce[22], zuby[23] a thalamus podle zona limitans intrathalamica.[24][25] U obratlovců rozvoj z končetiny a číslice závisí na sekreci sonického ježka zóna polarizační aktivity, který se nachází na zadní straně embrya končetinový pupen.[9] Mutace v lidském genu sonického ježka SHH způsobit holoprosencephaly typ 3 HPE3 v důsledku ztráty břišní střední čára. Cesta transkripce sonického ježka byla také spojena s tvorbou specifických druhů rakovinných nádorů, včetně embryonálních mozeček nádor[26] a meduloblastom[27], stejně jako postup rakovina prostaty nádory.[28] Aby byl SHH exprimován ve vyvíjejících se končetinách embrya, volal se morfogen růstové faktory fibroblastů musí být vylučován z vrcholový ektodermální hřeben.[29]
Bylo také prokázáno, že Sonic ježek působí jako axonální vodicí tágo. Bylo prokázáno, že SHH přitahuje komisurální axony na ventrální středové čáře vyvíjející se míchy.[30] Konkrétně SHH přitahuje gangliová buňka sítnice (RGC) axony při nízkých koncentracích a odpuzuje je při vyšších koncentracích.[31] Bylo prokázáno, že absence (neexprese) SHH reguluje růst rodících se zadních končetin kytovci[32] (velryby a delfíni ).
The SHH Gen je členem rodiny genových ježků s pěti variantami změn sekvence DNA nebo variant sestřihu.[33] SHH se nachází na sedmém chromozomu a iniciuje produkci proteinu Sonic Hedgehog.[33] Tento protein vysílá signály krátkého a dlouhého dosahu do embryonálních tkání, aby reguloval vývoj.[34] Pokud SHH Gen je mutovaný nebo chybí, protein Sonic Hedgehog nemůže dělat svou práci správně. Sonic hedgehog přispívá k buněčnému růstu, specifikaci a formování buněk, strukturování a organizaci tělesného plánu.[35] Tento protein funguje jako důležitá morfogenní signální molekula a hraje důležitou roli při tvorbě mnoha různých struktur ve vývoji embryí.[35] The SHH Gen ovlivňuje několik hlavních orgánových systémů, jako je nervový systém, kardiovaskulární systém, dýchací systém a pohybový aparát.[33][34] Mutace v SHH Gen může způsobit malformaci složek těchto systémů, což může mít za následek velké problémy ve vývoji embrya. Například mozek a oči mohou být významně ovlivněny mutacemi v tomto genu a způsobit poruchy, jako je Mikroftalmie a Holoprosencefalie.[35] Mikroftalmie je stav, který postihuje oči, což vede k malým nerozvinutým tkáním v jednom nebo obou očích.[35] To může vést k problémům od colobomu po jedno malé oko až po úplnou absenci očí.[34] Holoprosencephaly je stav nejčastěji způsobený mutací SHH gen, který způsobuje nesprávné oddělení levého a pravého mozku a dysmorfie obličeje.[34][35] Mnoho systémů a struktur se silně spoléhá na správné vyjádření SHH gen a následný protein sonic hedgehog, což mu dává vyznamenání za základní gen pro vývoj.
Vzorec centrální nervové soustavy
Signální molekula sonického ježka (SHH) zaujímá při vzorování různé role centrální nervový systém (CNS) během obratlovců rozvoj. Jednou z nejcharakterističtějších funkcí SHH je jeho role při indukci podlahovou desku a různé typy ventrálních buněk v rámci neurální trubice.[36] The notochord - struktura odvozená od osy mezoderm - produkuje SHH, který extracelulárně cestuje do ventrální oblasti neurální trubice a dává pokyn těmto buňkám, aby vytvořily podlahovou desku.[37] Jiný pohled na indukci podlahové desky předpokládá, že některé prekurzorové buňky umístěné v notochordu jsou vloženy do neurální desky před jejím vytvořením, což později vede k podlahové desce.[38]
The neurální trubice sama o sobě je počátečním základem CNS obratlovců a podlahovou desku je specializovaná struktura, umístěná ve ventrálním středu neurální trubice. Důkazy podporující notochord jako signální centrum pocházejí ze studií, ve kterých je druhý neurochord implantován poblíž neurální trubice in vivo, což vede k vytvoření ektopické podlahové desky v neurální trubici.[39]

Gradienty SHH a BMP v neurální trubici obratlovců

Vytvoření ektopické podlahové desky

Ventrální neurální domény v neurální trubici



Sonic ježek je vylučovaný protein který zprostředkovává signalizační aktivity notochordu a podlahové desky.[40] Studie zahrnující ektopický výraz SHH in vitro[41] a in vivo[42] vést k indukci podlahové desky a diferenciace z motorický neuron a ventrální interneurony. Na druhé straně myší mutanty pro SHH postrádají charakteristiky míchy.[43]In vitro blokování signalizace SHH pomocí protilátky proti tomu vykazuje podobné fenotypy.[42] SHH uplatňuje své účinky způsobem závislým na koncentraci[44], takže vysoká koncentrace SHH má za následek místní inhibice z buněčná proliferace.[45] Tato inhibice způsobí ztenčení podlahové desky ve srovnání s bočními oblastmi neurální trubice. Nižší koncentrace SHH vede k buněčné proliferaci a indukci různých typů ventrálních nervových buněk.[42] Jednou podlahovou desku je stanoveno, buňky pobývající v této oblasti budou následně exprimovat SHH samy[45], generování a koncentrační gradient uvnitř neurální trubice.
Ačkoli neexistují žádné přímé důkazy o SHH spád, existuje nepřímý důkaz prostřednictvím vizualizace Patched (Ptc) genová exprese, která kóduje ligand vazebná doména SHH receptor[46] skrz ventrální neurální trubici.[47] In vitro studie ukazují, že přírůstkové dvoj- a trojnásobné změny v koncentraci SHH vedou ke vzniku motorických neuronů a různých interneuronálních podtypů, které se nacházejí ve ventrální míše.[48] Tyto přírůstkové změny in vitro odpovídají vzdálenosti domén ze signální tkáně (notochord a podlahová deska), která se následně diferencuje na různé neuronální podtypy, jak se vyskytuje in vitro.[49] Navrhuje se, aby odstupňovaná signalizace SHH byla zprostředkována prostřednictvím Gli rodina proteinů, které jsou homology obratlovců Drosophila zinkový prst -obsahující transkripční faktor Cubitus interruptus (Ci). Ci je rozhodujícím prostředníkem ježka (Hh) signalizace Drosophila.[50] U obratlovců jsou přítomny tři různé proteiny Gli, viz. Gli1, Gli2 a Gli3, které jsou vyjádřeny v neurální trubici.[51] Myší mutanti pro Gli1 vykazují normální vývoj míchy, což naznačuje, že je postradatelný pro zprostředkování aktivity SHH.[52] Gli2 mutantní myši však vykazují abnormality ve ventrální míše, s vážnými defekty na podlahové desce a nejvíce ventrálních interneuronech (V3).[53] Gli3 antagonizuje Funkce SHH v a dávka -závislým způsobem, podporujícím dorzální neuronální podtypy. Mutantní fenotypy SHH lze zachránit v SHH / Gli3 double mutant.[54] Gli proteiny mají C-koncovou aktivační doménu a N-koncovou represivní doménu.[51][55]
SHH se navrhuje k podpoře aktivační funkce Gli2 a inhibici represivní aktivity Gli3. Zdá se, že SHH také podporuje aktivační funkci Gli3, ale tato aktivita není dostatečně silná.[54] Stupněná koncentrace SHH vede ke stupňované aktivitě Gli 2 a Gli3, které podporují ventrální a dorzální neuronální podtypy ve ventrální míše. Důkazy od mutantů Gli3 a SHH / Gli3 ukazují, že SHH primárně reguluje prostorové omezení předek spíše než induktivní domény, protože mutanty SHH / Gli3 ukazují vzájemné míchání buněčných typů.[54][56]
SHH také indukuje další proteiny, se kterými interaguje, a tyto interakce mohou ovlivnit citlivost buňky vůči SHH. Protein interagující s ježky (HHIP ) je indukován SHH, což zase tlumí jeho signalizační aktivitu.[57] Vitronektin je další protein, který je indukován SHH; působí jako povinný faktor pro signalizaci SHH v neurální trubici.[58]
Ve ventrální neurální trubici je pět odlišných domén progenitorů: V3 interneurony, motorické neurony (MN), V2, V1 a V0 interneurony (ve ventrálním až dorzálním pořadí).[48] Tyto různé progenitorové domény jsou založeny „komunikací“ mezi různými třídami homeobox transkripční faktory. (Vidět Trigeminální nerv.) Tyto transkripční faktory reagovat na koncentraci gradientu SHH. V závislosti na povaze jejich interakce s SHH jsou klasifikovány do dvou skupin - třída I a třída II - a jsou složeny z členů z Pax, Nkx, Dbx a Irx rodiny.[45] Proteiny třídy I jsou potlačovány na různých prahových hodnotách SHH vymezujících ventrální hranice progenitorové domény, zatímco proteiny třídy II jsou aktivovány na různých prahových hodnotách SHH vymezujících dorzální limit domén. Selektivní křížrepresivní interakce mezi proteiny třídy I a třídy II vedou k pěti kardinálním ventrálním neuronálním podtypům.[59]
Je důležité si uvědomit, že SHH není jediný signální molekula působící na vyvíjející se neurální trubici. Mnoho dalších molekul, cesty a mechanismy jsou aktivní (např. RA, FGF, BMP ) a jsou možné komplexní interakce mezi SHH a jinými molekulami. Předpokládá se, že BMP hrají rozhodující roli při určování citlivosti nervových buněk na signalizaci SHH. Důkazy podporující toto pocházejí ze studií používajících inhibitory BMP, které ventralizují osud buňky nervové desky pro danou koncentraci SHH.[60] Na druhou stranu mutace v antagonistech BMP (např. člun ze dřeva ) produkuje závažné defekty ve ventrálních charakteristikách míchy, po nichž následuje ektopický výraz BMP ve ventrální neurální trubici.[61] Interakce SHH s Fgf a RA ještě nebyly molekulárně podrobně studovány.
Morfogenetická aktivita
Koncentrační a časově závislá aktivita SHH určující osud buněk v břišní neurální trubice je vynikajícím příkladem a morfogen. U obratlovců je signalizace SHH ve ventrální části neurální trubice nejvýznamnější odpovědností za indukci podlahovou desku buňky a motorické neurony.[62] SHH vyzařuje z notochord a ventrální podlahovou desku vyvíjející se neurální trubice k vytvoření a koncentrační gradient který se klene nad osou dorso-ventrální a je antagonizován inverzí Wnt gradient, který určuje hřbetní míchu.[63][64] Vyšší koncentrace ligandu SHH se nacházejí v nejvíce ventrálních aspektech neurální trubice a notochordu, zatímco nižší koncentrace se nacházejí ve více dorzálních oblastech neurální trubice.[63] Gradient koncentrace SHH byl vizualizován v neurální trubici myší konstruovaných tak, aby exprimoval fúzní protein SHH :: GFP, aby se ukázalo toto odstupňované rozdělení SHH v době vzorování ventrální neurální trubice.[65]
Předpokládá se, že gradient SHH pracuje na vyvolání více různých buněčných osudů mechanismem závislým na koncentraci a čase, který indukuje řadu transkripčních faktorů ve ventrální oblasti progenitorové buňky.[63][65] Každá z ventrálních progenitorových domén exprimuje vysoce individualizovanou kombinaci transkripčních faktorů - Nkx2.2, Olig2, Nkx6.1, Nkx6.2, Dbx1, Dbx2, Irx3, Pax6 a Pax7 - která je regulována gradientem SHH. Tyto transkripční faktory jsou indukovány postupně podél koncentračního gradientu SHH s ohledem na množství a dobu expozice SHH ligandu.[63] Protože každá populace progenitorových buněk reaguje na různé hladiny proteinu SHH, začínají exprimovat jedinečnou kombinaci transkripčních faktorů, která vede k diferenciaci osudu neuronových buněk. Tato diferenciální genová exprese indukovaná SHH vytváří ostré hranice mezi diskrétními domén exprese transkripčního faktoru, který nakonec formuje ventrální neurální trubici.[63]
Prostorový a časový aspekt progresivní indukce genů a buněčných osudů ve ventrální neurální trubici je ilustrován expresními doménami dvou z nejlépe charakterizovaných transkripčních faktorů Olig2 a Nkx2.2.[63] Na počátku vývoje byly buňky na ventrální středové čáře vystaveny nízké koncentraci SHH pouze po relativně krátkou dobu a exprimují transkripční faktor Olig2.[63] Exprese Olig2 se rychle rozšiřuje v dorzálním směru současně s kontinuálním dorzálním prodloužením gradientu SHH v čase.[63] Jak se však morfogenetická fronta SHH ligandu pohybuje a začíná růst více koncentrovaná, buňky, které jsou vystaveny vyšším hladinám ligandu, reagují vypnutím Olig2 a zapnutím Nkx2.2[63], vytvoření ostré hranice mezi buňkami exprimujícími transkripční faktor Nkx2.2 ventrálně s buňkami exprimujícími Olig2. To je tímto způsobem, že každá z domén šesti populací progenitorových buněk je považována za postupně vzorovanou v celé neurální trubici gradientem koncentrace SHH.[63] Vzájemná inhibice mezi páry transkripčních faktorů exprimovaných v sousedních doménách přispívá k rozvoji ostrých hranic; v některých případech však byl nalezen inhibiční vztah i mezi dvojicemi transkripčních faktorů ze vzdálenějších domén. Zejména, NKX2-2 se uvádí, že exprimovaný v doméně V3 inhibuje IRX3 exprimován ve V2 a více hřbetních doménách, ačkoli V3 a V2 jsou odděleny další doménou nazvanou MN.[66]
Vývoj zubů
Sonic hedgehog (SHH) je signální molekula, která je kódována stejným genem jako sonic hedgehog. SHH hraje velmi důležitou roli v organogenezi a především v kraniofaciálním vývoji. Jelikož SHH je signální molekula, funguje primárně difúzí podél koncentračního gradientu a ovlivňuje buňky různými způsoby. V časném vývoji zubu se SHH uvolňuje z primárního smaltovaný uzel - signální centrum - poskytovat informace o poloze jak v laterálním, tak v rovinném signálním vzoru ve vývoji zubů a regulaci růstu zubních hrotů.[67] Zejména SHH je potřebný pro růst epiteliálních krčních smyček, kde se vnější a vnitřní epitel spojují a tvoří rezervoár pro zubní kmenové buňky. Po apoptování primárních smaltových uzlů se vytvoří sekundární smaltové uzly. Sekundární uzly skloviny vylučují SHH v kombinaci s dalšími signálními molekulami, aby zesílily orální ektodermu a začaly modelovat složité tvary korunky zubu během diferenciace a mineralizace.[68] V modelu knockoutového genu je absence SHH indikativní pro holoprosencephaly. SHH však aktivuje následné molekuly Gli2 a Gli3. Embrya mutovaných Gli2 a Gli3 mají abnormální vývoj řezáků, které jsou zastaveny v raném vývoji zubu, stejně jako malé stoličky.[69]
Vývoj plic
Ačkoli SHH je nejčastěji spojován s vývojem číslic mozku a končetin, je také důležitý ve vývoji plic.[70][71][72][73] Studie využívající qPCR a knockouty ukázaly, že SHH přispívá k embryonálnímu vývoji plic. Rozvětvení savčích plic se vyskytuje v epitelu vyvíjejícího se průdušky a plíce.[74][75] SHH vyjádřený v celém předpažbí endoderm (nejvnitřnější ze tří zárodečných vrstev) v distálním epitelu, kde se vyvíjejí embryonální plíce.[72][75] To naznačuje, že SHH je částečně zodpovědný za větvení plic. Další důkazy role SHH při větvení plic byly pozorovány u qPCR. Exprese SHH se vyskytuje ve vyvíjejících se plicích kolem embryonálního dne 11 a je silně vyjádřena v pupenech plodových plic, ale nízko ve vyvíjejících se průduškách.[72][75] U myší s nedostatkem SHH se může vyvinout tracheoezofageální píštěl (abnormální spojení jícnu a průdušnice).[76][72] Navíc dvojitý (SHH - / -) knockoutový myší model vykazoval špatný vývoj plic. Plíce dvojitého knockoutu SHH neprošly lobací a rozvětvením (tj. Abnormální plíce vyvinuly pouze jednu větev, ve srovnání s extenzivně rozvětveným fenotypem divokého typu).[72]
Potenciální regenerační funkce
Sonic ježek může hrát roli u savců vlásková buňka regenerace. Modulací retinoblastomový protein aktivita u krysího slimáka, sonický ježek umožňuje dospět vláskové buňky které se za normálních okolností nemohou vrátit do proliferačního stavu, aby se rozdělily a rozlišily. Proteiny retinoblastomu potlačit růst buněk tím, že brání buňkám v návratu do buněčný cyklus, čímž zabrání šíření. Zdá se, že inhibice aktivity Rb umožňuje buňkám dělení. Proto se sonický ježek - označený jako důležitý regulátor Rb - může také ukázat jako důležitý rys při opětovném růstu vlasových buněk po poškození.[77]
zpracovává se
SHH prochází řadou zpracovatelských kroků, než je vylučován z buňky. Nově syntetizovaný SHH váží 45kDa a označuje se jako preproprotein. Jako vylučovaný protein obsahuje krátký sekvence signálu na svém N-konci, který je rozpoznán částice pro rozpoznávání signálu během přemístění do endoplazmatické retikulum (ER), první krok v bílkovinách vylučování. Jakmile je translokace dokončena, signální sekvence je odstraněna pomocí signální peptidáza v pohotovostním režimu. Zde SHH prochází automatickým zpracováním za účelem generování 20-kDa N-terminální signální domény (SHH-N) a 25 kDa C-terminální domény bez známé signální role.[78] Štěpení je katalyzováno a proteáza v C-terminální doméně. Během reakce a cholesterol molekula je přidána na C-konec SHH-N.[79][80] C-koncová doména tedy funguje jako intein a cholesterol transferáza. Další hydrofobní skupina, a palmitát, se přidá k alfa-aminu na N-konci cystein SHH-N. Tato modifikace je nutná pro efektivní signalizaci, která má za následek třicetinásobné zvýšení účinnosti nad nepalitylovanou formou a provádí ji člen membránově vázaná O-acyltransferáza rodina Protein-cystein-N-palmitoyltransferáza HHAT.[81]
Robotnikinin
Byl nalezen potenciální inhibitor signální dráhy ježka a byl nazván „Robotnikinin“ - na počest nemesis Sonic the Hedgehog, Dr. Ivo "Eggman" Robotnik[82].
Kontroverze kolem jména
Gen byl spojen se stavem známým jako holoprosencephaly, což může mít za následek závažné poškození mozku, lebky a obličeje, což způsobí, že lékaři a vědci toto jméno kritizují z toho důvodu, že zní příliš frivolně. Bylo poznamenáno, že zmínka o mutaci v a zvukový ježek gen nemusí být dobře přijat v diskusi o vážné poruše s pacientem nebo jeho rodinou.[13][83][84]
Galerie
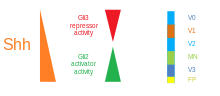 SHH gradient a aktivita Gli v neurální trubici obratlovců. |  Zpracování SHH | 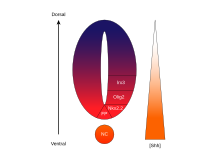 |
Viz také
- Pikachurin retinální protein pojmenovaný po Pikachu
- Zbtb7 onkogen, který byl původně pojmenován „Pokémon "
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000164690 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000002633 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Marigo V, Roberts DJ, Lee SM, Tsukurov O, Levi T, Gastier JM, Epstein DJ, Gilbert DJ, Copeland NG, Seidman CE (červenec 1995). „Klonování, exprese a chromozomální umístění SHH a IHH: dva lidské homology hedgehogu genu pro polaritu segmentu Drosophila“. Genomika. 28 (1): 44–51. doi:10.1006 / geno.1995.1104. PMID 7590746.
- ^ Nüsslein-Volhard C, Wieschaus E (říjen 1980). Msgstr "Mutace ovlivňující počet segmentů a polaritu v Drosophile". Příroda. 287 (5785): 795–801. Bibcode:1980Natur.287..795N. doi:10.1038 / 287795a0. PMID 6776413. S2CID 4337658.
- ^ Krauss S, Concordet JP, Ingham PW (prosinec 1993). „Funkčně konzervovaný homolog genu hh polarity segmentu Drosophila je exprimován v tkáních s polarizační aktivitou v embryích zebrafish“. Buňka. 75 (7): 1431–44. doi:10.1016/0092-8674(93)90628-4. PMID 8269519. S2CID 46266627.
- ^ Echelard Y, Epstein DJ, St-Jacques B, Shen L, Mohler J, McMahon JA, McMahon AP (prosinec 1993). „Sonic ježek, člen rodiny domnělých signálních molekul, je zapojen do regulace polarity CNS“. Buňka. 75 (7): 1417–30. doi:10.1016/0092-8674(93)90627-3. PMID 7916661. S2CID 6732599.
- ^ A b Riddle RD, Johnson RL, Laufer E, Tabin C (1993). „Sonic ježek zprostředkovává polarizační aktivitu ZPA.“ Buňka. 75 (7): 1401–16. doi:10.1016/0092-8674(93)90626-2. PMID 8269518. S2CID 4973500.
- ^ Angier N (11.01.1994). „Biologové najdou klíčové geny, které formují vzor embryí“. Věda. New York Times.
- ^ Anwood R. (2007-09-06). Emus nemůže jít zpět. Ebury Press. str. 113–114. ISBN 978-0-09-192151-4.
- ^ Tom Simonite (2005-12-15). „Pokémon blokuje genové jméno“. 438 (897). Příroda. doi:10.1038 / 438897a. Citováno 2013-05-23.
- ^ A b „Gene jménem Sonic“. The New York Times. 1994-01-11.
- ^ Keen A, Tabin C (12. dubna 2004). „Cliff Tabin: Super Sonic An Interview“. Týdenní šelest. Archivovány od originál 10. listopadu 2005. Citováno 24. dubna 2014.
- ^ Interview s Robertem Riddleem, o pojmenování genu, obsahuje „Ingenious: The Cyclops Gene“, funkci BBC Radio od Kat Arney https://www.bbc.co.uk/sounds/play/m000h263
- ^ „Zebrafish SHHa“. University of Oregon. Archivovány od originál dne 25. 06. 2009.
- ^ „Zebrafish SHHb“. University of Oregon. Archivovány od originál dne 26.06.2009.
- ^ Currie PD, Ingham PW (srpen 1996). „Indukce konkrétního typu svalové buňky proteinem podobným ježkovi v zebrafish“. Příroda. 382 (6590): 452–5. Bibcode:1996 Natur.382..452C. doi:10.1038 / 382452a0. PMID 8684485. S2CID 4271898.
- ^ Herzog W, Zeng X, Lele Z, Sonntag C, Ting JW, Chang CY, Hammerschmidt M (únor 2003). "Tvorba adenohypofýzy u zebrafish a její závislost na sonic ježka". Dev. Biol. 254 (1): 36–49. doi:10.1016 / S0012-1606 (02) 00124-0. PMID 12606280.
- ^ Rash BG, Grove EA (říjen 2007). „Vzory hřbetního telencephalonu: role sonického ježka?“. The Journal of Neuroscience. 27 (43): 11595–603. doi:10.1523 / JNEUROSCI.3204-07.2007. PMC 6673221. PMID 17959802.
- ^ Lewis KE, Eisen JS (září 2001). "Ježková signalizace je nutná pro primární indukci motoneuronu u zebrafish". Rozvoj. 128 (18): 3485–95. PMID 11566854.
- ^ Wolpert L (2015). Zásady rozvoje (5. vydání). Oxford University Press. str. 500.
- ^ Dassule HR, Lewis P, Bei M, Maas R, McMahon AP (listopad 2000). "Sonic ježek reguluje růst a morfogenezi zubu" (PDF). Rozvoj. 127 (22): 4775–85. PMID 11044393.
- ^ Scholpp S, Wolf O, značka M, Lumsden A (březen 2006). "Ježek signalizuje z zona limitans intrathalamica organizuje vzorování zebrafish diencephalon ". Rozvoj. 133 (5): 855–64. doi:10.1242 / dev.02248. PMID 16452095. S2CID 27550686.
- ^ Rash BG, Grove EA (listopad 2011). „Pst a Gli3 regulují tvorbu telencefalicko-diencefalického spojení a potlačují zdroj signálu podobný šíji v předním mozku“. Vývojová biologie. 359 (2): 242–50. doi:10.1016 / j.ydbio.2011.08.026. PMC 3213684. PMID 21925158.
- ^ Taylor MD, Northcott PA, Korshunov A, Remke M, Cho YJ, Clifford SC, Eberhart CG, Parsons DW, Rutkowski S, Gajjar A, Ellison DW, Lichter P, Gilbertson RJ, Pomeroy SL, Kool M, Pfister SM (duben 2012 ). „Molekulární podskupiny meduloblastomu: současná shoda“. Acta Neuropathologica. 123 (4): 465–72. doi:10.1007 / s00401-011-0922-z. PMC 3306779. PMID 22134537.
- ^ DeSouza RM, Jones BR, Lowis SP, Kurian KM (22. července 2014). „Pediatrický meduloblastom - aktuální informace o molekulární klasifikaci při cílené terapii“. Hranice v onkologii. 4: 176. doi:10.3389 / fonc.2014.00176. PMC 4105823. PMID 25101241.
- ^ Lubik AA, Nouri M, Truong S, Ghaffari M, Adomat HH, Corey E, Cox ME, Li N, Guns ES, Yenki P, Pham S, Buttyan R (2016). „Signalizace parakrinního sonického ježka významně přispívá k získané steroidogenezi v mikroprostředí nádoru prostaty“. International Journal of Cancer. 140 (2): 358–369. doi:10.1002 / ijc.30450. PMID 27672740. S2CID 2354209.
- ^ Tabin C, Riddle R (únor 1999). "Jak se končetiny vyvíjejí". Scientific American. 280 (2): 74–9. Bibcode:1999SciAm.280b..74R. doi:10.1038 / scientificamerican0299-74. PMID 9924814.
- ^ Charron F, Stein E, Jeong J, McMahon AP, Tessier-Lavigne M (2003). „Morfogenní sonický ježek je axonální chemoatraktant, který spolupracuje s netrin-1 při vedení axonů ve střední linii“. Buňka. 113 (1): 11–23. doi:10.1016 / S0092-8674 (03) 00199-5. PMID 12679031. S2CID 13909497.
- ^ Kolpak A, Zhang J, Bao ZZ (březen 2005). „Sonic ježek má dvojí účinek na růst axonů ganglií sítnice v závislosti na jeho koncentraci“. J. Neurosci. 25 (13): 3432–41. doi:10.1523 / JNEUROSCI.4938-04.2005. PMC 1564194. PMID 15800198.
- ^ Thewissen J, Cohn MJ, Stevens LS, Bajpai S, Heyning J, Horton WE (květen 2006). „Vývojový základ pro ztrátu zadních končetin u delfínů a původ tělesného plánu kytovců“. Proc. Natl. Acad. Sci. USA. 103 (22): 8414–8. Bibcode:2006PNAS..103.8414T. doi:10.1073 / pnas.0602920103. PMC 1482506. PMID 16717186.
- ^ A b C „ENSG00000164690“. GRCh38. Vydání souboru 99. Citováno 9. dubna 2020.
- ^ A b C d „UniprotKB - Q15465 (SHH_HUMAN)“. UniProt. Konsorcium UniProt. Citováno 9. dubna 2020.
- ^ A b C d E "Gen SHH". Genetická domácí reference. Americká národní lékařská knihovna. Citováno 9. dubna 2020.
- ^ Litingtung Y, Chiang C (říjen 2000). "Řízení aktivity SHH a signalizace v neurální trubici". Dynamika vývoje. 219 (2): 143–54. doi:10.1002 / 1097-0177 (2000) 9999: 9999 <:: AID-DVDY1050> 3.0.CO; 2-Q. PMID 11002335.
- ^ Placzek M (srpen 1995). "Role notochordu a podlahové desky v indukčních interakcích". Aktuální názor na genetiku a vývoj. 5 (4): 499–506. doi:10.1016 / 0959-437X (95) 90055-L. PMID 7580143.
- ^ Teillet MA, Lapointe F, Le Douarin NM (září 1998). „Vztahy mezi notochordem a podlahovou deskou při vývoji obratlovců byly znovu navštíveny“. Sborník Národní akademie věd USA. 95 (20): 11733–8. Bibcode:1998PNAS ... 9511733T. doi:10.1073 / pnas.95.20.11733. PMC 21709. PMID 9751734.
- ^ van Straaten HW, Hekking JW, Thors F, Wiertz-Hoessels EL, Drukker J (říjen 1985). "Indukce další podlahové desky v neurální trubici". Acta Morphol Neerl Scand. 23 (2): 91–7. PMID 3834777.
- ^ Patten I, Placzek M (2000). "Role Sonic ježka ve vzorcích neurální trubice". Buněčné a molekulární biologické vědy. 57 (12): 1695–1708. doi:10.1007 / PL00000652. PMID 11130176. S2CID 20950575.
- ^ Martí E, Bumcrot DA, Takada R, McMahon AP (květen 1995). "Požadavek 19K formy Sonic ježka pro indukci odlišných typů ventrálních buněk v CNS explantátech". Příroda. 375 (6529): 322–325. Bibcode:1995 Natur.375..322M. doi:10.1038 / 375322a0. PMID 7753196. S2CID 4362006.
- ^ A b C Ericson J, Morton S, Kawakami A, Roelink H, Jessell TM (listopad 1996). "Dvě kritická období signalizace Sonic Hedgehog vyžadovaná pro specifikaci identity motorických neuronů". Buňka. 87 (4): 661–73. doi:10.1016 / S0092-8674 (00) 81386-0. PMID 8929535. S2CID 11578260.
- ^ Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA (říjen 1996). "Cyclopia a defektní axiální vzorování u myší postrádajících funkci genu Sonic hedgehog". Příroda. 383 (6599): 407–13. Bibcode:1996 Natur.383..407C. doi:10.1038 / 383407a0. PMID 8837770. S2CID 4339131.
- ^ Placzek M, Tessier-Lavigne M, Yamada T, Jessell T, Dodd J (listopad 1990). "Mezodermální řízení identity neurálních buněk: indukce podlahové desky notochordem". Věda. 250 (4983): 985–8. Bibcode:1990Sci ... 250..985P. doi:10.1126 / science.2237443. PMID 2237443.
- ^ A b C Wilson L, Maden M (červen 2005). "Mechanismy dorzoventrálního vzorování v neurální trubici obratlovců". Dev. Biol. 282 (1): 1–13. doi:10.1016 / j.ydbio.2005.02.027. PMID 15936325.
- ^ Stone DM, Hynes M, Armanini M, Swanson TA, Gu Q, Johnson RL, Scott MP, Pennica D, Goddard A, Phillips H, Noll M, Hooper JE, de Sauvage F, Rosenthal A (listopad 1996). "Patchovaný gen potlačující nádor kóduje kandidátský receptor pro Sonic hedgehog". Příroda. 384 (6605): 129–34. Bibcode:1996 Natur.384..129S. doi:10.1038 / 384129a0. PMID 8906787. S2CID 4342540.
- ^ Marigo V, Tabin CJ (1996). "Regulace záplaty sonickým ježkem ve vyvíjející se neurální trubici". Proc. Natl. Acad. Sci. USA. 93 (18): 9346–51. Bibcode:1996PNAS ... 93,9346M. doi:10.1073 / pnas.93.18.9346. PMC 38430. PMID 8790332.
- ^ A b Ericson J, Briscoe J, Rashbass P, van Heyningen V, Jessell TM (1997). "Gradovaná zvuková signalizace ježků a specifikace buněčného osudu ve ventrální neurální trubici". Cold Spring Harb Symp Quant Biol. 62: 451–66. doi:10.1101 / SQB.1997.062.01.053. PMID 9598380.
- ^ Ericson J, Rashbass P, Schedl A, Brenner-Morton S, Kawakami A, van Heyningen V, Jessell TM, Briscoe J (červenec 1997). "Pax6 řídí identitu progenitorových buněk a osud neuronů v reakci na gradovanou signalizaci SHH". Buňka. 90 (1): 169–80. doi:10.1016 / S0092-8674 (00) 80323-2. PMID 9230312. S2CID 17054686.
- ^ Lum L, Beachy PA (červen 2004). "Síť reakce na ježka: senzory, přepínače a směrovače". Věda. 304 (5678): 1755–9. Bibcode:2004Sci ... 304.1755L. CiteSeerX 10.1.1.476.3902. doi:10.1126 / science.1098020. PMID 15205520. S2CID 13949436.
- ^ A b Ruiz i Altaba A (červen 1998). "Kombinatoriální funkce genu Gli v podlahové desce a neuronové indukce pomocí sonického ježka". Rozvoj. 125 (12): 2203–12. PMID 9584120.
- ^ Park HL, Bai C, Platt KA, Matise MP, Beeghly A, Hui CC, Nakashima M, Joyner AL (duben 2000). „Myší mutanti Gli1 jsou životaschopní, ale mají defekty v signalizaci SHH v kombinaci s mutací Gli2“. Rozvoj. 127 (8): 1593–605. PMID 10725236.
- ^ Matise MP, Epstein DJ, Park HL, Platt KA, Joyner AL (srpen 1998). „Gli2 je vyžadován pro indukci podlahové desky a přilehlých buněk, ale ne většiny ventrálních neuronů v myším centrálním nervovém systému“. Rozvoj. 125 (15): 2759–70. PMID 9655799.
- ^ A b C Litingtung Y, Chiang C (říjen 2000). „Specifikace typů ventrálních neuronů je zprostředkována antagonistickou interakcí mezi SHH a Gli3“. Nat Neurosci. 3 (10): 979–85. doi:10.1038/79916. PMID 11017169. S2CID 10102370.
- ^ Sasaki H, Nishizaki Y, Hui C, Nakafuku M, Kondoh H (září 1999). „Regulace aktivit Gli2 a Gli3 aminoterminální represivní doménou: implikace Gli2 a Gli3 jako primárních mediátorů signalizace SHH“. Rozvoj. 126 (17): 3915–24. PMID 10433919.
- ^ Persson M, Stamataki D, te Welscher P, Andersson E, Böse J, Rüther U, Ericson J, Briscoe J (listopad 2002). „Dorsálně-ventrální vzorování míchy vyžaduje aktivitu transkripčního represoru Gli3“. Genes Dev. 16 (22): 2865–78. doi:10,1101 / gad.243402. PMC 187477. PMID 12435629.
- ^ Chuang PT, McMahon AP (únor 1999). "Signalizace obratlovců ježek modulovaná indukcí proteinu vázajícího ježek". Příroda. 397 (6720): 617–21. Bibcode:1999 Natur.397..617C. doi:10.1038/17611. PMID 10050855. S2CID 204991314.
- ^ Pons S, Martí E (leden 2000). „Sonic hedgehog synergizuje s proteinem extracelulární matrix vitronektin k indukci diferenciace spinálních motorických neuronů“. Rozvoj. 127 (2): 333–42. PMID 10603350.
- ^ Briscoe J, Pierani A, Jessell TM, Ericson J (květen 2000). "Homeodoménový proteinový kód specifikuje progenitorovou buněčnou identitu a neuronální osud ve ventrální neurální trubici". Buňka. 101 (4): 435–45. doi:10.1016 / S0092-8674 (00) 80853-3. PMID 10830170. S2CID 17295764.
- ^ Liem KF, Jessell TM, Briscoe J (listopad 2000). „Regulace aktivity nervových vzorců sonického ježka vylučovanými inhibitory BMP vyjádřenými notochordem a somity“. Rozvoj. 127 (22): 4855–66. PMID 11044400.
- ^ McMahon JA, Takada S, Zimmerman LB, Fan CM, Harland RM, McMahon AP (květen 1998). „Nogginem zprostředkovaný antagonismus signalizace BMP je nutný pro růst a tvorbu vzorů neurální trubice a somitu“. Genes Dev. 12 (10): 1438–52. doi:10.1101 / gad.12.10.1438. PMC 316831. PMID 9585504.
- ^ Roelink H, Porter JA, Chiang C, Tanabe Y, Chang DT, Beachy PA, Jessell TM (květen 1995). „Indikace podlahové desky a motorických neuronů různými koncentracemi aminoterminálního štěpného produktu sonické autoproteolýzy ježka“. Buňka. 81 (3): 445–55. doi:10.1016/0092-8674(95)90397-6. PMID 7736596. S2CID 18937753.
- ^ A b C d E F G h i j Ribes V, Briscoe J (srpen 2009). „Zavedení a interpretace odstupňovaného sonického ježka během modelování neurální trubice obratlovců: role negativní zpětné vazby“. Cold Spring Harb Perspect Biol. 1 (2): a002014. doi:10.1101 / cshperspect.a002014. PMC 2742090. PMID 20066087.
- ^ Muroyama Y, Fujihara M, Ikeya M, Kondoh H, Takada S (březen 2002). „Signalizace Wnt hraje zásadní roli v neuronální specifikaci hřbetní míchy“. Geny a vývoj. 16 (5): 548–53. doi:10,1101 / gad. 937102. PMC 155351. PMID 11877374.
- ^ A b Chamberlain CE, Jeong J, Guo C, Allen BL, McMahon AP (březen 2008). „Shh odvozený od notochordu se koncentruje v těsné asociaci s apikálně umístěným bazálním tělem v nervových cílových buňkách a vytváří dynamický gradient během neurálního vzorování“. Rozvoj. 135 (6): 1097–106. doi:10.1242 / dev.013086. PMID 18272593. S2CID 17431502.
- ^ Lovrics A, Gao Y, Juhász B, Bock I, Byrne HM, Dinnyés A, Kovács KA (listopad 2014). „Booleovské modelování odhaluje nová regulační spojení mezi transkripčními faktory, které zorganizují vývoj ventrální míchy“. PLOS ONE. 9 (11): 11430. Bibcode:2014PLoSO ... 9k1430L. doi:10.1371 / journal.pone.0111430. PMC 4232242. PMID 25398016.
- ^ Nanci A (2012). Orální histologie Ten Cate: vývoj, struktura a funkce (8. vydání). St. Louis, Mo .: Elsevier. ISBN 978-0-323-07846-7.
- ^ Thesleff I (2003). "Epiteliálně-mezenchymální signalizace regulující morfogenezi zubu". J. Cell Sci. 116 (Pt 9): 1647–8. doi:10.1242 / jcs.00410. PMID 12665545. S2CID 45648812.
- ^ Hardcastle Z, Mo R, Hui CC, Sharpe PT (1998). "Signální dráha SHH ve vývoji zubu: defekty u mutantů Gli2 a Gli3". Rozvoj. 125 (15): 2803–11. PMID 9655803.
- ^ Wolpert L (2015). Zásady rozvoje (5. vydání). Oxford University Press. str. 500. ISBN 978-0-19-967814-3.
- ^ Bellusci S, Furuta Y, Rush MG, Henderson R, Winnier G, Hogan BL (1997). "Zapojení Sonic hedgehog (Shh) do embryonálního růstu plic myší a morfogeneze" (PDF). Rozvoj. 124 (1): 53–63. PMID 9006067.
- ^ A b C d E Pepicelli CV, Lewis PM, McMahon AP (1998). "Sonic ježek reguluje větvící morfogenezi v plicích savců". Aktuální biologie. 8 (19): 1083–6. doi:10.1016 / S0960-9822 (98) 70446-4. PMID 9768363. S2CID 12711144.
- ^ White AC, Xu J, Yin Y, Smith C, Schmid G, Ornitz DM (2006). „Signalizace FGF9 a SHH koordinuje růst a vývoj plic prostřednictvím regulace odlišných mezenchymálních domén“. Rozvoj. 133 (8): 1507–17. doi:10.1242 / dev.02313. PMID 16540513. S2CID 23839558.
- ^ Miura T (2008). Modelování morfogeneze větvení plic. Aktuální témata ve vývojové biologii. 81. str. 291–310. doi:10.1016 / S0070-2153 (07) 81010-6. ISBN 9780123742537. PMID 18023732.
- ^ A b C Kugler MC, Joyner AL, Loomis CA, Munger JS (2015). „Sonic hedgehog signalizace v plicích. Od vývoje k nemoci“. American Journal of Respiratory Cell and Molecular Biology. 52 (1): 1–13. doi:10.1165 / rcmb.2014-0132TR. PMC 4370254. PMID 25068457.
- ^ Cardoso WV, Lü J (2006). „Regulace časné morfogeneze plic: otázky, fakta a kontroverze“. Rozvoj. 133 (9): 1611–24. doi:10.1242 / dev.02310. PMID 16613830. S2CID 18195361.
- ^ Lu N, Chen Y, Wang Z, Chen G, Lin Q, Chen ZY, Li H (2013). "Sonic hedgehog initiates cochlear hair cell regeneration through downregulation of retinoblastoma protein". Biochem. Biophys. Res. Commun. 430 (2): 700–5. doi:10.1016/j.bbrc.2012.11.088. PMC 3579567. PMID 23211596.
- ^ Bumcrot DA, Takada R, McMahon AP (April 1995). "Proteolytic processing yields two secreted forms of sonic hedgehog". Molekulární a buněčná biologie. 15 (4): 2294–303. doi:10.1128/MCB.15.4.2294. PMC 230457. PMID 7891723.
- ^ Ingham PW, Nakano Y, Seger C (červen 2011). "Mechanismy a funkce ježka signalizujícího přes metazoa". Recenze přírody. Genetika. 12 (6): 393–406. doi:10.1038 / nrg2984. PMID 21502959. S2CID 33769324.
- ^ Porter JA, Young KE, Beachy PA (October 1996). "Cholesterol modification of hedgehog signaling proteins in animal development". Věda. 274 (5285): 255–9. Bibcode:1996Sci...274..255P. doi:10.1126/science.274.5285.255. PMID 8824192. S2CID 11125394.
- ^ Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, et al. (Květen 1998). "Identification of a palmitic acid-modified form of human Sonic hedgehog". The Journal of Biological Chemistry. 273 (22): 14037–45. doi:10.1074/jbc.273.22.14037. PMID 9593755. S2CID 22783982.
- ^ Stanton BZ, Peng LF, Maloof N, Nakai K, Wang X, Duffner JL, Taveras KM, Hyman JM, Lee SW, Koehler AN, Chen JK, Fox JL, Mandinova A, Schreiber SL (March 2009). "A small molecule that binds Hedgehog and blocks its signaling in human cells". Nat. Chem. Biol. 5 (3): 154–6. doi:10.1038/nchembio.142. PMC 2770933. PMID 19151731.
- ^ Maclean K (January 2006). "Humour of gene names lost in translation to patients". Příroda. 439 (7074): 266. Bibcode:2006Natur.439..266M. doi:10.1038/439266d. PMID 16421543. S2CID 19861153.
- ^ Cohen MM (July 2006). "Problems in the naming of genes". Dopoledne. J. Med. Genet. A. 140 (13): 1483–4. doi:10.1002/ajmg.a.31264. PMID 16718675. S2CID 221388561.
Další čtení
- Dorus S, Anderson JR, Vallender EJ, Gilbert SL, Zhang L, Chemnick LG, Ryder OA, Li W, Lahn BT (2006). "Sonic Hedgehog, a key development gene, experienced intensified molecular evolution in primates". Lidská molekulární genetika. 15 (13): 2031–7. doi:10,1093 / hmg / ddl123. PMID 16687440.
- Gilbert, Scott F. (2000). Vývojová biologie (6. vydání). Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0-87893-243-6.
- Kim J, Kim P, Hui CC (2001). "The VACTERL association: lessons from the Sonic hedgehog pathway". Klinická genetika. 59 (5): 306–15. doi:10.1034/j.1399-0004.2001.590503.x. PMID 11359461. S2CID 34304310.
- Morton JP, Lewis BC (2007). "SHH signaling and pancreatic cancer: implications for therapy?". Buněčný cyklus. 6 (13): 1553–7. doi:10.4161/cc.6.13.4467. PMID 17611415. S2CID 4670615.
- Mullor JL, Sánchez P, Ruiz i Altaba A (2003). "Pathways and consequences: Hedgehog signaling in human disease". Trends Cell Biol. 12 (12): 562–9. doi:10.1016/S0962-8924(02)02405-4. PMID 12495844.
- Nanni L, Ming JE, Du Y, Hall RK, Aldred M, Bankier A, Muenke M (2001). "SHH mutation is associated with solitary median maxillary central incisor: a study of 13 patients and review of the literature". American Journal of Medical Genetics. 102 (1): 1–10. doi:10.1002/1096-8628(20010722)102:1<1::AID-AJMG1336>3.0.CO;2-U. PMID 11471164.
- Williams JA (2006). "Hedgehog and spinal cord injury". Znalecký posudek na terapeutické cíle. 9 (6): 1137–45. doi:10.1517/14728222.9.6.1137. PMID 16300466. S2CID 5548531.
externí odkazy
- Úvodní článek o SHH na Davidson College
- Rediscovering biology: Unit 7 Genetics of development .. Expert interview transcripts interview with John Incardona PhD .. explanation of the discovery and naming of the sonic hedgehog gene
- ‘Sonic Hedgehog’ sounded funny at first .. New York Times November 12 2006 ..
- Přehled GeneReviews / NCBI / NIH / UW v přehledu Anophthalmia / Microphthalmia
- SHH – sonic hedgehog Americká národní lékařská knihovna
- Přehled všech strukturálních informací dostupných v PDB pro UniProt: Q15465 (Human Sonic hedgehog protein) at the PDBe-KB.
- Přehled všech strukturálních informací dostupných v PDB pro UniProt: Q62226 (Mouse Sonic hedgehog protein) at the PDBe-KB.
Galerie PDB | |
|---|---|
|