Genetika a archeogenetika jižní Asie - Genetics and archaeogenetics of South Asia
Genetika a archeogenetika jižní Asie je studium genetika a archeogenetika z etnické skupiny jižní Asie. Jeho cílem je odhalit tyto skupiny genetická historie. Díky geografické poloze jižní Asie je její biologická rozmanitost důležitá pro studium časného rozšíření anatomicky moderní lidé přes Asie.
Studie založené na mitochondriální DNA (mtDNA ) variace uváděly genetickou jednotu v různých jihoasijských subpopulacích.[1][2][3][4] Závěry studií založených na variacích chromozomů Y a variacích autosomálních DNA byly různé, ačkoli mnoho vědců tvrdí, že většina rodových uzlů fylogenetického stromu všech typů mtDNA pocházela z jižní Asie. Zdá se, že nedávné studie genomu ukazují, že většina jihoasijských obyvatel je potomky dvou hlavních složek předků, z nichž jedna je omezena na jižní Asii (Rodový jihoindický, pocházející z IVC -lidé a původní jihoasijská populace vzdáleně příbuzná s Andamanský ) a další složka (Rodový severoindický) odvozený od lidí IVC a lidí ze stepi, díky čemuž je užší příbuzný lidem v Střední Asie, Západní Asie a Evropa.[5][6][7] Studie z roku 2016 identifikovala další dvě složky předků v Indii na pevnině, označené jako AAA a ATB, které jsou významné pro rakousko-asijsky mluvící kmeny a tibetsko-burmanské mluvčí. Studie také vyvozuje, že populace Andamanské ostrovy souostroví tvoří zřetelný původ, ke kterému „bylo shledáno, že je coancestralem Oceánský populace “.[8] Genomické studie popsaly genetickou krajinu jižní Asie jako směsici západoevropských a východní Asiat exogenní složky, které se mísily s původními jihoasijskými skupinami a vytvořily moderní jižní Asiaté.[9] Složka východoasijských předků zjištěná v Indii je omezena hlavně na konkrétní populace v podhůří Himálaje a severovýchodní části Indie.[10][11]
Bylo zjištěno, že rodový uzel fylogenetického stromu všech mtDNA typy (mitochondriální DNA haploskupiny ) typicky se vyskytující ve střední Asii, západní Asii a Evropě lze také nalézt v jižní Asii s relativně vysokými frekvencemi. Odhadovaná divergence tohoto společného rodového uzlu se odhaduje na něco méně než před 50 000 lety.[12] V Indii jsou hlavní mateřské linie různé M subclades, následovaný R a U podněty. Doby shlukování těchto mitochondriálních haploskupin byly doposud přibližně 50 000 BP.[12]
Hlavní otcovské linie představované Y chromozomy jsou haploskupiny R1a1, R2, H, L a Haploskupina J2.[13] Někteří vědci tvrdili, že Y-DNA Haploskupina R1a1 (M17) je autochtonní Jihoasijský původ.[14][15] Docela běžné jsou však také návrhy středoasijského původu pro R1a1.[16][17]
Přehled
Všechny linie mtDNA a Y-chromozomu mimo Afriku pocházejí ze tří linií zakladatelů:
Všech těchto šest zakladatelských haploskupin lze nalézt v současných populacích Jížní Asie. Kromě toho jsou haploskupiny mtDNA M a haploskupiny Y a chromozomu C a D omezeny na oblast východní jižní Asie. Celý západ euroasijský populace pocházejí z N a R haploskupin mtDNA a F haploskupiny Y-chromozomu.[18]
Endicott a kol. uvádějí, že tato fakta jsou v souladu s hypotézou jediného exodu z východní Afriky před 65 000 lety prostřednictvím a jižní pobřežní trasa, přičemž západoasijské linie se oddělují od jihoasijských linií někde mezi východní / severovýchodní Afrikou a jižní Asií.[19]
Převážná většina markerů genomu jižních Asiatů úzce souvisí Západní Eurasijci a mohou mít původ v Západní Asie nebo Jížní Asie sám.[20]
mtDNA

Nejčastější haploskupiny mtDNA v jižní Asii jsou M, R a U (kde U je potomkem R).[13]
Argumentovat pro dlouhodobější „konkurenční model Y-chromozomu“,[14] Stephen Oppenheimer je přesvědčen, že je velmi sugestivní, že původem Indie je Indie euroasijský mtDNA haploskupin, které nazývá „euroasijské evy“. Podle Oppenheimera je vysoce pravděpodobné, že téměř všechny lidské mateřské linie ve Střední Asii, na Středním východě a v Evropě pocházely pouze ze čtyř linií mtDNA, které vznikly v jižní Asii před 50 000–100 000 lety.[21]
Makrohaploskupina M.
The makrohaploskupina M, který je považován za shluk protoasijských mateřských linií,[12] představuje více než 60% jihoasijské MtDNA.[22]
Makrohaplotyp M v Indii zahrnuje mnoho podskupin, které se hluboce liší od ostatních podnětů ve východní Asii, zejména u mongoloidních populací.[12] Hluboké kořeny M. fylogeneze jasně zjišťují relikty jihoasijských linií ve srovnání s ostatními M liniemi (ve východní Asii a jinde), což naznačuje „in situ“ původ těchto subhaploskupin v jižní Asii, nejpravděpodobněji v Indii. Tyto hluboce zakořeněné linie nejsou jazykově specifické a jsou rozšířeny do všech jazykových skupin v Indii.[22]
Zdá se, že prakticky všechny moderní středoasijské linie MtDNA M patří k východní euroasijské (mongolský ) spíše než jihoasijské podtypy haploskupiny M, což naznačuje, že nedochází k rozsáhlé migraci ze současnosti Turkic - Indie mluvící populace střední Asie. Absence haploskupiny M u Evropanů ve srovnání s její stejně vysokou frekvencí mezi jihoasijskými, východoasijskými a některými středoasijskými populacemi kontrastuje se západoasijskými sklony jihoasijských otcovských linií.[12]
Většina existujících hranic mtDNA v jižní a jihozápadní Asii byla pravděpodobně formována během počátečního osídlení Eurasie anatomicky moderními lidmi.[23]
| Haploskupina | Důležité podřízené subtypy | Populace |
|---|---|---|
| M2 | M2a, M2b | Na celém kontinentu s nízkou přítomností na severozápadě Vrcholem je Bangladéš, Andhra Pradesh, pobřežní Tamil Nadu a Srí Lanka |
| M3 | M3a | Soustředěn do severozápadní Indie Nejvyšší z parsejských Bombajů |
| M4 | M4a | Vrcholy v Pákistánu, Kašmíru a Ándhrapradéši |
| M6 | M6a, M6b | Kašmír a poblíž pobřeží Bengálského zálivu na Srí Lance |
| M18 | V celé jižní Asii Vrchol v Rádžasthánu a Ándhrapradéši | |
| M25 | Mírně časté v Kerale a Maharashtra, ale spíše zřídka jinde v Indii |
Makrohaploskupina R



The makrohaploskupina R (velmi velké a staré dělení na makrohaploskupina N ) je také široce zastoupena a představuje dalších 40% jihoasijské MtDNA. Je to velmi staré a nejdůležitější členění haploskupina U že, zatímco je také přítomen v Západní Eurasie, má několik subclades specifických pro jižní Asii.
Nejdůležitější jihoasijské haploskupiny v rámci R:[23]
| Haploskupina | Populace |
|---|---|
| R2 | Distribuováno široce po subkontinentu |
| R5 | široce distribuován většinou Indie. Vrcholy v pobřežním JZ Indii |
| R6 | rozšířený za nízké ceny po celé Indii. Vrcholy mezi Tamily a Kašmírci |
| Ž | Nalezeno v severozápadních státech. Vrcholy v Gudžarátu, Paňdžábu a Kašmíru, frekvence je jinde nízká. |
Haploskupina U
Haploskupina U je sub-haploskupina makrohaploskupiny R.[23] Distribuce haploskupiny U je zrcadlovým obrazem haploskupiny M: první nebyla dosud popsána u východních Asiatů, ale je častá v evropských populacích i mezi jižními Asijci.[24] Jihoasijské linie U se podstatně liší od linií v Evropě a jejich splynutí se společným předkem také pochází z doby před asi 50 000 lety.[1]
| Haploskupina | Populace |
|---|---|
| U2 * | (A parahaplogroup ) je řídce distribuován speciálně v severní polovině jižní Asie. To je také nalezené v SW Arábii. |
| U2a | vykazuje relativně vysokou hustotu v Pákistánu a SZ Indii, ale také v Karnatace, kde dosahuje vyšší hustoty. |
| U2b | má nejvyšší koncentraci v Uttarpradéši, ale nachází se také na mnoha dalších místech, zejména v Kérale a na Srí Lance. To je také nalezené v Ománu. |
| U2c | je zvláště důležitá v Bangladéši a Západním Bengálsku. |
| U2l | je možná numericky nejdůležitější mezi subclady U v jižní Asii a dosahuje zvláště vysokých koncentrací (přes 10%) v Uttarpradéši, na Srí Lance, v Sindhu a v částech Karnataka. Nějaký význam to má také v Ománu. haploskupina mtDNA U2i je v Bamšádu přezdíván „západoasijský“ et al. studovat, ale „východoasijský (většinou specifický pro Indii)“ v Kivisild et al. studie. |
| U7 | tato haploskupina má významné zastoupení v Gudžarátu, Paňdžábu a Pákistánu. Možná vlast této haploskupiny zahrnuje Gujarat (nejvyšší frekvence, 12%) a Írán, protože odtud její frekvence prudce klesá jak na východ, tak na západ. |
Y chromozom
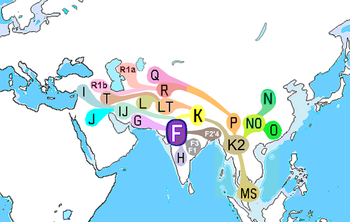
Hlavní haploskupiny DNA jihoasijského Y-chromozomu jsou H, J2, L, R1a1 a R2.[13] Jejich geografický původ je uveden podle posledního stipendia následovně:
| Hlavní jihoasijské Y-chromozomální linie: | H | J2 | L | R1a | R2 |
|---|---|---|---|---|---|
| Basu et al. (2003) | bez komentáře | bez komentáře | bez komentáře | Střední Asie | bez komentáře |
| Kivisild et al. (2003) | Indie | Západní Asie | Indie | Jižní a západní Asie | Jihovýchodní Asie |
| Cordaux et al. (2004) | Indie | Západní nebo střední Asie | Středního východu | Střední Asie | Jihovýchodní Asie |
| Sengupta et al. (2006) | Indie | Střední východ a střední Asie | Jižní Indie | Severní Indie | Severní Indie |
| Thanseem et al. (2006) | Indie | Levant | Střední východ | Jižní a střední Asie | Jižní a střední Asie |
| Sahoo et al. (2006) | Jížní Asie | Blízký východ | Jížní Asie | Jižní nebo západní Asie | Jížní Asie |
| Mirabal et al. (2009) | bez komentáře | bez komentáře | bez komentáře | Severozápadní Indie nebo střední Asie | bez komentáře |
| Zhao et al. (2009) | Indie | Střední východ | Střední východ | Střední Asie nebo západní Eurasie | Střední Asie nebo západní Eurasie |
| Sharma et al. (2009) | bez komentáře | bez komentáře | bez komentáře | Jížní Asie | bez komentáře |
| Thangaraj et al. (2010) | Jížní Asie | Blízký východ | Blízký východ | Jížní Asie | Jížní Asie |
Haploskupina H
Haploskupina H (Y-DNA) se v jižní Asii vyskytuje velmi často. H se dnes zřídka vyskytuje mimo jižní Asii, ale je běžný mezi Romanis, zejména podskupina H-M82. H byl také docela běžný ve starodávných vzorcích Evropy a dodnes se s nízkou frekvencí vyskytuje u Evropanů a Arabů v Evropě Levant. Haploskupina H se často vyskytuje u populací Indie, Srí Lanka, Nepál, Pákistán a Maledivy. Všechny tři pobočky Haploskupina H (Y-DNA) se nacházejí v jižní Asii.
Je to pobočka Haploskupina F a sestupuje z GHIJK rodina. Haploskupina H Předpokládá se, že vznikly v jižní Asii před 30 000 až 40 000 lety.[25] Pravděpodobným místem jeho zavedení je jižní Asie, protože se zde koncentruje. Zdá se, že představuje hlavní haploskupinu Y-chromozomu paleolitických obyvatel jižní Asie. Bylo také prokázáno, že někteří jedinci v jižní Asii patří k mnohem vzácnější subklade H3 (Z5857).[25] Haploskupina H není v žádném případě omezena na konkrétní populace. Například H je posedlý asi 28,8% indoárijských kast.[14][26] a v kmenech asi 25–35%.[16][26]
Haploskupina J2
Haploskupina J2 je v jižní Asii přítomna většinou jako J2a-M410 a J2b-M102 již od neolitu (9500 YBP).[27][28] Klady J2 dosahují špičkových frekvencí v severozápadní a jižní Indii[27] a v jihoindických kastách je 19%, v severoindických kastách 11% a v Pákistánu 12%.[14] v Jižní Indie, přítomnost J2 je vyšší u středních kast 21%, následovaných horními kastami 18,6% a nižšími kastami 14%.[14] Ze skupin kast je nejvyšší frekvence J2-M172 pozorována u tamilštiny Vellalars jižní Indie, na 38,7%.[14] J2 je přítomen také v kmenech[27] a má frekvenci 11% v rakousko-asijských kmenech. Mezi rakousko-asijskými kmeny převládal J2 vyskytuje se u Lodhy (35%).[14] J2 je také přítomen v jižní Indii horský kmen Toda na 38,46%,[29] v Andh kmen z Telangana na 35,19%[16] a v Kmen Kol z Uttarpradéš s frekvencí 33,34%.[30] Bylo zjištěno, že haploskupina J-P209 je běžnější v Indii Šíitští muslimové, z nichž 28,7% patří do haploskupiny J, s 13,7% v J-M410, 10,6% v J-M267 a 4,4% v J2b (Eaaswarkhanth 2009 ).
v Pákistán, nejvyšší frekvence J2-M172 byly pozorovány mezi Parsis na 38,89%, Dravidian mluvení Brahuis na 28,18% a Makrani Balochové na 24%.[31] Vyskytuje se také na 18,18% v Makrani Siddis a 3% v Karnataka Siddis.[31][32]
J2-M172 se nachází v celkové frekvenci 10,3% mezi Sinhálci z Srí Lanka.[33] v Maledivy Bylo shledáno, že 20,6% maledivské populace je haploskupiny J2 pozitivní.[34]
Haploskupina L
Podle Dr. Spencer Wells, L-M20 pochází z Pamírský uzel region v Tádžikistán a migroval do Pákistán a Indie ca. Před 30 000 lety.[35][36][37] Většina ostatních studií však navrhla a Západoasijský původ pro L-M20 a s tím spojená jeho expanze v Údolí Indu (~ 7 000 YBP) do neolitický farmáři.[31][38][16][26][39][40] Existují tři dílčí větve haploskupiny L: L1-M76 (L1a1), L2-M317 (L1b) a L3-M357 (L1a2), které se v jižní Asii nacházejí na různých úrovních.[14]
Indie
Haploskupina L ukazuje čas neolitické expanze.[41] Klad je v indické populaci přítomen s celkovou frekvencí ca. 7–15%.[14][16][42][26] Haploskupina L má vyšší frekvenci mezi jihoindickými kastami (asi 17–19%) a v některých kastách dosahuje až 68% Karnataka ale v severoindických kastách je o něco vzácnější (asi 5–6%).[14] Přítomnost haploskupiny L je u kmenových skupin poměrně vzácná (asi 5,6–7%),[14][16][26] mezi EU a USA však bylo pozorováno mírných 14,6% Chenchus.[33]
Mezi regionálními a sociálními skupinami byly mírné až vysoké frekvence pozorovány v Konkanastha Brahmins (18,6%), Punjabis (12,1%), Gujaratis (10,4%), Lambadis (17.1%), Jats (36.8%)[33][36]
Pákistán
V Pákistánu dosahují subklady L1-M76 a L3-M357 L-M20 celkových frekvencí 5,1%, respektive 6,8%.[14]Haploskupina L3 (M357) se často vyskytuje mezi Burusho (přibližně 12%[43]) a Paštunové (přibližně 7%[43]). Jeho nejvyšší frekvence se nachází na jihozápadě Balúčistán provincie podél Makran pobřeží (28%) do Řeka Indus delta. L3a (PK3) se nachází v přibližně 23% Nuristani na severozápadě Pákistán.[43]
Clade je přítomen ve střední distribuci mezi obecnou pákistánskou populací (přibližně 14%).[44][45]
Srí Lanka
V jedné studii bylo 16% Sinhálců shledáno pozitivními na haploskupinu L-M20.[46] V jiné studii bylo zjištěno, že 18% patří do L1.[33]
Haploskupina R1a1
V jižní Asii byl R1a1 pozorován často s vysokou frekvencí v řadě demografických skupin,[15][47][48] stejně jako s nejvyšší STR rozmanitost, která některé vede k tomu, že je považují za místo původu.[33][30][49]
Zatímco R1a vznikl ca. 22 000[30] na 25 000[50] před lety jeho subclade M417 (R1a1a1) diverzifikoval ca. Před 5800 lety.[50] Distribuce M417-subclades R1-Z282 (včetně R1-Z280)[51] ve střední a východní Evropě a R1-Z93 v Asii[51][50] naznačuje, že R1a1a se v rámci EU diverzifikovala Euroasijské stepi nebo střední východ a Kavkaz kraj.[51] Místo původu těchto subclades hraje roli v debatě o původu Indoevropané.
Indie
v Indie, vysoké procento této haploskupiny je pozorováno v Západní Bengálsko Brahmins (72%)[47] na východ, Gujarat Lohanas (60%)[48] na západ, Khatris (67%)[48] na severu, Iyengar Brahmins (31%) na jihu.[47] Bylo také nalezeno v několika Jihoindický Dravidian -mluvení domorodci včetně Kotasů (41%) z Tamil Nadu[52] Chenchu (26%) a Valmikis z Andhra Pradesh[33] stejně jako Yadav a Kallar z Tamil Nadu což naznačuje, že M17 je v těchto kmenech jižních indiánů rozšířená.[33] Kromě toho studie ukazují vysoké procento v regionálně různorodých skupinách, jako je Manipuris (50%)[48] na extrémní severovýchod a mezi nimi Punjabis (47%)[33] na extrémní severozápad.
Pákistán
V Pákistánu je 71% mezi Mohannou z Provincie Sindh na jih a 46% mezi Baltis z Gilgit-Baltistan na sever.[48]
Srí Lanka
23% z Sinhálci ze vzorku 87 subjektů bylo podle výzkumu z roku 2003 shledáno pozitivní na R1a1a (R-SRY1532).[46]
Maledivy
v Maledivy, 23,8% z Maledivští lidé bylo zjištěno, že jsou pozitivní na R1a1a (M17).[34]
Nepál
Lidé v Terai Kraj, Nepál ukázat R1a1a na 69%.[53]
Haploskupina R2
V jižní Asii je frekvence R2 a R2a linie se pohybuje kolem 10–15% v Indii a Srí Lanka a 7–8% v Pákistánu. Nejméně 90% osob R-M124 se nachází v jižní Asii.[54] Uvádí se také v Kavkaz a Střední Asie při nižší frekvenci. Genetická studie Mondal et al. 2017 uzavřel tuto haploskupinu Haploskupina R2 pocházel ze severní Indie a byl již přítomen před stepní migrací.[55]
Indie
Mezi regionálními skupinami se vyskytuje mezi Západ Bengálci (23%), Nové Dillí Hinduisté (20%), Punjabis (5%) a Gujaratis (3%).[33] Mezi kmenovými skupinami byl Karmalis z Západní Bengálsko ukázal nejvyšší na 100%[15] následován Lodhas (43%)[56] na východ, zatímco Bhil z Gudžarát na západě bylo 18%,[30] Tharusi severu to ukázalo na 17%,[4] Chenchu a Pallan jihu činily 20%, respektive 14%.[15][14] Mezi kastovními skupinami je vysoké procento znázorněno Jaunpur Kshatriyas (87%), kamma (73%), Bihar Yadav (50%), Khandayat (46%) a Kallar (44%).[15]
U mnoha je také výrazně vysoká Bráhman skupiny včetně Punjabi Brahmins (25%), Bengálské Brahmins (22%), Konkanastha Brahminové (20%), Chaturvedis (32%), Bhargavové (32%), Kašmírské Pandits (14%) a Lingayat Brahminové (30%).[30][17][4][15]
Severoindičtí muslimové mají frekvenci 19% (Sunni ) a 13% (Shia ),[17] zatímco Muslim Dawoodi Bohra v západním státě Gudžarát mají frekvenci 16% a Mappila muslimové jižní Indie mají frekvenci 5%.[57]
Pákistán
Haploskupina R2 se vyskytuje u 14% Burusho lidé.[43] Mezi Hunza lidé nachází se na 18%, zatímco Parsis ukázat to na 20%. To je také nalezené v severovýchodní části Afghánistán.[Citace je zapotřebí ]
Srí Lanka
38% Sinhálština Srí Lanky byly podle výzkumu z roku 2003 shledány pozitivními na R2.[33]
Maledivy
12% z Maledivští lidé Malediv má R2.[34]
Nepál
V Nepálu se procenta R2 pohybují od 2% do 26% v různých skupinách podle různých studií. Newars vykazují výrazně vysokou frekvenci 26%, zatímco lidé z Káthmándú ukázat to na 10%.
Rekonstrukce historie jihoasijské populace
The Konsorcium pro variaci indického genomu (2008), rozděluje populaci jižní Asie do čtyř etnolingvistických skupin: Indoevropský, Dravidian, Tibeto-Burman a Rakousko-asijský.[58][59][60][61][62] Studie molekulární antropologie používají tři různé typy markerů: variace mitochondriální DNA (mtDNA), která je mateřsky zděděná a vysoce polymorfní, variace chromozomu Y, která zahrnuje uniparentální přenos podél mužských linií, a variace autosomální DNA.[4]:04
variace mtDNA
Většina studií založených na variacích mtDNA uvádí genetickou jednotu jihoasijských populací napříč jazykovými, kastovými a kmenovými skupinami.[1][2][3] Je pravděpodobné, že haploskupina M byla do Asie přivezena z východní Afriky jižní cestou nejranější migrační vlnou před 78 000 lety.[1]
Podle Kivisild et al. (1999), „Menší přesahy s liniemi popsanými v jiných euroasijských populacích jasně ukazují, že nedávná imigrace měla velmi malý dopad na vrozenou strukturu mateřských genofond jihoasijských. Navzdory variacím zjištěným v Indii tyto populace pocházejí z omezeného počtu počtů zakladatelů. Tyto linie byly s největší pravděpodobností zavedeny do jižní Asie během středního paleolitu, před osídlením Evropy před 48 000 lety a možná i Starým světem obecně. “[1] Basu a kol. (2003) také zdůrazňuje základní jednotu ženských linií v Indii.[42]
Y Variace chromozomu
Závěry založené na variaci chromozomu Y byly rozmanitější než závěry založené na variaci mtDNA. Zatímco Kivisild a kol. (2003) navrhuje starodávné a sdílené genetické dědictví mužských linií v jižní Asii, Bamshad et al. (2001) navrhuje afinitu mezi jihoasijskými mužskými liniemi a západními Eurasijci úměrně k vyšší kastovní pozici a přibližuje populace vyšších kast v jižních indických státech Východní Evropané.[63]
Basu a kol. (2003) dochází k závěru, že rakousko-asijské kmenové populace vstoupily do Indie nejprve ze severozápadního koridoru a mnohem později některé z nich severovýchodním koridorem.[42] Zatímco Kumar et al. (2007) analyzovali 25 jihoasijských rakousko-asijských kmenů a našli silnou otcovskou genetickou vazbu mezi subjazykovými skupinami jihoasijských rakousko-asijských populací.[56] Mukherjee a kol. (2001) umisťuje Pákistánce a severní Indy mezi populace západní Asie a Střední Asie,[64] vzhledem k tomu, že Cordaux a kol. (2004) tvrdí, že indické populace kast jsou blíže populacím střední Asie.[26] Sahoo a kol. (2006) a Sengupta et al. (2006) naznačují, že populace indických kast nebyly podrobeny žádným nedávným příměsím.[14][15] Sanghamitra Sahoo uzavírá svoji studii:[15]
Na základě současných důkazů není nutné hledat za jižní Asií původ otcovského dědictví většiny Indů v době nástupu osídleného zemědělství. Vytrvalá koncepce lidí, jazyka a zemědělství, kteří společně přicházejí do Indie severozápadním koridorem, neobstojí v úzké kontrole. Nedávná tvrzení o spojení haploskupin J2, L, R1a a R2 se současným původem u většiny otcovských linií indických kast mimo jižní Asii jsou odmítnuta, ačkoli naše zjištění podporují místní původ haploskupin F * a H. Z ostatních pouze J2 naznačuje jednoznačný nedávný externí příspěvek, a to ze západní Asie, spíše než ze střední Asie. Současná rozdělení frekvencí haploskupin jsou, s výjimkou linií, převážně řízena geografickými, nikoli kulturními determinanty. Je ironií, že právě na severovýchodě Indie, mezi skupinami TB, existují jednoznačné důkazy o rozsáhlé demické difúzi vysledovatelné podle genů, kultury a jazyka, ale zjevně ne podle zemědělství.
Analýza nejbližšího souseda provedená Mondalem a kol. 2017 dospěl k závěru, že indické linie Y jsou blízko jižního evropský populace a doba divergence mezi dvěma předcházejícími stepními migracemi. ":[65]
Tyto výsledky naznačují, že evropský původ v indických populacích může být mnohem starší a složitější, než se předpokládalo, a může pocházet z první vlny zemědělců nebo dokonce dříve
— Mondal a kol. 2017
Variace autozomální DNA
AASI-ANI-ASI
Také se měnily výsledky studií založených na variacích autozomální DNA. Ve velké studii (2009) používající více než 500 000 bialelických autozomálních markerů Reich vyslovil hypotézu, že moderní jihoasijská populace byla výsledkem příměsi mezi dvěma geneticky odlišnými populacemi předků z doby po holocénu. Tyto dvě „rekonstruované“ starověké populace nazval „rodové jižní indiány“ (ASI) a „původní rodové indiány“ (ANI). Podle Reicha: „ANI předci jsou významně vyšší v indoevropských než dravidských mluvčích, což naznačuje, že rodová ASI mohla před smícháním s ANI mluvit dravidským jazykem.“ Zatímco ANI je geneticky blízká Středním východům, Středoasijcům a Evropanům, ASI není úzce spjata se skupinami mimo subkontinent. Protože není k dispozici žádná „ASI“ starodávná DNA, domorodá Andamanský Onge jsou používány jako (nedokonalý) zástupce ASI (podle Reicha et al., Andamanese, ačkoli odlišný od nich, je nejbližší žijící populace k ASI). Podle Reicha a kol. Se původ ANI i ASI vyskytuje na celém subkontinentu (v severní i jižní Indii) v různých poměrech a že „původ ANI se pohybuje v rozmezí 39–71% v Indii a je vyšší v tradičně vyšších kastovní a indoevropští mluvčí. “[66]
Moorjani a kol. 2013 uvádí, že ASI, i když úzce nesouvisí s žádnou živou skupinou, „souvisí (vzdáleně) s původními Andamanskými ostrovany“. Moorjani a kol. také naznačují možný tok genů do Andamanese z populace související s ASI. Studie dospěla k závěru, že „téměř všechny skupiny, které hovoří indoevropskými nebo dravidskými jazyky, leží na gradientu různé příbuznosti se západoevropskými jazyky v PCA (dále jen„ indický jazyk “)“.[67]
Studie z roku 2013 využívající jedno-nukleotidový polymorfismus (SNP) ukazuje, že genom andamanských lidí (Onge) je blíže genomu jiných oceánských skupin Negrito než jižní Asiatům.[68]
Podle Basu et al. 2016, další analýza odhalila, že genomovou strukturu kontinentálních indických populací lze nejlépe vysvětlit příspěvky od čtyř předků. Kromě ANI a ASI Basu et. al (2016) identifikoval dvě součásti předků v Indii na pevnině, které jsou významné pro rakousko-asijsky mluvící kmeny a tibetsko-barmské mluvčí, které označili jako AAA (pro „Ancestral Austro-Asiatic“) a ATB (pro „Ancestral Tibeto“ -Burman "). Studie také vyvozuje, že populace Andamanské ostrovy souostroví tvoří zřetelný původ, ke kterému „bylo shledáno, že je coancestralem Oceánský populace “.[8]
Cline příměsi mezi liniemi ANI a ASI je datována do období c. 4,2–1,9 kya od Moorjani et al. (2013), což odpovídá indické době bronzové a je autory spojeno s procesem deurbanizace Civilizace Indus Valley a přesun populace do systému Gangetic v počínající indické době železné.[6] Basu a kol. (2003) naznačuje, že „dravidské kmeny byly pravděpodobně rozšířeny po celé Indii před příchodem indoevropsky mluvících nomádů“ a že „tvorba populací štěpením, která vedla k efektům zakladatele a driftu, zanechala své stopy na genetických strukturách současníků populace “.[42] Genetik PP Majumder (2010) nedávno tvrdil, že nálezy Reich et al. (2009) jsou v pozoruhodné shodě s předchozími výzkumy využívajícími mtDNA a Y-DNA:[69]
Populace ve střední Asii mají být hlavními přispěvateli do indického genofondu, zejména do severoindického genofondu, a migranti se údajně přestěhovali do Indie prostřednictvím dnešního Afghánistánu a Pákistánu. Použitím mitochondriální DNA Údaje o variacích shromážděné z různých studií jsme prokázali, že populace Střední Asie a Pákistánu vykazují nejnižší koeficient genetické diferenciace u severoindických populací, vyšší diferenciaci u jihoindických populací a nejvyšší u severovýchodních indických populací. Populace severní Indie jsou geneticky blíže ke středoasijským populacím než populace jiných zeměpisných oblastí Indie .... V souladu s výše uvedenými poznatky nedávná studie používající více než 500 000 bialelických autosomálních markerů našla gradient od severu k jihu s genetickou blízkostí indických populací k západním Eurasijcům. Tato vlastnost pravděpodobně souvisí s proporcemi předků odvozených ze západoasijského genofondu, který je, jak ukázala tato studie, větší u populací obývajících severní Indii než u populací obývajících jižní Indii.
Chaubey a kol. V roce 2015 byla zjištěna výrazná východoasijská předková složka, omezená hlavně na konkrétní populace v podhůří Himálaje a severovýchodní části Indie. Nejvyšší frekvence této složky je pozorována u tibeto-barmských mluvících skupin na severovýchodě a byla zjištěna také v andamanských populacích, přičemž výrazná přítomnost je mezi austroasiatickými mluvčími. Zjistilo se, že v indoevropských a dravidských mluvčích do značné míry chybí, s výjimkou některých konkrétních etnických skupin žijících v podhůří Himálaje a ve střední a jižní Indii.[10] Vědci však navrhli, že východoasijský původ (představovaný Hanem) měřený ve studovaných andamanských skupinách může ve skutečnosti odrážet zachycení spřízněnosti Andamanese s Melanésany a malajskými Negrity (spíše než s pravou východoasijskou příměsí),[10] jako předchozí studie Chaubey et al. navrhl „hluboký společný původ“ mezi Andamanese, Melanésijci a dalšími skupinami Negrito (stejně jako jižní Asiaté),[10] a spřízněnost mezi Negritosem z jihovýchodní Asie a Melanésijci (stejně jako andamanskými) s východoasijskými.[68]
Lazaridis a kol. (2016) uvádí: „Demografický dopad populací souvisejících se stepí na jižní Asii byl značný, jelikož Mala, jihoindický Dalit populace s minimem severoindických předků (ANI) podél „indické linie“ takového původu má ~ 18% stepní předky, zatímco Kalash z Pákistánu je odvozeno, že mají ~ 50% předků souvisejících se stepemi. “[70] Studie Lazaridis et al. Z roku 2016 odhaduje (6,5–50,2%) příměs související s stepí v jižních Asijcích. Lazaridis a kol. dále konstatuje, že „užitečným směrem budoucího výzkumu je komplexnější vzorkování starověké DNA ze stepních populací i populací ve střední Asii (na východ od Íránu a na jih od stepi), což může odhalit bližší zdroje ANI než ti, o nichž se zde uvažuje, a jižní Asie, aby přímo určili trajektorii populačních změn v oblasti.
Pathak a kol. 2018 dospěl k závěru, že Indoevropský mluvčí Gangetické pláně a Dravidian reproduktory mají významné Yamnaya raně střední doba bronzová (Steppe_EMBA) předky, ale žádný předky ze střední a pozdní doby bronzové (Steppe_MLBA). Na druhé straně populace „severozápadní Indie a Pákistánu“ (PNWI) vykazovaly významné předky Steppe_MLBA spolu s předky Yamnaya (Steppe_EMBA). Studie také poznamenala, že starověké jihoasijské vzorky měly významně vyšší Steppe_MLBA než Steppe_EMBA (nebo Yamnaya). Studie rovněž naznačila, že Rors lze použít jako proxy pro ANI.[71]
David Reich ve své knize z roku 2018 Kdo jsme a jak jsme se sem dostali uvádí, že analýzy z roku 2016 zjistily, že ASI má značné množství složky původu pocházející z íránských farmářů (asi 25% jejich původu), přičemž zbývajících 75% jejich původu pochází od původních jihoasijských lovců a sběračů. Dodává, že ASI byly nepravděpodobné místní lovci a sběrači jižní Asie, jak již bylo dříve stanoveno, ale populace odpovědná za šíření zemědělství po jižní Asii. V případě ANI je íránský původ farmářů 50%, zbytek tvoří stepní skupiny související s Yamnayou.[72]
Narasimhan a kol. (2018), podobně k závěru, že ANI a ASI byly vytvořeny ve 2. tisíciletí před naším letopočtem.[73] Předcházela jim směs AASI (starodávní předkové z jižní Indie, tj. Lovci a sběrači, kteří sdíleli společný kořen s Andamanese); a íránští zemědělci, kteří přijeli do Indie ca. 4700–3 000 př. N. L. A „muselo se dostat do údolí Indu do 4. tisíciletí př. N. L.“.[73] Podle Narasimhana a kol. Tato smíšená populace, která pravděpodobně pocházela z civilizace údolí Indu, „přispěla ve velkých poměrech k ANI i ASI“, které se formovaly během 2. tisíciletí př. N. L. ANI vzniklo ze směsi „Periferie Indussouvisející skupiny "a migranti ze stepi, zatímco ASI byla vytvořena z"Periferie Indus„související skupiny“, které se přesunuly na jih a dále se mísily s místními lovci a sběrači. Předpokládá se, že původ populace ASI měl v průměru asi 73% z AASI a 27% od íránských zemědělců. Narasimhan a kol. pozorují, že vzorky z skupina periferií Indu jsou vždy směsí dvou stejných proximálních zdrojů původu AASI a předků souvisejících s íránským zemědělcem; „jeden z indiánských periferních jedinců má ~ 42% původ AASI a další dva jedinci mají ~ 14–18% původ AASI“ (přičemž zbytek jejich předků pochází z populace spojené s íránským zemědělcem).[73] Autoři zjistili, že domorodí lovci a sběrači AASI představují divergentní větev, která se odštěpila přibližně ve stejné době, kdy se od sebe oddělili východoasijští, ongeovští (andamanští) a australští domorodí předkové. Z toho vyvodil, že „v podstatě veškerý původ dnešních východních a jižních Asiatů (před příměsí západoasijských v jižních Asijcích) pochází z jediného rozšíření na východ, které v krátkém čase vedlo k liniím vedoucím k AASI , Východní Asiaté, Onge a Australané.[73]
Genetická studie od Yelmen et al. (2019) ukazuje, že moderní jihoasijské populace jsou obecně nejblíže západoevropským. Došli k závěru, že moderní jižní Asiaté jsou v zásadě směsí nativní jihoasijské genetické složky a později přicházející západo-euroasijské složky (odvozené jak ze západní Asie, tak ze západních stepí). Autoři také argumentují, že původní jihoasijská genetická složka je odlišná od andamanské, a že andamanští jsou tedy nedokonalým zástupcem. Tato složka (pokud ji představuje Andamanese Onge) nebyla u severního Inda detekována Gudžarátština, a proto se navrhuje, aby jihoindický kmen Paniya lidé (skupina převážně ASI předků) by sloužili jako lepší zástupce než Andamanese (Onge) pro „domorodou jihoasijskou“ složku v moderních jižních Asijcích, protože Paniya jsou přímo odvozeny od domorodců z jižní Asie (spíše než vzdáleně) související s nimi jako s Onge).[9]
Dvě genetické studie (Shinde et al. 2019 a Narasimhan et al. 2019), které analyzovaly pozůstatky civilizace v údolí Indu (části severozápadní Indie a východního Pákistánu z doby bronzové), zjistily, že mají směs předků: Shinde et al. zjistili, že jejich vzorky mají asi 50–98% jejich genomu od lidí souvisejících s ranými íránskými farmáři a od 2–50% jejich genomu od původních jihoasijských lovců a sběračů sdílejících společný původ s andamanskými, s íránskými příbuznými původ je v průměru převládající. Vzorky analyzované Narasimhanem a kol. měl 45–82% předků souvisejících s íránskými farmáři a 11–50% AASI (neboli předky lovců a sběračů související s Andamanese). Analyzované vzorky obou studií mají malý až žádný z „Stepní původ "Složka spojená s pozdějšími indoevropskými migracemi do Indie. Autoři zjistili, že příslušná množství těchto předků se mezi jednotlivci významně lišila, a dospěli k závěru, že k získání úplného obrazu o historii indické populace je zapotřebí více vzorků."[74][75]
Genetická vzdálenost mezi kastovními skupinami a kmeny
Studie Watkins et al. (2005) a Kivisild et al. (2003) na základě autosomálních markerů dochází k závěru, že indická kasta a kmenové populace mají společný původ.[33][76] Reddy a kol. (2005) zjistili poměrně rovnoměrné rozdělení frekvencí alel mezi kastovními skupinami jižních Andhra Pradesh, ale výrazně větší genetická vzdálenost mezi kastovními skupinami a kmeny genetická izolace kmenů a kast.[77]
Viswanathan a kol. (2004) ve studii o genetické struktuře a příbuznosti kmenových populací v jižní Indii dochází k závěru, "Genetická diferenciace byla vysoká a genetické vzdálenosti nebyly významně korelovány s geografickými vzdálenostmi. Genetický drift proto pravděpodobně hrál významnou roli při formování vzorů genetických variací pozorovaných v populacích kmenů na jihu Indie. V opačném případě analýzy populačních vztahů ukázaly, že všechny indické a jihoasijské populace jsou si stále navzájem podobné, bez ohledu na fenotypové charakteristiky, a nevykazují žádnou zvláštní afinitu k Afričanům. Dospěli jsme k závěru, že fenotypové podobnosti některých indických skupin s africkými nedělej odrážejí blízký vztah mezi těmito skupinami, ale jsou lépe vysvětleny konvergence."[78]
Studie z roku 2011 zveřejněná v American Journal of Human Genetics[5] naznačuje, že indické komponenty předků jsou výsledkem složitější demografické historie, než se dříve myslelo. Podle výzkumníků v jižní Asii jsou uloženy dvě hlavní složky předků, z nichž jedna se šíří se srovnatelnou frekvencí a genetickou rozmanitostí v populacích střední Asie, západní Asie a Evropy; druhá složka je více omezena na jižní Asii. However, if one were to rule out the possibility of a large-scale Indo-Aryan migration, these findings suggest that the genetic affinities of both Indian ancestral components are the result of multiple gene flows over the course of thousands of years.[5]
A genetic study published in 2020 in the Journal ScienceDirect by Rowold et al. found that the great majority of Indičtí lidé ("caste people"), which make up about 91,4%, are closely related to West-Eurasians (Evropané a Středního východu ). Northern Africans (samplified by Egypťané ) were part of this cluster but shifted towards Subsaharští Afričané. Východní Asiaté a Jihovýchodní Asiaté were most distant to both West-Eurasians and Sub-Saharan Africans and formed a completely separate cluster. Interesingly, non-caste populations of India (samplified by the Soligové, jeden z Adivasi ) which make up about 8,6% of Indians population, were found to be shifted towards Sub-Saharan Africans and share specific alleles. A link to certain Oceanians was also detected, suggesting ancient gene-flow.[79]
Moreover, the presence of these Y-STR profiles in several sub-Saharan populations and conspicuous absence from the other Eurasian collections suggest a unique genetic connection between Indian tribal groups and sub-Saharan Africans.
— Rowold et al. 2020
Viz také
- Archeogenetika
- Etnické skupiny jižní Asie
- List of ethnolinguistic regions of South Asia
- Peopling of India
- Y-DNA haploskupiny v populacích jižní Asie
- Genetické studie na lidech Gujarati
Reference
- ^ A b C d E Kivisild T, Kaldma K, Metspalu M, Parik J, Papiha S, Villems R (1999). "The Place of the Indian Mitochondrial DNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World". Genomic Diversity. str. 135–152. doi:10.1007/978-1-4615-4263-6_11. ISBN 978-1-4613-6914-1.CS1 maint: ref = harv (odkaz)
- ^ A b Baig MM, Khan AA, Kulkarni KM (September 2004). "Mitochondrial DNA diversity in tribal and caste groups of Maharashtra (India) and its implication on their genetic origins". Annals of Human Genetics. 68 (Pt 5): 453–60. doi:10.1046/j.1529-8817.2004.00108.x. PMID 15469422. S2CID 23032872.
- ^ A b Singh AK (2007). Science & Technology For Upsc. Tata McGraw-Hill Education. p. 595. ISBN 978-0-07-065548-5. Archivováno z původního dne 3. ledna 2014. Citováno 24. května 2016.
- ^ A b C d Tripathy, Vikal; Nirmala, A .; Reddy, B. Mohan (4 September 2017). "Trends in Molecular Anthropological Studies in India". International Journal of Human Genetics. 8 (1–2): 1–20. doi:10.1080/09723757.2008.11886015. S2CID 12763485.
- ^ A b C Metspalu M, Romero IG, Yunusbayev B, Chaubey G, Mallick CB, Hudjashov G, et al. (Prosinec 2011). "Shared and unique components of human population structure and genome-wide signals of positive selection in South Asia". American Journal of Human Genetics. 89 (6): 731–44. doi:10.1016/j.ajhg.2011.11.010. PMC 3234374. PMID 22152676.
- ^ A b Moorjani P, Thangaraj K, Patterson N, Lipson M, Loh PR, Govindaraj P, et al. (Září 2013). "Genetic evidence for recent population mixture in India". American Journal of Human Genetics. 93 (3): 422–38. doi:10.1016/j.ajhg.2013.07.006. PMC 3769933. PMID 23932107.
- ^ Silva M, Oliveira M, Vieira D, Brandão A, Rito T, Pereira JB, et al. (Březen 2017). "A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals". BMC Evoluční biologie. 17 (1): 88. doi:10.1186/s12862-017-0936-9. PMC 5364613. PMID 28335724.
- ^ A b Basu A, Sarkar-Roy N, Majumder PP (February 2016). "Genomic reconstruction of the history of extant populations of India reveals five distinct ancestral components and a complex structure". Sborník Národní akademie věd Spojených států amerických. 113 (6): 1594–9. Bibcode:2016PNAS..113.1594B. doi:10.1073/pnas.1513197113. PMC 4760789. PMID 26811443.
- ^ A b Yelmen, Burak; Mondal, Mayukh; Marnetto, Davide; Pathak, Ajai K.; Montinaro, Francesco; Gallego Romero, Irene; Kivisild, Toomas; Metspalu, Mait; Pagani, Luca (1 August 2019). "Ancestry-Specific Analyses Reveal Differential Demographic Histories and Opposite Selective Pressures in Modern South Asian Populations". Molekulární biologie a evoluce. 36 (8): 1628–1642. doi:10.1093/molbev/msz037. ISSN 0737-4038. PMC 6657728. PMID 30952160.
- ^ A b C d Chaubey, Gyaneshwer (January 2015). „Východoasijský původ v Indii“ (PDF). Indian Journal of Physical Anthropology and Human Genetics. 34 (2): 193–199.
Here the analysis of genome wide data on Indian and East/Southeast Asian demonstrated their restricted distinctive ancestry in India mainly running along the foothills of Himalaya and northeastern part.
- ^ Chaubey, Gyaneshwer; Kadian, Anurag; Bala, Saroj; Rao, Vadlamudi Raghavendra (10 June 2015). "Genetic Affinity of the Bhil, Kol and Gond Mentioned in Epic Ramayana". PLOS ONE. 10 (6): e0127655. Bibcode:2015PLoSO..1027655C. doi:10.1371/journal.pone.0127655. ISSN 1932-6203. PMC 4465503. PMID 26061398. S2CID 848806.
- ^ A b C d E Kivisild T, Papiha SS, Rootsi S, Parik J, Kaldma K, Reidla M, et al. (2000), An Indian Ancestry: a Key for Understanding Human Diversity in Europe and Beyond (PDF), McDonald Institute Monographs, archivováno (PDF) z původního dne 19. února 2006, vyvoláno 11. listopadu 2005
- ^ A b C "Y Haplogroups of the World, 2005, McDonald" (PDF). Archivovány od originál (PDF) on 28 July 2004. Citováno 24. října 2006.
- ^ A b C d E F G h i j k l m n Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE a kol. (Únor 2006). „Polarita a dočasnost distribucí y-chromozomů s vysokým rozlišením v Indii identifikuje domorodé i exogenní expanze a odhaluje menší genetický vliv středoasijských pastevců.“. American Journal of Human Genetics. 78 (2): 202–21. doi:10.1086/499411. PMC 1380230. PMID 16400607.
- ^ A b C d E F G h Sahoo S, Singh A, Himabindu G, Banerjee J, Sitalaximi T, Gaikwad S a kol. (Leden 2006). „Pravěk indických chromozomů Y: hodnocení scénářů demické difúze“. Sborník Národní akademie věd Spojených států amerických. 103 (4): 843–8. Bibcode:2006PNAS..103..843S. doi:10.1073 / pnas.0507714103. PMC 1347984. PMID 16415161.
- ^ A b C d E F Thanseem I, Thangaraj K, Chaubey G, Singh VK, Bhaskar LV, Reddy BM, et al. (Srpen 2006). „Genetické spříznění mezi nižšími kastami a kmenovými skupinami Indie: závěr z chromozomu Y a mitochondriální DNA“. Genetika BMC. 7: 42. doi:10.1186/1471-2156-7-42. PMC 1569435. PMID 16893451.
- ^ A b C Zhao Z, Khan F, Borkar M, Herrera R, Agrawal S (2009). "Presence of three different paternal lineages among North Indians: a study of 560 Y chromosomes". Annals of Human Biology. 36 (1): 46–59. doi:10.1080/03014460802558522. PMC 2755252. PMID 19058044.
- ^ Endicott, Metspalu & Kivisild 2007, str. 231.
- ^ Endicott, Metspalu & Kivisild 2007, str. 234–235.
- ^ Das, Ranajit; Upadhyai, Priyanka (25 November 2016). "Tracing the biogeographical origin of South Asian populations using DNA SatNav". bioRxiv: 089466. doi:10.1101/089466. S2CID 88966532.
Our hypothesis is supported by archaeological, linguistic and genetic evidences that suggest that there were two prominent waves of immigrations to India. A majority of the Early Caucasoids were proto-Dravidian language speakers that migrated to India putatively ~ 6000 YBP.
- ^ Oppenheimer 2003[stránka potřebná ]
- ^ A b Puente XS, Velasco G, Gutiérrez-Fernández A, Bertranpetit J, King MC, López-Otín C (January 2006). "Comparative analysis of cancer genes in the human and chimpanzee genomes". BMC Genomics. 7: 15. doi:10.1186/1471-2164-7-15. PMC 1382208. PMID 16438707.
- ^ A b C Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (Srpen 2004). „Většina existujících hranic mtDNA v jižní a jihozápadní Asii byla pravděpodobně formována během počátečního osídlení Eurasie anatomicky moderními lidmi“. Genetika BMC. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- ^ Kivisild, T .; Bamshad, M.J.; Kaldma, K .; Metspalu, M .; Metspalu, E .; Reidla, M .; Laos, S.; Parik, J .; Watkins, W.S.; Dixon, M.E.; Papiha, S.S.; Mastana, S.S.; Mir, M.R.; Ferak, V.; Villems, R. (November 1999). "Deep common ancestry of Indian and western-Eurasian mitochondrial DNA lineages". Aktuální biologie. 9 (22): 1331–1334. doi:10.1016/s0960-9822(00)80057-3. PMID 10574762. S2CID 2821966.
- ^ A b "Y-DNA Haplogroup H and its Subclades – 2015". Archivováno z původního dne 1. listopadu 2015. Citováno 11. října 2015.
- ^ A b C d E F Cordaux R, Aunger R, Bentley G, Nasidze I, Sirajuddin SM, Stoneking M (February 2004). "Nezávislý původ indické kasty a kmenových otcovských linií". Aktuální biologie. 14 (3): 231–5. doi:10.1016 / j.cub.2004.01.024. PMID 14761656. S2CID 5721248.
- ^ A b C Singh, Sakshi; Singh, Ashish; Rajkumar, Raja; Sampath Kumar, Katakam; Kadarkarai Samy, Subburaj; Nizamuddin, Sheikh; Singh, Amita; Ahmed Sheikh, Shahnawaz; Peddada, Vidya; Khanna, Vinee; Veeraiah, Pandichelvam; Pandit, Aridaman; Chaubey, Gyaneshwer; Singh, Lalji; Thangaraj, Kumarasamy (12 January 2016). "Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup". Vědecké zprávy. 6 (1): 19157. Bibcode:2016NatSR...619157S. doi:10.1038/srep19157. ISSN 2045-2322. PMC 4709632. PMID 26754573.
- ^ Herrera, Rene J .; Garcia-Bertrand, Ralph (2018). Ancestral DNA, Human Origins, and Migrations. Akademický tisk. p. 250. ISBN 978-0-12-804128-4.
- ^ Arunkumar G, Soria-Hernanz DF, Kavitha VJ, Arun VS, Syama A, Ashokan KS, et al. (2012). "Population differentiation of southern Indian male lineages correlates with agricultural expansions predating the caste system". PLOS ONE. 7 (11): e50269. Bibcode:2012PLoSO...750269A. doi:10.1371/journal.pone.0050269. PMC 3508930. PMID 23209694.
- ^ A b C d E Sharma S, Rai E, Sharma P, Jena M, Singh S, Darvishi K, et al. (Leden 2009). "The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system". Journal of Human Genetics. 54 (1): 47–55. doi:10.1038 / jhg.2008.2. PMID 19158816. S2CID 22162114.
- ^ A b C Qamar R, Ayub Q, Mohyuddin A, Helgason A, Mazhar K, Mansoor A, et al. (Květen 2002). "Y-chromosomal DNA variation in Pakistan". American Journal of Human Genetics. 70 (5): 1107–24. doi:10.1086/339929. PMC 447589. PMID 11898125.
- ^ Shah AM, Tamang R, Moorjani P, Rani DS, Govindaraj P, Kulkarni G, et al. (Červenec 2011). "Indian Siddis: African descendants with Indian admixture". American Journal of Human Genetics. 89 (1): 154–61. doi:10.1016 / j.ajhg.2011.05.030. PMC 3135801. PMID 21741027.
- ^ A b C d E F G h i j k Kivisild (2003)
- ^ A b C Pijpe, Jeroen; de Voogt, Alex; van Oven, Mannis; Henneman, Peter; van der Gaag, Kristiaan J .; Kayser, Manfred; de Knijff, Peter (21 March 2013). "Indian ocean crossroads: Human genetic origin and population structure in the maldives". American Journal of Physical Anthropology. 151 (1): 58–67. doi:10.1002/ajpa.22256. ISSN 0002-9483. PMC 3652038. PMID 23526367.
- ^ Wells, Spencer (2007). Hluboký původ: uvnitř Genografického projektu. Washington, DC: National Geographic. ISBN 978-1426201189.
- ^ A b Mahal DG, Matsoukas IG (20 September 2017). „Diverzita haploskupin Y-STR v populaci Jat odhaluje několik různých starověkých původů“. Frontiers in Genetics. 8: 121. doi:10.3389 / fgene.2017.00121. PMC 5611447. PMID 28979290.
- ^ Spencer Wells (2003), The Journey of Man. A Genetic Odyssey. New Delhi: Penguin Books India, p. 167
- ^ Zhao Z, Khan F, Borkar M, Herrera R, Agrawal S (2009). "Presence of three different paternal lineages among North Indians: a study of 560 Y chromosomes". Annals of Human Biology. 36 (1): 46–59. doi:10.1080/03014460802558522. PMC 2755252. PMID 19058044.
- ^ McElreavey K, Quintana-Murci L (2005). „Perspektiva populační genetiky v údolí Indu prostřednictvím uniperarentálně zděděných markerů“. Annals of Human Biology. 32 (2): 154–62. doi:10.1080/03014460500076223. PMID 16096211. S2CID 109014.
- ^ Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK, et al. (Prosinec 2010). "The influence of natural barriers in shaping the genetic structure of Maharashtra populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. doi:10.1371 / journal.pone.0015283. PMC 3004917. PMID 21187967.
- ^ Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK, et al. (Prosinec 2010). Cordaux R (ed.). "The influence of natural barriers in shaping the genetic structure of Maharashtra populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. doi:10.1371 / journal.pone.0015283. PMC 3004917. PMID 21187967.
- ^ A b C d Basu A, Mukherjee N, Roy S, Sengupta S, Banerjee S, Chakraborty M, et al. (Říjen 2003). "Ethnic India: a genomic view, with special reference to peopling and structure". Výzkum genomu. 13 (10): 2277–90. doi:10,1101 / gr. 1413403. PMC 403703. PMID 14525929.CS1 maint: ref = harv (odkaz)
- ^ A b C d Firasat S, Khaliq S, Mohyuddin A, Papaioannou M, Tyler-Smith C, Underhill PA, Ayub Q (January 2007). „Y-chromozomální důkazy o omezeném řeckém příspěvku k Pathan populace v Pákistánu“. European Journal of Human Genetics. 15 (1): 121–6. doi:10.1038 / sj.ejhg.5201726. PMC 2588664. PMID 17047675.CS1 maint: ref = harv (odkaz)
- ^ Qamar 2002.
- ^ Mcelreavey 2005.
- ^ A b Kivisild T, Rootsi S, Metspalu M, Metspalu E, Parik J, Kaldma K, Usanga E, Mastana S, Papiha SS, Villems R (2003). "The Genetics of Language and Farming Spread in India" (PDF). In Bellwood P, Renfrew C (ed.). Zkoumání hypotézy šíření zemědělství / jazyka. McDonald Institute for Archaeological Research, Cambridge, United Kingdom. 215–222.
- ^ A b C Sengupta a kol. (2005)[úplná citace nutná ]
- ^ A b C d E Underhill PA, Myres NM, Rootsi S, Metspalu M, Zhivotovsky LA, King RJ a kol. (Duben 2010). „Oddělení post glaciálního spoluvlastnictví evropských a asijských chromozomů Y v haploskupině R1a“. European Journal of Human Genetics. 18 (4): 479–84. doi:10.1038 / ejhg.2009.194. PMC 2987245. PMID 19888303.CS1 maint: ref = harv (odkaz)
- ^ Mirabal S, Regueiro M, Cadenas AM, Cavalli-Sforza LL, Underhill PA, Verbenko DA, et al. (Říjen 2009). "Y-chromosome distribution within the geo-linguistic landscape of northwestern Russia". European Journal of Human Genetics. 17 (10): 1260–73. doi:10.1038 / ejhg.2009.6. PMC 2986641. PMID 19259129.
- ^ A b C Underhill 2014.
- ^ A b C Pamjav 2012.
- ^ ArunkumarG 2012.
- ^ Fornarino a kol. (2009)
- ^ Manoukian, Jean-Grégoire (2006), "A Synthesis of Haplogroup R2 – 2006 Archivováno 1. Října 2015 v Wayback Machine."
- ^ Mondal, Mayukh; Bergström, Anders; Xue, Yali; Calafell, Francesc; Laayouni, Hafid; Casals, Ferran; Majumder, Partha P .; Tyler-Smith, Chris; Bertranpetit, Jaume (1 May 2017). „Y-chromozomální sekvence různých indických populací a původ Andamanů“. Genetika člověka. 136 (5): 499–510. doi:10.1007 / s00439-017-1800-0. hdl:10230/34399. ISSN 1432-1203. PMID 28444560. S2CID 3725426.
- ^ A b Kumar V, Reddy AN, Babu JP, Rao TN, Langstieh BT, Thangaraj K, et al. (Březen 2007). „Důkazy chromozomu Y naznačují společné otcovské dědictví rakousko-asijských populací“. BMC Evoluční biologie. 7: 47. doi:10.1186/1471-2148-7-47. PMC 1851701. PMID 17389048.
- ^ Eaaswarkhanth M, Haque I, Ravesh Z, Romero IG, Meganathan PR, Dubey B, et al. (Březen 2010). „Stopy subsaharské a středovýchodní linie v indických muslimských populacích“. European Journal of Human Genetics. 18 (3): 354–63. doi:10.1038 / ejhg.2009.168. PMC 2859343. PMID 19809480.
- ^ "The Place of the Indian mtDNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World" (PDF). Archivováno (PDF) z původního dne 8. března 2012. Citováno 28. září 2011.
- ^ "Ethnologue report for Indo-European". Ethnologue.com. Archivováno z původního dne 15. října 2012. Citováno 24. listopadu 2015.
- ^ Baldi P (1990). Linguistic Change and Reconstruction Methodology. Walter de Gruyter. p. 342. ISBN 978-3-11-011908-4.
- ^ Burling (2003), str. 174–178.
- ^ Bradley (2012) notes, MK in the wider sense including the Munda languages of eastern South Asia is also known as Austroasiatic.Languages and Language Families in China Archivováno 30 April 2017 at the Wayback Machine
- ^ Bamshad M, Kivisild T, Watkins WS, Dixon ME, Ricker CE, Rao BB, et al. (Červen 2001). "Genetic evidence on the origins of Indian caste populations". Výzkum genomu. 11 (6): 994–1004. doi:10.1101/gr.GR-1733RR. PMC 311057. PMID 11381027.
- ^ Mukherjee N, Nebel A, Oppenheim A, Majumder PP (December 2001). "High-resolution analysis of Y-chromosomal polymorphisms reveals signatures of population movements from Central Asia and West Asia into India". Journal of Genetics. 80 (3): 125–35. doi:10.1007 / BF02717908. PMID 11988631. S2CID 13267463.
- ^ Mondal, Mayukh; Bergström, Anders; Xue, Yali; Calafell, Francesc; Laayouni, Hafid; Casals, Ferran; Majumder, Partha P .; Tyler-Smith, Chris; Bertranpetit, Jaume (25 April 2017). „Y-chromozomální sekvence různých indických populací a původ Andamanů“. Genetika člověka. 136 (5): 499–510. doi:10.1007 / s00439-017-1800-0. hdl:10230/34399. PMID 28444560. S2CID 3725426.
- ^ Reich D, Thangaraj K, Patterson N, Price AL, Singh L (September 2009). "Reconstructing Indian population history". Příroda. 461 (7263): 489–94. Bibcode:2009Natur.461..489R. doi:10.1038/nature08365. PMC 2842210. PMID 19779445.
- ^ Moorjani P, Thangaraj K, Patterson N, Lipson M, Loh PR, Govindaraj P, et al. (Září 2013). "Genetic evidence for recent population mixture in India". American Journal of Human Genetics. 93 (3): 422–38. doi:10.1016/j.ajhg.2013.07.006. PMC 3769933. PMID 23932107.
- ^ A b Chaubey, Gyaneshwer; Endicott, Phillip (June 2013). "The Andaman Islanders in a Regional Genetic Context: Reexamining the Evidence for an Early Peopling of the Archipelago from South Asia". Biologie člověka. 85 (1–3): 153–172. doi:10.3378/027.085.0307. PMID 24297224. S2CID 7774927.
- ^ Majumder PP (February 2010). "The human genetic history of South Asia". Aktuální biologie. 20 (4): R184-7. doi:10.1016/j.cub.2009.11.053. PMID 20178765. S2CID 1490419.
- ^ Chyba citace. Viz vložený komentář, jak opravit.[je nutné ověření ]
- ^ Pathak, Ajai K.; Kadian, Anurag; Kushniarevich, Alena; Montinaro, Francesco; Mondal, Mayukh; Ongaro, Linda; Singh, Manvendra; Kumar, Pramod; Rai, Niraj; Parik, Jüri; Metspalu, Ene; Rootsi, Siiri; Pagani, Luca; Kivisild, Toomas; Metspalu, Mait; Chaubey, Gyaneshwer; Villems, Richard (December 2018). "The Genetic Ancestry of Modern Indus Valley Populations from Northwest India". American Journal of Human Genetics. 103 (6): 918–929. doi:10.1016/j.ajhg.2018.10.022. PMC 6288199. PMID 30526867.
- ^ Reich, David (2018). Who We Are and How We Got Here: Ancient DNA and the new science of the human past. OUP Oxford. ISBN 978-0-19-257040-6. Citováno 2. března 2020.
- ^ A b C d Narasimhan a kol. 2018, str. 15.
- ^ Shinde V, Narasimhan VM, Rohland N, Mallick S, Mah M, Lipson M, Nakatsuka N, Adamski N, Broomandkhoshbacht N, Ferry M, Lawson AM, Michel M, Oppenheimer J, Stewardson K, Jadhav N, Kim YJ, Chatterjee M, Munshi A, Panyam A, Waghmare P, Yadav Y, Patel H, Kaushik A, Thangaraj K, Meyer M, Patterson N, Rai N, Reich D (September 2019). "An Ancient Harappan Genome Lacks Ancestry from Steppe Pastoralists or Iranian Farmers". Buňka. 179 (3): 729–735.e10. doi:10.1016/j.cell.2019.08.048. PMC 6800651. PMID 31495572.
- ^ Narasimhan VM, Patterson N, Moorjani P, Rohland N, Bernardos R, Mallick S, et al. (Září 2019). „Formování lidské populace v jižní a střední Asii“. Věda. 365 (6457): eaat7487. doi:10.1126 / science.aat7487. PMC 6822619. PMID 31488661.
- ^ Watkins WS, Prasad BV, Naidu JM, Rao BB, Bhanu BA, Ramachandran B, et al. (Listopad 2005). "Diversity and divergence among the tribal populations of India". Annals of Human Genetics. 69 (Pt 6): 680–92. doi:10.1046/j.1529-8817.2005.00200.x. PMID 16266407. S2CID 31907598.
- ^ Reddy BM, Naidu VM, Madhavi VK, Thangaraj LK, Kumar V, Langstieh BT, et al. (Prosinec 2005). "Microsatellite diversity in Andhra Pradesh, India: genetic stratification versus social stratification". Biologie člověka. 77 (6): 803–23. doi:10.1353/hub.2006.0018. PMID 16715839. S2CID 18446485.
- ^ Vishwanathan H, Deepa E, Cordaux R, Stoneking M, Usha Rani MV, Majumder PP (March 2004). "Genetic structure and affinities among tribal populations of southern India: a study of 24 autosomal DNA markers". Annals of Human Genetics. 68 (Pt 2): 128–38. doi:10.1046/j.1529-8817.2003.00083.x. PMID 15008792. S2CID 24230856.
- ^ "The Y-chromosome of the Soliga, an ancient forest-dwelling tribe of South India". Gene: X. 5: 100026. 1 December 2020. doi:10.1016/j.gene.2019.100026. ISSN 2590-1583.
Další čtení
- Allikas A, Ord D, Kurg R, Kivi S, Ustav M (June 2001). "Roles of the hinge region and the DNA binding domain of the bovine papillomavirus type 1 E2 protein in initiation of DNA replication". Virový výzkum. 75 (2): 95–106. doi:10.1016/S0168-1702(01)00219-2. PMID 11325464.CS1 maint: ref = harv (odkaz)
- Behar DM, Garrigan D, Kaplan ME, Mobasher Z, Rosengarten D, Karafet TM, et al. (Březen 2004). „Kontrastní vzorce variace chromozomu Y v aškenázských židovských a hostitelských nežidovských evropských populacích“. Genetika člověka. 114 (4): 354–65. doi:10.1007 / s00439-003-1073-7. PMID 14740294. S2CID 10310338.
- Bhattacharyya NP, Basu P, Das M, Pramanik S, Banerjee R, Roy B, et al. (Srpen 1999). "Negligible male gene flow across ethnic boundaries in India, revealed by analysis of Y-chromosomal DNA polymorphisms". Výzkum genomu. 9 (8): 711–9. doi:10.1101/gr.9.8.711 (inactive 25 September 2020). PMID 10447506.CS1 maint: DOI neaktivní od září 2020 (odkaz)
- Cann RL (March 2001). "Genetic clues to dispersal in human populations: retracing the past from the present". Věda. 291 (5509): 1742–8. Bibcode:2001Sci...291.1742C. doi:10.1126/science.1058948. PMID 11249820. S2CID 19367408.CS1 maint: ref = harv (odkaz)
- Cinnioğlu C, King R, Kivisild T, Kalfoğlu E, Atasoy S, Cavalleri GL a kol. (Leden 2004). "Vykopávání vrstev haplotypu Y-chromozomu v Anatolii". Genetika člověka. 114 (2): 127–48. doi:10.1007 / s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- Das B, Chauhan PS, Seshadri M (October 2004). "Minimal sharing of Y-chromosome STR haplotypes among five endogamous population groups from western and southwestern India". Biologie člověka. 76 (5): 743–63. doi:10.1353/hub.2005.0003. PMID 15757245. S2CID 12835244.
- Hemphill BE, Christensen AF (3 November 1994). The Oxus Civilization as a Link between East and West: A Non-Metric Analysis of Bronze Age Bactrain Biological Affinities. Madison, Wisconsin. p. 13.CS1 maint: ref = harv (odkaz) (paper read at the South Asia Conference)
- Jobling MA, Tyler-Smith C (August 2003). "The human Y chromosome: an evolutionary marker comes of age". Recenze přírody. Genetika. 4 (8): 598–612. doi:10.1038/nrg1124. PMID 12897772. S2CID 13508130.
- Kivisild T, Rootsi S, Metspalu M, Mastana S, Kaldma K, Parik J, et al. (Únor 2003). „Genetické dědictví prvních osadníků přetrvává jak v indických kmenových, tak v kastovních populacích“. American Journal of Human Genetics. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373.CS1 maint: ref = harv (odkaz)
- Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (Srpen 2004). „Většina existujících hranic mtDNA v jižní a jihozápadní Asii byla pravděpodobně formována během počátečního osídlení Eurasie anatomicky moderními lidmi“. Genetika BMC. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- Patowary A, Purkanti R, Singh M, Chauhan RK, Bhartiya D, Dwivedi OP, et al. (Červenec 2012). "Systematic analysis and functional annotation of variations in the genome of an Indian individual". Lidská mutace. 33 (7): 1133–40. doi:10.1002/humu.22091. PMID 22461382. S2CID 11466942.
- Rootsi S, Magri C, Kivisild T, Benuzzi G, Help H, Bermisheva M, et al. (Červenec 2004). "Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in europe". American Journal of Human Genetics. 75 (1): 128–37. doi:10.1086/422196. PMC 1181996. PMID 15162323.
- Qamar R, Ayub Q, Mohyuddin A, Helgason A, Mazhar K, Mansoor A, et al. (Květen 2002). "Y-chromosomal DNA variation in Pakistan". American Journal of Human Genetics. 70 (5): 1107–24. doi:10.1086/339929. PMC 447589. PMID 11898125.
- Semino O, Magri C, Benuzzi G, Lin AA, Al-Zahery N, Battaglia V, et al. (Květen 2004). „Původ, difúze a diferenciace haploskupin E a J chromozomu Y: závěry o neolitizaci Evropy a pozdějších migračních událostech ve středomořské oblasti“. American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
Zdroje
- Indian Genome Variation Consortium (April 2008). "Genetic landscape of the people of India: a canvas for disease gene exploration". Journal of Genetics. 87 (1): 3–20. doi:10.1007/s12041-008-0002-x. PMID 18560169. S2CID 21473349.
- Endicott P, Metspalu M, Kivisild T (2007). "Genetic evidence on modern human dispersals in South Asia: Y chromose and mitochondrial DNA perspectives". In Michael D. Petraglia, Bridget Allchin (eds.). Vývoj a historie lidských populací v jižní Asii. Springer. 201–228. ISBN 978-1-4020-5561-4.
- Hemphill B, Lukacs J, Kennedy K (1991). "Biological Adaptations and Affinities of Bronze Age Harappans". In Meadow RH (ed.). Harappa excavations 1986–1990: a multidisciplinary approach to third millennium urbanism. pp. 137–82. ISBN 978-0-9629110-1-9.CS1 maint: ref = harv (odkaz)
- Kennedy KA (1984). "A Reassessment of the Theories of Racial Origins of the People of the Indus Valley Civilization from Recent Anthropological Data". In Kennedy KA, Possehl GL (eds.). Studies in the Archaeology and Palaeoanthropology of South Asia. Atlantic Highlands, NJ: Humanities Press. pp. 99–107.CS1 maint: ref = harv (odkaz)
- Kennedy KA (1995). "Have Aryans been identified in the prehistoric skeletal record from South Asia?". In George Erdosy (ed.). The Indo-Aryans of Ancient South Asia. Walter de Gruyter. str. 49–54. ISBN 978-3-11-014447-5.
- Kivisild T (2000b). The origins of southern and western Eurasian populations: an mtDNA study (PDF). Tartu University, Estonia.CS1 maint: ref = harv (odkaz) (PhD)
- Mascarenhas DD, Raina A, Aston CE, Sanghera DK (2015). "Genetic and Cultural Reconstruction of the Migration of an Ancient Lineage". BioMed Research International. 2015: 651415. doi:10.1155/2015/651415. PMC 4605215. PMID 26491681.
- Narasimhan VM, Anthony D, Mallory J, Reich D (2018). „Genomická formace jižní a střední Asie“. bioRxiv 10.1101/292581. doi:10.1101/292581. S2CID 89658279. Citovat deník vyžaduje
| deník =(Pomoc) - Oppenheimer S (2003). The Real Eve: Modern Man's Journey out of Africa. New York: Carroll a Graf Publishers. ISBN 978-0-7867-1192-5.CS1 maint: ref = harv (odkaz)
- Pamjav (December 2012), "Brief communication: New Y-chromosome binary markers improve phylogenetic resolution within haplogroup R1a1", American Journal of Physical Anthropology, 149 (4): 611–615, doi:10.1002 / ajpa.22167, PMID 23115110, S2CID 4820868
- Renfrew C, Boyle K, eds. (2000a). An Indian Ancestry: a Key for Understanding Human Diversity in Europe and Beyond (PDF). ISBN 978-1-902937-08-3.CS1 maint: ref = harv (odkaz)
- Underhill PA (2003). "Inferring human history: clues from Y-chromosome haplotypes". Cold Spring Harbor Symposia o kvantitativní biologii. 68: 487–93. doi:10.1101/sqb.2003.68.487. PMID 15338652. S2CID 15527457.
- Underhill PA, Poznik GD, Rootsi S, Järve M, Lin AA, Wang J, et al. (Leden 2015). "The phylogenetic and geographic structure of Y-chromosome haplogroup R1a". European Journal of Human Genetics. 23 (1): 124–31. doi:10.1038 / ejhg.2014.50. PMC 4266736. PMID 24667786.
- Wells S (2003). The Journey of Man: A Genetic Odyssey. Princeton University Press.
externí odkazy
- Introduction to Haplogroups and Haplotypes, Mark A. Jobling, University of Leicester. [1]
- Journey of Man: Peopling of the World, Bradshaw Foundation, in association with Stephen Oppenheimer.
- Indian Genome Variation Database Ústav genomiky a integrační biologie
- Seznam frekvence R2
- Maledivy
| Podtémata | |
|---|---|
| Genetická historie podle regionu | |
| Populační genetika podle skupiny | |
| |