STIM2 - STIM2




Molekula stromální interakce 2 (STIM2) je protein že u lidí je kódován STIM2 gen.[5][6]
Tento gen je členem rodiny stromálních interakčních molekul (STIM), která obsahuje pouze dva členy spolu s homologem STIM1 a pravděpodobně vznikl ze společného genu předků. Zakódují typ 1 transmembránové proteiny které se nacházejí v sarko / endoplazmatickém retikulu (SR / ER ) do buňky. Alternativní iniciace překladu z počátečního místa AUG a non-AUG (UUG) vede k produkci dvou různých STIM2 izoformy.
Oba členové rodiny STIM byli v roce 2005 identifikováni jakovápník (Ca2+) senzory, které se účastní mechanismu Ca2+ vstup do buňky označované jako provozováno v obchodě Ca2+ vstup (SOCE). Mnoho buněčných procesů a signálních drah je spuštěno předchozím uvolněním Ca2+ uloženo v subcelulární organely, který potřebuje průběžné doplňování. SOCE je považován za mechanismus doplňování zásob a základní mechanismus Ca2+ signalizace v neelektricky excitovatelných buňkách. Zatímco se aktivuje STIM1 SOCE, výzkum funkce STIM2 naznačuje hlavní roli jako regulátor zpětné vazby, který stabilizuje bazální cytosolický a S / ER Ca2+ koncentrace [Ca2+]. STIM2 detekuje malé poklesy Ca2+ obsah uložený v S / ER, přepne do aktivovaného stavu a interaguje s tzv provozováno v obchodě Ca2+ (SOC) kanály nachází se v plazmatická membrána, jako Orai nebo TRPC kanály, umožňující SOCE. Ačkoli funkční role STIM2 je po mnoho let nepolapitelná, studie provedené v letech 2009–2010 na myších modelech naznačují, že se STIM2 účastní procesů vývoje a fungování mnoha typů buněk, včetně hladkého svalstva myoblasty buňky buňky imunitní systém a neurony, a je zapojen do tumorigeneze, vývoj autoimunitní onemocnění a mechanismy poškození neuronů po přechodném stavu ischemická podmínky.
Gen
V roce 2001 STIM2 byl identifikován jako nový lidský homolog STIM1 gen, představující druhého člena rodiny dvou genů u obratlovců.[5] The STIM2 gen obsahuje 12 exony a 11 introny umístěný na lidském chromozomu 4p15.1 a na velkém rameni myšího chromozomu 5, blízko k centroméra. Členové STIM rodina se nejpravděpodobněji vyvinula z jediného genu v nižší mnohobuněčné eukaryoty do dvou příbuzných genů u obratlovců, od člověka STIM1 a STIM2 stejně jako Drosophila melanogaster Stim (D-Stim) mají konzervovanou genomickou organizaci. D-STIM protein 570 aas vykazuje stejnou podobnost s oběma STIM1 (33% identické; 50% aminokyselina sekvence konzervována) a STIM2 (31% identické; 46% aminokyselinové sekvence konzervováno). Jednobuněčný eukaryoty jako Monosiga brevicollis jednobuněčný choanoflagellate bylo hlášeno, že má gen podobný STIM,[7] nicméně ne STIM- podobné geny byly identifikovány v prokaryoty. U obratlovců nebyly dosud identifikovány žádné další proteiny podobné STIM.[5]
Struktura bílkovin
Protein STIM2 je transmembránový protein typu I nacházející se v S / ER. Lidský STIM2 se skládá z 833 aminokyselinových zbytků (aas ) (105-115 kDa) (obr. 1), 148 dalších aas ve srovnání s lidským STIM1. Jejich N-terminál regiony mají 66% podobnost s 577 aas (85% aminokyselinové sekvence STIM1). Pouze extrém C-terminál oblast vykazuje významnou divergenci sekvence. Architektura domény obou izoforem je u obratlovců vysoce zachována (obr. 1). Myší STIM2 sdílí 92% identitu s lidským STIM2 v sekvenci aminokyselin podle párového zarovnání generovaného VÝBUCH. Jejich doména struktura je také vysoce konzervovaná (obr. 1). Lidský STIM2 je posttranslačně upraven in vivo, jako je zrání štěpením N-koncového S / ER signálního peptidu (14 aas), glykosylace a různé stupně fosforylace, ale fosforylovaná místa jsou stále neznámá (obr. 1).[5]
Architektura domény
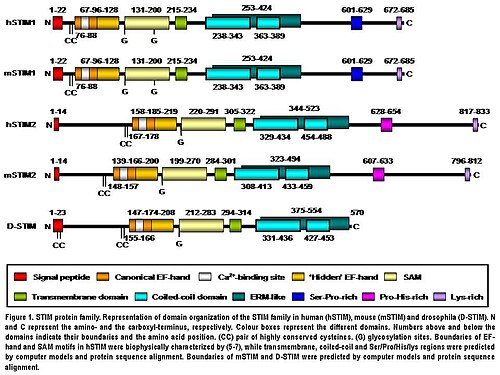
N-koncová oblast STIM2 je umístěna v S / ER lumen a obsahuje kanonický EF ruka Ca.2+-vazba motiv „skrytý“ EF-ruka Ca2+nedávno objevený vazebný motiv a sterilní doména a-motivu (SAM), známý motiv interakce protein-protein (obr. 1).[8][9][10]N-koncová část je oddělena od C-koncové oblasti jedním průchodem transmembránový motiv který je vysoce konzervovaný ve všech proteinech STIM. C-koncová oblast obsahuje vysoký stupeň α-šroubovice struktur. Velká část blízká transmembránové doméně zahrnuje oblast podobnou ezrinu / radixinu / moesinu (ERM ) doména, která obsahuje dvě svinutá cívka domén.[11] Domény coiled-coil zprostředkovávají interakce mezi proteiny STIM, což jim umožňuje navzájem se vázat a vytvářet homo a heterodimery (obr. 1).[12][13][14] Nakonec dále směrem k C-konci obsahuje STIM2 motiv bohatý na prolin / histidin a ocas bohatý na lysin 17 aas (obr. 1).[5]
EF-ruka-SAM oblast
Protože domény EF-ruka a SAM (EF-SAM) jsou životně důležité pro funkci STIM a regulaci SOCE, jsou nyní podrobně diskutovány. EF doménou je Ca2+ senzor používaný proteinem STIM k detekci změn Ca2+ koncentrace uvnitř S / ER. Izoformy STIM se aktivují, když Ca2+ vázaný na motiv EF ruky se uvolní v důsledku snížení Ca2+ úrovně uvnitř obchodu S / ER po IP3 receptor - zprostředkované vyčerpání. Bylo hlášeno, že STIM EF-ruční mutanti, kteří nejsou schopni vázat Ca2+ jsou konstitutivně aktivní a neustále aktivují SOCE nezávisle na S / ER [Ca2+], in vitro[15] a in vivo.[16][17][18]Doména SAM je důležitá pro oligomerizaci STIM, protože mutantům v této doméně chybí schopnost vytvářet indukovatelné punctae.[19]Ca.2+-vázací experimenty in vitro s použitím lidských fragmentů STIM1 EF – SAM (zbytek 58–201) nebo STIM2 EF – SAM (zbytek 149–292) ukazuje, že obě izoformy vážou Ca2+ s podobnou afinitou (STIM2 Kd ~ 0,5 mM; STIM1 Kd ~ 0,2–0,6 mM),[20][21] což je v rozsahu hodnot hlášených pro S / ER [Ca2+].[22][23] STIM2 se však liší od STIM1 v tom, že je již částečně aktivní při bazálním S / ER [Ca2+] a plně se aktivuje dříve během vyčerpání úložiště S / ER. Přes stejný Ca2+ afinita vykazovaná fragmenty STIM EF-SAM, plný protein STIM2 vykazoval nižší [Ca2+] citlivost než STIM1 v transfekovaných buňkách in vitro.[24] Tento nesoulad naznačuje, že další proteinové oblasti navíc přispívají k různým [Ca2+] prahová hodnota citlivosti nebo aktivace zobrazená oběma izoformami. „Skrytá“ doména EF-hand neváže Ca2+, ale je to zásadní pro intramolekulární asociaci, skládání a stabilitu domén EF-ruka a SAM. Velmi nedávno bylo oznámeno, že strukturálně kritické mutace v kanonické ruce EF, „skryté“ ruce EF nebo SAM naruší Ca2+ citlivost v důsledku destabilizace celé oblasti EF-SAM.[25]
C-koncová oblast
Kromě N-konce je C-koncová oblast také podstatnou součástí proteinů STIM. Ukazuje významnou divergenci sekvence mezi oběma izoformami a v STIM1 je C-koncová oblast nezbytná pro interakci s SOC kanály.[26] Human STIM2 obsahuje a prolin - a histidin -rich motiv (PHAPHPSHPRHPHHPQHTPHSLPSPDP) na podobné pozici jako a serin - a prolin -rich region (SPSAPPGGSPHLDSSRSHSPSSPDPDTPSP) v STIM1. Významná divergence v těchto oblastech by mohla naznačovat divergenci ve funkci izoforem STIM. Na rozdíl od STIM1 má STIM2 dilysin ER retenční signál (K (X) KXX) na svém extrémním C-konci, který zadržuje protein v ER, zatímco STIM1 může cestovat na povrch buňky.[27] Konečně podobné lysin - bohaté konce 14 a 17 zbytků v STIM1 a STIM2 jsou umístěny na samém konci C-terminální oblasti. lineární peptidy z C-terminální polybazické oblasti lidského STIM1 (zbytky 667-685) a STIM2 (zbytky 730-746) se váží na klimodulin s vysokou nebo nízkou afinitou v přítomnosti nebo nepřítomnosti Ca2+, resp.[28] Většina studií interakcí C-terminální oblasti byla provedena s izoformou STIM1. Přidání thapsigargin (dále jen SERCA inhibitor pumpy, který stimuluje SOCE pasivním vyčerpáním intracelulárního Ca2+ zásoby) do buněk lidské slinné žlázy a také na rozptýlenou myš submandibulární žláza buněk přibývá koimunoprecipitace TRPC1 a Orai1 se STIM1.[29]Podle in vitro koexprese různých lidských mutantů STIM1, kterým chybí různé C-koncové oblasti v buňkách HEK293, tři nedávné práce uvádějí, že doména ERM na C-konci (aas 251-535, obr. 1), obsahující svinutá cívka domén, zprostředkovává vazbu STIM1 na TRPC (1, 2,4 a 5) a migraci STIM1 na plazmatickou membránu. Kromě toho je pro klonování TRPC1 nezbytná oblast bohatá na kationtové lysiny.[14][26][30] Li a kol. dále vymezil tyto oblasti (aas 425-672) jako možná místa interakce STIM1-Orai1.[14] In vitro koimunoprecipitace experimenty po přechodné koexpresi proteinů STIM2 a Orai1 v HEK293 buňky odhalily, že také STIM2 může fyzicky interagovat s Orai1, pravděpodobně přes C-koncovou oblast STIM2.[31]
Exprese a distribuce tkáně
STIM2 mRNA je vyjádřen většinou lidských tkání. Protein STIM2 je exprimován mnoha lidmi buněčné linie společně se STIM1, což naznačuje, že izoformy STIM jsou koexprimovány ve stejné buňce, alespoň v zavedených buněčných liniích.[5] Protein STIM2 je široce exprimován ve tkáních, obvykle přítomných v nižších hladinách než STIM1, s výjimkou mozku nebo jater, pokud se STIM2 jeví jako dominantní izoforma.[5][32] Transkripce Stim2 je také dynamicky regulovaná, například je upregulována při diferenciaci naivního T buňky do Čt1 nebo Čt2 lymfocyty.[33]
Funkce
Funkce STIM2 byla kontroverzní. Počáteční studie to zjistily siRNA srazit STIM1, ale ne STIM2, silně snížil SOCE v buňkách savců.[15][24][34][35][36] Liou et al. uvádějí mírné snížení SOCE také snížením STIM2 v buňkách HeLa.[15] Soboloff et al. navrhl, že STIM2 inhibuje SOCE, když je exprimován samostatně,[13] ale společně s Orai1 způsobuje podstatnou konstitutivní SOCE.[37] Naproti tomu Brandman et al. navrhl, že STIM2 by mohl působit jako regulátor, který stabilizuje bazální cytosolický a ER Ca2+ úrovně.[24] Parvez et al., použitím in vitro přechodná koexprese lidského STIM2 a různých SOC kanály v HEK293 buňky uvádí, že STIM2 zprostředkovává SOCE prostřednictvím dvou režimů závislých na obchodu a nezávislých na obchodu.[31] Společně tyto výsledky naznačují komplexní interakci jemně regulovanou buněčným poměrem STIM1: STIM2: Orai a jejich endogenními hladinami.
Studie provedené v letech 2009–2010 s použitím člověka in vitro nebo myší in vivo modely potvrdily Brandmana et al. výsledky a naznačil, že STIM2 se účastní procesů vývoje a fungování mnoha typů buněk, včetně hladkého svalstva myoblasty, buňky imunitního systému a neurony. Kromě toho se podílí na tumorigenezi, vývoji autoimunitních onemocnění a mechanismech poškození neuronů po přechodných ischemických stavech. V klidových podmínkách mají kultivované buňky HEK293 nadměrně exprimující nebo kortikální neurony bez STIM2 zvýšené nebo snížené klidové intracelulární Ca2+ úrovně,[31][32] podpora myšlenky, že STIM2 je nezbytný pro regulaci intracelulárního bazálního Ca2+ úrovně. Buňky jsou však velmi aktivní in vivo a intracelulární Ca2+ úrovně neustále kolísají. Vývoj nových metod ke studiu in vivo role STIM2 v intracelulárním Ca2+ úrovně by byly nutné. V kultivovaném lidském myoblastu se STIM2 účastní buněčné diferenciace na myotubes.[38] V imunitním systému se účastní STIM2 T buňka aktivací vyvolaná produkce interleukin2 (IL-2) a interferon gama (IFNy), pravděpodobně stabilizací NFAT bydliště v jádro, stejně jako v diferenciaci naivních T buňky do Čt17 lymfocyty, které jsou pravděpodobně důležité v časných fázích autoimunitních onemocnění.[33][39] Ve skutečnosti vykazovaly myši s deficitem STIM2 mírnou symptomatologii v rané fázi autoimunitních onemocnění.[39] V neuronální tkáni hraje STIM2 klíčovou roli ischemií indukované poškození neuronů a absence STIM2 ve Windows knockout myši snížilo neuronální poškození způsobené ischemií po přechodném přerušení průtoku krve v mozku.[32] Tento neuroprotektivní účinek deficitu STIM2 po ischemické epizodě naznačuje, že inhibitory funkce STIM2 mohou mít tedy potenciální terapeutickou hodnotu jako neuroprotektivní látky k léčbě ischemické poranění a další neurodegenerativní poruchy zahrnující změněnou Ca2+ homeostáza. Stejná vědecká studie navíc navrhla důležitou roli STIM2 v hipokampus -závislý prostorová paměť, synaptický přenos a plasticita.[32]
Nakonec byla pro STIM2 spolu s STIM1 v roce prokázána onkogenní funkce glioblastoma multiforme, kde oba proteiny mají zvýšenou expresi a / nebo zvýšený počet kopií.[40][41] Navíc se STIM2 nachází v chromozomu 4p15.1, oblasti zapojené do invazivních karcinomů plic, prsu, krku a hlavy.[42][43][44]
Interakce
Jak již bylo zmíněno, bylo prokázáno, že STIM2 interaguje s STIM1,[12][13][14] SOC kanály jako Orai (ICRACM) nebo TRPC,[31] kalmodulin (CaM)[28][31] a také plazmatická membrána fosfoinositidy.[45] Ukázalo se, že exprese STIM2 je ovlivňována nebo regulována preseniliny v myších embryonálních fibroblastech a lidských B lymfocytech.[46]
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000109689 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000039156 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b C d E F G Williams RT, Manji SS, Parker NJ, Hancock MS, Van Stekelenburg L, Eid JP, Senior PV, Kazenwadel JS, Shandala T, Saint R, Smith PJ, Dziadek MA (srpen 2001). „Identifikace a charakterizace rodiny genů STIM (molekula stromální interakce): kódování nové třídy transmembránových proteinů“. The Biochemical Journal. 357 (Pt 3): 673–85. doi:10.1042/0264-6021:3570673. PMC 1221997. PMID 11463338.
- ^ "Entrez Gene: STIM2 stromální interakce molekula 2".
- ^ Cai X (červenec 2008). „Jednobuněčný Ca2 + signalizující„ sada nástrojů “na počátku metazoa“. Molekulární biologie a evoluce. 25 (7): 1357–61. doi:10.1093 / molbev / msn077. PMID 18385221.
- ^ Kim CA, Bowie JU (prosinec 2003). "SAM domény: jednotná struktura, různorodost funkcí". Trendy v biochemických vědách. 28 (12): 625–8. doi:10.1016 / j.tibs.2003.11.001. PMID 14659692.
- ^ Qiao F, Bowie JU (květen 2005). "Mnoho tváří SAMa". Věda STKE. 2005 (286): re7. doi:10.1126 / stke.2862005re7. PMID 15928333. S2CID 31699798.
- ^ Schultz J, Ponting CP, Hofmann K, Bork P (leden 1997). „SAM jako doména interakce proteinů zapojená do vývojové regulace“. Věda o bílkovinách. 6 (1): 249–53. doi:10.1002 / pro.5560060128. PMC 2143507. PMID 9007998.
- ^ Parry DA, Fraser RD, Squire JM (září 2008). „Padesát let svinutých cívek a alfa-šroubovicových svazků: úzký vztah mezi posloupností a strukturou“. Journal of Structural Biology. 163 (3): 258–69. doi:10.1016 / j.jsb.2008.01.016. PMID 18342539.
- ^ A b Williams RT, Senior PV, Van Stekelenburg L, Layton JE, Smith PJ, Dziadek MA (duben 2002). „Stromální interakční molekula 1 (STIM1), transmembránový protein s potlačující růstovou aktivitou, obsahuje extracelulární doménu SAM modifikovanou N-vázanou glykosylací“. Biochimica et Biophysica Acta (BBA) - proteinová struktura a molekulární enzymologie. 1596 (1): 131–7. doi:10.1016 / S0167-4838 (02) 00211-X. PMID 11983428.
- ^ A b C Soboloff J, Spassova MA, Hewavitharana T, He LP, Xu W, Johnstone LS, Dziadek MA, Gill DL (červenec 2006). "STIM2 je inhibitor STIM1 zprostředkovaného obchodu Ca2 + Entry". Aktuální biologie. 16 (14): 1465–70. doi:10.1016 / j.cub.2006.05.051. PMID 16860747. S2CID 16570108.
- ^ A b C d Li Z, Lu J, Xu P, Xie X, Chen L, Xu T (říjen 2007). "Mapování interagujících domén STIM1 a Orai1 v aktivaci Ca2 + kanálu Ca2 + uvolňováním". The Journal of Biological Chemistry. 282 (40): 29448–56. doi:10,1074 / jbc.M703573200. PMID 17702753.
- ^ A b C Liou J, Kim ML, Heo WD, Jones JT, Myers JW, Ferrell JE, Meyer T (červenec 2005). „STIM je senzor Ca2 + nezbytný pro příliv Ca2 + vyvolaný vyčerpáním skladu Ca2 +“. Aktuální biologie. 15 (13): 1235–41. doi:10.1016 / j.cub.2005.05.055. PMC 3186072. PMID 16005298.
- ^ Grosse J, Braun A, Varga-Szabo D, Beyersdorf N, Schneider B, Zeitlmann L, Hanke P, Schropp P, Mühlstedt S, Zorn C, Huber M, Schmittwolf C, Jagla W, Yu P, Kerkau T, Schulze H, Nehls M, Nieswandt B (listopad 2007). „EF ruka mutace v Stim1 způsobuje předčasnou aktivaci krevních destiček a krvácení u myší“. The Journal of Clinical Investigation. 117 (11): 3540–50. doi:10.1172 / JCI32312. PMC 2040319. PMID 17965774.
- ^ Mercer JC, Dehaven WI, Smyth JT, Wedel B, Boyles RR, Bird GS, Putney JW (srpen 2006). „Velké skladovací vápníkové selektivní proudy v důsledku společné exprese Orai1 nebo Orai2 s intracelulárním vápníkovým senzorem, Stim1“. The Journal of Biological Chemistry. 281 (34): 24979–90. doi:10,1074 / jbc.M604589200. PMC 1633822. PMID 16807233.
- ^ Zhang SL, Yeromin AV, Zhang XH, Yu Y, Safrina O, Penna A, Roos J, Stauderman KA, MD Cahalan (červen 2006). „Screening příjmu Ca (2+) v celém genomu RNAi identifikuje geny, které regulují aktivitu Ca (2+) aktivovaného kanálu Ca (2+)“. Sborník Národní akademie věd Spojených států amerických. 103 (24): 9357–62. doi:10.1073 / pnas.0603161103. PMC 1482614. PMID 16751269.
- ^ Baba Y, Hayashi K, Fujii Y, Mizushima A, Watarai H, Wakamori M, Numaga T, Mori Y, Iino M, Hikida M, Kurosaki T (listopad 2006). „Spojení STIM1 se vstupem Ca2 + provozovaným v obchodě prostřednictvím jeho konstitutivního a indukovatelného pohybu v endoplazmatickém retikulu“. Sborník Národní akademie věd Spojených států amerických. 103 (45): 16704–9. doi:10.1073 / pnas.0608358103. PMC 1636519. PMID 17075073.
- ^ Zheng L, Stathopulos PB, Li GY, Ikura M (duben 2008). "Biofyzikální charakterizace EF-ruky a SAM domény obsahující Ca2 + senzorickou oblast STIM1 a STIM2". Sdělení o biochemickém a biofyzikálním výzkumu. 369 (1): 240–6. doi:10.1016 / j.bbrc.2007.12.129. PMID 18166150.
- ^ Stathopulos PB, Li GY, Plevin MJ, Ames JB, Ikura M (listopad 2006). „Uložená oligomerizace stromální interakce molekuly 1 (STIM1) vyvolaná vyčerpáním Ca2 + prostřednictvím oblasti EF-SAM: Mechanismus iniciace pro kapacitní vstup Ca2 +“. The Journal of Biological Chemistry. 281 (47): 35855–62. doi:10,1074 / jbc.M608247200. PMID 17020874.
- ^ Demaurex N, Frieden M (srpen 2003). "Měření koncentrace volného luminálního ER Ca (2+) s cíleným" cameleon "fluorescenčními proteiny". Buněčný vápník. 34 (2): 109–19. doi:10.1016 / S0143-4160 (03) 00081-2. PMID 12810053.
- ^ Barrero MJ, Montero M, Alvarez J (říjen 1997). "Dynamika [Ca2 +] v endoplazmatickém retikulu a cytoplazmě intaktních buněk HeLa. Srovnávací studie". The Journal of Biological Chemistry. 272 (44): 27694–9. doi:10.1074 / jbc.272.44.27694. PMID 9346910.
- ^ A b C Brandman O, Liou J, Park WS, Meyer T (prosinec 2007). „STIM2 je regulátor zpětné vazby, který stabilizuje hladiny Ca2 + v bazálním cytosolickém a endoplazmatickém retikulu“. Buňka. 131 (7): 1327–39. doi:10.1016 / j.cell.2007.11.039. PMC 2680164. PMID 18160041.
- ^ Stathopulos PB, Zheng L, Li GY, Plevin MJ, Ikura M (říjen 2008). "Strukturální a mechanické pohledy na zahájení vstupu vápníku provozovaného v obchodě pomocí STIM1". Buňka. 135 (1): 110–22. doi:10.1016 / j.cell.2008.08.006. PMID 18854159. S2CID 15447873.
- ^ A b Huang GN, Zeng W, Kim JY, Yuan JP, Han L, Muallem S, Worley PF (září 2006). "STIM1 karboxylový konec aktivuje nativní kanály SOC, I (crac) a TRPC1". Přírodní buněčná biologie. 8 (9): 1003–10. doi:10.1038 / ncb1454. PMID 16906149. S2CID 22135544.
- ^ Ercan E, Chung SH, Bhardwaj R, Seedorf M (červenec 2012). „Di-argininové signály a doména bohatá na K zadržují Ca2⁺ senzor STIM1 v endoplazmatickém retikulu.“ Provoz. 13 (7): 992–1003. doi:10.1111 / j.1600-0854.2012.01359.x. PMID 22498042. S2CID 33126543.
- ^ A b Bauer MC, O'Connell D, Cahill DJ, Linse S (červen 2008). "Vazba kalmodulinu na polybazické C-konce STIM proteinů zapojených do vstupu vápníku provozovaného v obchodě". Biochemie. 47 (23): 6089–91. doi:10.1021 / bi800496a. PMID 18484746.
- ^ Ong HL, Cheng KT, Liu X, Bandyopadhyay BC, Paria BC, Soboloff J, Pani B, Gwack Y, Srikanth S, Singh BB, Gill DL, Gill D, Ambudkar IS (březen 2007). „Dynamické sestavování ternárního komplexu TRPC1-STIM1-Orai1 se podílí na přílivu vápníku provozovaného v obchodě. Důkazy o podobnostech ve složkách vápníkového kanálu aktivovaného v obchodě a uvolňování vápníku. The Journal of Biological Chemistry. 282 (12): 9105–16. doi:10,1074 / jbc.M608942200. PMC 3309402. PMID 17224452.
- ^ Yuan JP, Zeng W, Huang GN, Worley PF, Muallem S (červen 2007). „STIM1 heteromultimerizuje kanály TRPC, aby určil jejich funkci jako kanálů provozovaných v obchodě“. Přírodní buněčná biologie. 9 (6): 636–45. doi:10.1038 / ncb1590. PMC 2699187. PMID 17486119.
- ^ A b C d E Parvez S, Beck A, Peinelt C, Soboloff J, Lis A, Monteilh-Zoller M, Gill DL, Fleig A, Penner R (březen 2008). „Protein STIM2 zprostředkovává odlišné režimy aktivace kanálu CRAC závislé na obchodu a nezávislé na obchodu“. FASEB Journal. 22 (3): 752–61. doi:10.1096 / fj.07-9449com. PMC 3601890. PMID 17905723.
- ^ A b C d Berna-Erro A, Braun A, Kraft R, Kleinschnitz C, Schuhmann MK, Stegner D, Wultsch T, Eilers J, Meuth SG, Stoll G, Nieswandt B (2009). „STIM2 reguluje kapacitní vstup Ca2 + v neuronech a hraje klíčovou roli při hypoxické smrti neuronových buněk“. Vědecká signalizace. 2 (93): ra67. doi:10.1126 / scisignal.2000522. PMID 19843959.
- ^ A b Oh-Hora M, Yamashita M, Hogan PG, Sharma S, Lamperti E, Chung W, Prakriya M, Feske S, Rao A (duben 2008). „Duální funkce pro senzory vápníku endoplazmatického retikula STIM1 a STIM2 v aktivaci a toleranci T buněk“. Přírodní imunologie. 9 (4): 432–43. doi:10.1038 / ni1574. PMC 2737533. PMID 18327260.
- ^ Roos J, DiGregorio PJ, Yeromin AV, Ohlsen K, Lioudyno M, Zhang S, Safrina O, Kozak JA, Wagner SL, Cahalan MD, Veliçelebi G, Stauderman KA (květen 2005). „STIM1, základní a konzervovaná součást funkce kanálu Ca2 + provozovaná v obchodě“. The Journal of Cell Biology. 169 (3): 435–45. doi:10.1083 / jcb.200502019. PMC 2171946. PMID 15866891.
- ^ Zhang SL, Yu Y, Roos J, Kozak JA, Deerinck TJ, Ellisman MH, Stauderman KA, Cahalan MD (říjen 2005). „STIM1 je senzor Ca2 +, který aktivuje kanály CRAC a migruje z úložiště Ca2 + do plazmatické membrány“. Příroda. 437 (7060): 902–5. doi:10.1038 / příroda04147. PMC 1618826. PMID 16208375.
- ^ Spassova MA, Soboloff J, He LP, Xu W, Dziadek MA, Gill DL (březen 2006). „STIM1 má roli plazmatické membrány v aktivaci Ca (2+) kanálů provozovaných v obchodě“. Sborník Národní akademie věd Spojených států amerických. 103 (11): 4040–5. doi:10.1073 / pnas.0510050103. PMC 1449642. PMID 16537481.
- ^ Soboloff J, Spassova MA, Tang XD, Hewavitharana T, Xu W, Gill DL (červenec 2006). „Orai1 a STIM rekonstituují funkci kalciového kanálu provozovaného v obchodě“. The Journal of Biological Chemistry. 281 (30): 20661–5. doi:10,1074 / jbc.C600126200. PMID 16766533.
- ^ Darbellay B, Arnaudeau S, Ceroni D, Bader CR, Konig S, Bernheim L (červenec 2010). „Diferenciace myoblastů lidské ekonomiky a spojování excitace a kontrakce používají stejné molekulární partnery, STIM1 a STIM2“. The Journal of Biological Chemistry. 285 (29): 22437–47. doi:10.1074 / jbc.M110.118984. PMC 2903423. PMID 20436167.
- ^ A b Schuhmann MK, Stegner D, Berna-Erro A, Bittner S, Braun A, Kleinschnitz C, Stoll G, Wiendl H, Meuth SG, Nieswandt B (únor 2010). „Stromální interakční molekuly 1 a 2 jsou klíčovými regulátory autoreaktivní aktivace T buněk při zánětu myšího autoimunitního centrálního nervového systému“. Journal of Immunology. 184 (3): 1536–42. doi:10,4049 / jimmunol.0902161. PMID 20028655.
- ^ Scrideli CA, Carlotti CG, Okamoto OK, Andrade VS, Cortez MA, Motta FJ, Lucio-Eterovic AK, Neder L, Rosemberg S, Oba-Shinjo SM, Marie SK, Tone LG (červenec 2008). „Analýza profilu genové exprese primárních glioblastomů a nenádorové mozkové tkáně: identifikace potenciálních cílových genů pomocí oligonukleotidové microarray a kvantitativní PCR v reálném čase“. Journal of Neuro-Oncology. 88 (3): 281–91. doi:10.1007 / s11060-008-9579-4. PMID 18398573. S2CID 31862636.
- ^ Ruano Y, Mollejo M, Ribalta T, Fiaño C, Camacho FI, Gómez E, de Lope AR, Hernández-Moneo JL, Martínez P, Meléndez B (2006). "Identifikace nových kandidátských cílových genů v amplikonech nádorů Glioblastoma multiforme detekovaných expresí a profilováním CGH microarray". Molekulární rakovina. 5: 39. doi:10.1186/1476-4598-5-39. PMC 1592108. PMID 17002787.
- ^ Pershouse MA, Ligon AH, Pereira-Smith OM, Killary AM, Yung WK, Steck PA (listopad 1997). „Potlačení transformovaného fenotypu a tumorigenicity po přenosu chromozomu 4 do lidských gliomových buněk U251“. Geny, chromozomy a rakovina. 20 (3): 260–7. doi:10.1002 / (SICI) 1098-2264 (199711) 20: 3 <260 :: AID-GCC6> 3.0.CO; 2-0. PMID 9365833.
- ^ Richard F, Pacyna-Gengelbach M, Schlüns K, Fleige B, Winzer KJ, Szymas J, Dietel M, Petersen I, Schwendel A (květen 2000). "Vzory chromozomální nerovnováhy u invazivního karcinomu prsu". International Journal of Cancer. 89 (3): 305–10. doi:10.1002 / 1097-0215 (20000520) 89: 3 <305 :: AID-IJC15> 3.0.CO; 2-8. PMID 10861509.
- ^ Petersen S, Aninat-Meyer M, Schlüns K, Gellert K, Dietel M, Petersen I (leden 2000). „Chromozomální změny v klonální evoluci do metastatického stadia spinocelulárních karcinomů plic“. British Journal of Cancer. 82 (1): 65–73. doi:10.1054 / bjoc.1999.0878. PMC 2363206. PMID 10638968.
- ^ Ercan E, Momburg F, Engel U, Temmerman K, Nickel W, Seedorf M (prosinec 2009). „Konzervovaný lipidem zprostředkovaný třídící mechanismus kvasnicových Ist2 a savčích proteinů STIM do periferního ER“. Provoz. 10 (12): 1802–18. doi:10.1111 / j.1600-0854.2009.00995.x. PMID 19845919. S2CID 31387679.
- ^ Bojarski L, Pomorski P, Szybinska A, Drab M, Skibinska-Kijek A, Gruszczynska-Biegala J, Kuznicki J (červen 2009). „Exprese proteinů STIM závislá na presenilinu a dysregulace kapacitního vstupu Ca2 + u familiární Alzheimerovy choroby“. Biochimica et Biophysica Acta (BBA) - výzkum molekulárních buněk. 1793 (6): 1050–7. doi:10.1016 / j.bbamcr.2008.11.008. PMID 19111578.