Drobný histokompatibilní antigen - Minor histocompatibility antigen
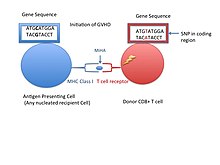
Drobný histokompatibilní antigen (také známý jako MiHA) jsou receptory na buněčném povrchu darovaných orgánů, o nichž je známo, že při některých orgánových transplantacích poskytují imunologickou odpověď.[1] Způsobují problémy s odmítnutím méně často než problémy s odmítnutím hlavní komplex histokompatibility (MHC). Drobné histokompatibilní antigeny (MiHA) jsou rozmanité, krátké segmenty bílkoviny a jsou označovány jako peptidy. Tyto peptidy mají obvykle délku přibližně 9-12 aminokyselin a jsou vázány jak na hlavní histokompatibilní komplex (MHC) třída Já a třída II bílkoviny.[2] Peptidové sekvence se mohou u jednotlivců lišit a tyto rozdíly vznikají SNP v kódující oblasti genů, genové delece, mutace posunu snímků nebo vložení.[3] Asi třetina charakterizovaných MiHA pochází z chromozomu Y.[4] Proteiny jsou složeny z jediné imunogenní alely HLA.[2] Předtím, než se stanou krátkou peptidovou sekvencí, musí být proteiny exprimované těmito polymorfními nebo rozmanitými geny štěpeny v proteazom na kratší peptidy. Tyto endogenní nebo vlastní peptidy jsou poté transportovány do endoplazmatické retikulum s tzv. pumpou na transport peptidů TAP kde se setkávají a váží se na molekulu MHC I. třídy. To kontrastuje s Molekuly MHC třídy II antigeny, které jsou peptidy odvozené od fagocytózy / endocytózy a molekulární degradace proteinů jiných než samostatných entit, obvykle buňkami prezentujícími antigen. Antigeny MiHA jsou buď všudypřítomně exprimovány ve většině tkání, jako je kůže a střeva, nebo restriktivně exprimovány v imunitních buňkách.[5]
Drobné antigeny histokompatibility jsou způsobeny normálními proteiny, které jsou samy o sobě polymorfní v dané populaci. I když jsou dárce a příjemce transplantátu ve vztahu k jejich totožní hlavní komplex histokompatibility geny, rozdíly aminokyselin v Méně důležitý proteiny mohou způsobit pomalé odmítnutí naroubované tkáně. Několik identifikovaných MiHAs kódovaných autosomálně a chromozomem Y[4]
Známé menší antigeny histokompatibility
Následující tabulka uvádí známé MiHA, variantu genů kódujících MiHA peptidy a jejich omezené alely HLA.
| MiHA ID | MiHA peptid | Omezená HLA | Chromozóm | Koordinovat | SNP ID | Gen | Ensembl Gene ID |
|---|---|---|---|---|---|---|---|
| HA-1 / A2 | VL [H / R] DDLLEA | A * 02: 01 | chr19 | 1068739 | rs1801284 | HMHA1 | ENSG00000180448 |
| HA-2 | YIGEVLVS [V / M] | A * 02: 01 | chr7 | 44977022 | rs61739531 | MYO1G | ENSG00000136286 |
| HA-8 | [R / P] TLDKVLEV | A * 02: 01 | chr9 | 2828765 | rs2173904 | KIAA0020 | ENSG00000080608 |
| HA-3 | V [T / M] EPGTAQY | A * 01: 01 | chr15 | 85579423 | rs2061821 | AKAP13 | ENSG00000170776 |
| C19ORF48 | CIPPD [S / T] LLFPA | A * 02: 01 | chr19 | 50798945 | rs3745526 | C19ORF48 | ENSG00000167747 |
| LB-ADIR-1F | SVAPALAL [F / S] PA | A * 02: 01 | chr1 | 179082165 | rs2296377 | TOR3A | ENSG00000186283 |
| LB-HIVEP1-1S | SLPKH [S / N] VTI | A * 02: 01 | chr6 | 12123016 | rs2228220 | HIVEP1 | ENSG00000095951 |
| LB-NISCH-1A | ALAPAP [A / V] EV | A * 02: 01 | chr3 | 52489389 | rs887515 | NISCH | ENSG00000010322 |
| LB-SSR1-1S | [S / L] LAVAQDLT | A * 02: 01 | chr6 | 7310026 | rs10004 | SSR1 | ENSG00000124783 |
| LB-WNK1-1I | RTLSPE [I / M] ITV | A * 02: 01 | chr12 | 889199 | rs12828016 | WNK1 | ENSG00000060237 |
| T4A | GLYTYWSAG [A / E] | A * 02: 01 | chr3 | 140688418 | rs9876490 | TRIM42 | ENSG00000155890 |
| UTA2-1 | QL [L / P] NSVLTL | A * 02: 01 | chr12 | 31981704 | rs2166807 | KIAA1551 | ENSG00000174718 |
| PANE1 | RVWDLPGVLK | A * 03: 01 | chr22 | 41940168 | rs5758511 | CENPM | ENSG00000100162 |
| SP110 | SLP [R / G] GTSTPK | A * 03: 01 | chr2 | 230207994 | rs1365776 | SP110 | ENSG00000135899 |
| ACC-1C | DYLQ [Y / C] VLQI | A * 24: 02 | chr15 | 79971064 | rs1138357 | BCL2A1 | ENSG00000140379 |
| ACC-1Y | DYLQ [Y / C] VLQI | A * 24: 02 | chr15 | 79971064 | rs1138357 | BCL2A1 | ENSG00000140379 |
| P2RX7 | WFHHC [H / R] PKY | A * 29: 02 | chr12 | 121167552 | rs7958311 | P2RX7 | ENSG00000089041 |
| ACC-4 | ATLPLLCA [R / G] | A * 31: 01 | chr15 | 78944951 | rs2289702 | CTSH | ENSG00000103811 |
| ACC-5 | WATLPLLCA [R / G] | A * 33: 03 | chr15 | 78944951 | rs2289702 | CTSH | ENSG00000103811 |
| LB-APOBEC3B-1K | [K / E] PQYHAEMCF | B * 07: 02 | chr22 | 38985821 | rs2076109 | APOBEC3B | ENSG00000179750 |
| LB-ARHGDIB-1R | LPRACW [R / P] EA | B * 07: 02 | chr12 | 14942624 | rs4703 | ARHGDIB | ENSG00000111348 |
| LB-BCAT2-1R | QP [R / T] RALLFVIL | B * 07: 02 | chr19 | 48799813 | rs11548193 | BCAT2 | ENSG00000105552 |
| LB-EBI3-1I | RPRARYY [I / V] QV | B * 07: 02 | chr19 | 4236999 | rs4740 | EBI3 | ENSG00000105246 |
| LB-ECGF-1H | RP [H / R] LETADLO | B * 07: 02 | chr22 | 50525826 | rs112723255 | TYP | ENSG00000025708 |
| LB-ERAP1-1R | HPRQEQIALLA | B * 07: 02 | chr5 | 96803547 | rs26653 | ERAP1 | ENSG00000164307 |
| LB-FUCA2-1V | RLRQ [V / M] GSWL | B * 07: 02 | chr6 | 143502020 | rs3762002 | FUCA2 | ENSG00000001036 |
| LB-GEMIN4-1V | FPALRFVE [V / E] | B * 07: 02 | chr17 | 746265 | rs4968104 | GEMIN4 | ENSG00000179409 |
| LB-PDCD11-1F | GPDSSKT [F / L] LCL | B * 07: 02 | chr10 | 103434329 | rs2986014 | PDCD11 | ENSG00000148843 |
| LB-TEP1-1S | APDGAKVA [S / P] L | B * 07: 02 | chr14 | 20383870 | rs1760904 | TEP1 | ENSG00000129566 |
| LRH-1 | TPNQRQNVC | B * 07: 02 | chr17 | 3690983 | rs3215407 | P2X5 | ENSG00000083454 |
| ZAPHIR | IPRDSWWVEL | B * 07: 02 | chr19 | 57492212 | rs2074071 | ZNF419 | ENSG00000105136 |
| HEATR1 | ISKERA [E / G] AL | B * 08: 01 | chr1 | 236554626 | rs2275687 | HEATR1 | ENSG00000119285 |
| HA-1 / B60 | KECVL [H / R] DDL | B * 40: 01 | chr19 | 1068739 | rs1801284 | HMHA1 | ENSG00000180448 |
| LB-SON-1R | SETKQ [R / C] TVL | B * 40: 01 | chr21 | 33553954 | rs13047599 | SYN | ENSG00000159140 |
| LB-SWAP70-1Q | MEQLE [Q / E] LEL | B * 40: 01 | chr11 | 9748015 | rs415895 | SWAP70 | ENSG00000133789 |
| LB-TRIP10-1EPC | G [E / G] [P / S] QDL [C / G] TL | B * 40: 01 | chr19 | 6751268 | rs1049229 | TRIP10 | ENSG00000125733 |
| SLC1A5 | AE [A / P] TANGGLAL | B * 40: 02 | chr19 | 46787917 | rs3027956 | SLC1A5 | ENSG00000105281 |
| ACC-2 | KEFED [D / G] IINW | B * 44: 03 | chr15 | 79970875 | rs3826007 | BCL2A1 | ENSG00000140379 |
| ACC-6 | MEIFIEVFSHF | B * 44: 03 | chr18 | 63953532 | rs9945924 | HMSD | ENSG00000221887 |
| HB-1H | EEKRGSL [H / Y] VW | B * 44: 03 | chr5 | 143820488 | rs161557 | HMHB1 | ENSG00000158497 |
| HB-1Y | EEKRGSL [H / Y] VW | B * 44: 03 | chr5 | 143820488 | rs161557 | HMHB1 | ENSG00000158497 |
| DPH1 | S [V / L] LPEVDVW | B * 57: 01 | chr17 | 2040586 | rs35394823 | DPH1 | ENSG00000108963 |
| UTDP4-1 | R [I / N] LAHFFCGW | DPB1 * 04 | chr9 | 128721272 | rs11539209 | ZDHHC12 | ENSG00000160446 |
| CD19 | WEGEPPC [L / V] str | DQB1 * 02: 01 | chr16 | 28933075 | rs2904880 | CD19 | ENSG00000177455 |
| LB-PI4K2B-1S | SRSS [S / P] AELDRSR | DQB1 * 06: 03 | chr4 | 25234395 | rs313549 | PI4K2B | ENSG00000038210 |
| LB-MTHFD1-1Q | SSIIAD [Q / R] IALKL | DRB1 * 03: 01 | chr14 | 64442127 | rs2236225 | MTHFD1 | ENSG00000100714 |
| LB-LY75-1K | LGITYR [N / K] KSLMWF | DRB1 * 13: 01 | chr2 | 159819916 | rs12692566 | LY75 | ENSG00000054219 |
| SLC19A1 | [R / H] LVCYLCFY | DRB1 * 15: 01 | chr21 | 45537880 | rs1051266 | SLC19A1 | ENSG00000173638 |
| LB-PTK2B-1T | VYMND [T / K] SPLTPEK | DRB3 * 01: 01 | chr8 | 27451068 | rs751019 | PTK2B | ENSG00000120899 |
| LB-MR1-1R | YFRLGVSDPI [R / H] G | DRB3 * 02: 02 | chr1 | 181049100 | rs2236410 | MR1 | ENSG00000153029 |
Reakce T buněk na MiHA
MiHA vázané na MHC přítomné na buněčném povrchu mohou být rozpoznány jako vlastní peptidy nebo nejsou rozpoznány ani jedním CD8 + nebo CD4 + T buňky. Nedostatek rozpoznání T buňky tomuto vlastnímu antigenu je důvodem, proč jsou alogenní kmenové buňky transplantace pro HLA shodný gen nebo vyvíjející se plod MiHAs během těhotenství nemusí být T buňkami rozpoznány a označeny jako cizí, což vede k imunitní odpovědi. Ačkoli receptory B buněk mohou také rozpoznávat MHC, zdá se, že imunitní odpovědi vyvolávají pouze T buňky.[6] Důsledky imunitní odpovědi jsou patrné u alogenní transplantace hematopoetických kmenových buněk (HCT), když se peptidy kódované polymorfními geny liší mezi příjemcem a dárcovskými T buňkami. Výsledkem je, že dárcovské T buňky mohou cílit na buňky příjemce, které se říká nemoc štěpu proti hostiteli (GVHD ).[5] Ačkoli odmítnutí štěpu nebo kostní dřeně může mít škodlivé účinky, existují výhody imunoterapie, když jsou cytotoxické T lymfocyty specifické pro vlastní antigen a mohou cílit na antigeny exprimované selektivně na leukemických buňkách, aby zničily tyto nádorové buňky označované jako účinek štěpu proti leukémii (GVL).[3]
Rozpoznání zralých T buněk na tento vlastní antigen by nemělo vyvolat imunitní odpověď. Během brzlíkové selekce vyskytující se v brzlíku, a thymocyt TCR, který rozpoznává buď MHC molekulu třídy I nebo třídy II plus peptid, by měl přežít pozitivní výběr. Existuje však smrt apoptóza thymocytů, které neinteragují s molekulami MHC nebo mají vysokoafinitní receptory pro vlastní MHC plus vlastní antigen, proces označovaný jako negativní výběr. Proces pozitivní a negativní selekce tedy znamená, že méně samovolně reagujících zralých T buněk opustí brzlík a povede k autoimunitním problémům.
Objev MiHA
Po transplantaci byl rozpoznán význam MiHAs v imunitní odpovědi. Příjemce vyvinul GVHD navzdory tomu, že měl geny shodné s HLA na lokusu hlavní histokompatibility. Experiment vyvolal otázky ohledně možnosti existence MiHA. Přesněji řečeno, první MiHA byl objeven, když došlo k transplantaci kostní dřeně mezi opačnými pohlavími. Ženská příjemce získala buňky kostní dřeně shodné s MHC, ale stále měla aktivní cytotoxické T buňky (CD8 +).[3] CD8 + T buňky byly aktivní a cílené na mužské buňky kostní dřeně. Bylo zjištěno, že buňky mužské kostní dřeně představují peptid v MHC drážce kódované genem na chromozomu Y. Peptid byl cizí ženským T buňkám a samicím chybí chromozom Y, a tedy i tento MiHA. MiHA kódované chromozomem Y jsou známé jako antigeny HY.[3]
H-Y antigen
Antigeny H-Y jsou kódovány geny na chromozomu Y. Bylo zjištěno, že obě antigeny alely HLA třídy I a II. Některé z těchto antigenů jsou všudypřítomně exprimovány v jaderných mužských buňkách a přítomnost těchto antigenů byla spojena s vyšším rizikem rozvoje transplantace alogenních kmenových buněk GVHD pro gen odpovídající HLA, pokud existuje mužský příjemce a ženská dárkyně.[7] H-Y MiHA hraje roli v těhotenství u mužského plodu, protože fetální buňky mohou přecházet z placenty do mateřského krevního oběhu, kde mateřské T buňky reagují na cizí antigen přítomný jak na MHC třídy I, tak na II. Proto se H-Y specifické CD8 + T buňky vyvíjejí v mateřské krvi a mohou cílit na fetální buňky s jádrem exprimujícím antigen na molekule MHC třídy I. Odpověď na tyto fetální antigeny H-Y souvisí se ženami, u nichž došlo k sekundárnímu opakujícímu se potratu a které byly dříve těhotné s mužským plodem.[3] Ženy s dřívějším mužským těhotenstvím mají T buňky, které byly dříve vystaveny těmto H-Y antigenům, a následně je rozpoznávají rychleji. Bylo zjištěno, že ženy s opakovaným potratem také obsahují MHC II se schopností prezentovat tyto antigeny T pomocným buňkám (CD4 +), což je významné pro aktivaci CD8 +.[8]
Histocompatibility Antigen 1 (HA1)
HA1 je výsledkem a SNP konvertování neimunogenní alely (KECVLRDDLLEA) na imunogenní alelu (KECVLHDDLLEA). Výsledkem tohoto SNP je lepší schopnost vázat peptid do drážky konkrétní molekuly MHC třídy I nacházející se na buňkách prezentujících antigen.[5] Význam změny peptidu na imunogenní formu spočívá v tom, že nyní specifické T buňky s omezeným HLA-A 0201 mohou rozpoznat peptid prezentovaný molekulami HLA-A0201 MHC třídy I. Toto rozpoznání vede k imunitní odpovědi, pokud T buňky rozpoznají peptid jako cizí. K tomuto rozpoznání dochází, když jednotlivci chybí imunogenní verze peptidu, ale je vystaven peptidu HA-1 během těhotenství nebo alogenní transplantace kmenových buněk. Během těhotenství bylo zjištěno, že fetální HA-1 pochází z placenty a byly identifikovány specifické mateřské CD8 + T buňky rozpoznávající tuto MiHA.[5]
Účinek imunoterapie Graft-Versus-Leukemia
CD8 + T buňky, které jsou specifické pro MiHA, mohou cílit na tyto antigeny, když jsou specificky exprimovány na nádorových buňkách, což umožňuje destrukci škodlivých nádorových buněk. U myší bylo prokázáno, že dárce alogenních kmenových buněk CD8 + T buňky specifické pro MiHA nalezené u příjemce inhibují dělení leukemický buňky. Existuje však riziko rozvoje GVHD, pokud jsou T buňky specifické pro MiHA exprimované všudypřítomně epiteliální buňky. Přesněji řečeno, HA-8, UGT2B17 a SMCY MiHA, které jsou všudypřítomně exprimovány, představují vyšší riziko vzniku GVHD. Proto, aby se zabránilo nepříznivým účinkům GVHD, jsou MiHA omezené na imunitní buňky ideálním cílem pro štěp versus leukemie (GVL), protože ne všechny nukleované buňky jsou cíleny odpovědí T buněk. Příkladem ideálního cíle je MiHA HB-1, který je vysoce exprimován ve škodlivých B buňkách, ale má nízkou expresi v jiných tkáňových buňkách.[9]
Klinické důsledky
Imunizace matek proti mužským specifickým antigenům menší histokompatibility (HY) má patogenní roli v mnoha případech sekundární opakovaný potrat, tj. opakovaný potrat v těhotenství, které následovalo po předchozím živém porodu. Příkladem tohoto účinku je, že poměr mužů a žen u dětí narozených před a po sekundárním opakujícím se potratu je 1,49, respektive 0,76.[10]
Viz také
Reference
- ^ Robertson NJ, Chai JG, Millrain M, Scott D, Hashim F, Manktelow E, Lemonnier F, Simpson E, Dyson J (březen 2007). „Přirozená regulace imunity na menší antigeny histokompatibility“. Journal of Immunology. 178 (6): 3558–65. doi:10,4049 / jimmunol.178.6.3558. PMID 17339452.
- ^ A b Dzierzak-Mietla M, Markiewicz M, Siekiera U, Mizia S, Koclega A, Zielinska P, Sobczyk-Kruszelnicka M, Kyrcz-Krzemien S (2012). „Výskyt a dopad drobných histokompatibilních rozdílů antigenů na výsledky transplantace hematopoetických kmenových buněk od dárců sourozenců odpovídajících HLA“. Výzkum kostní dřeně. 2012: 257086. doi:10.1155/2012/257086. PMC 3502767. PMID 23193478.
- ^ A b C d E Linscheid C, Petroff MG (duben 2013). „Drobné histokompatibilní antigeny a imunitní reakce matky na plod během těhotenství“. American Journal of Reproductive Immunology. 69 (4): 304–14. doi:10.1111 / aji.12075. PMC 4048750. PMID 23398025.
- ^ A b Hirayama M, Azuma E, Komada Y (2012). Hlavní a vedlejší histokompatibilní antigeny k nezděděným mateřským antigenům (NIMA), histokompatibilita. INTECH. p. 146. ISBN 978-953- 51-0589-3.
- ^ A b C d Bleakley M, Riddell SR (březen 2011). "Využití T buněk specifických pro lidské menší histokompatibilní antigeny pro léčbu leukémie". Imunologie a buněčná biologie. 89 (3): 396–407. doi:10.1038 / icb.2010.124. PMC 3061548. PMID 21301477.
- ^ Perreault C, Décary F, Brochu S, Gyger M, Bélanger R, Roy D (1990). „Drobné antigeny histokompatibility“ (PDF). Krev. 76 (7): 1269–80. PMID 2207305.
- ^ Nielsen HS (01.07.2011). „Sekundární opakovaný potrat a imunita vůči HY“. Aktualizace lidské reprodukce. 17 (4): 558–74. doi:10.1093 / humupd / dmr005. PMID 21482560.
- ^ Lissauer D, Piper K, Goodyear O, Kilby MD, Moss PA (červenec 2012). „Fetálně specifické CD8 + cytotoxické reakce T buněk se vyvíjejí během normálního lidského těhotenství a vykazují širokou funkční kapacitu“. Journal of Immunology. 189 (2): 1072–80. doi:10,4049 / jimmunol.1200544. PMID 22685312.
- ^ Bleakley M, Riddell SR (2004). "Molekuly a mechanismy účinku štěpu proti leukémii". Recenze přírody. Rakovina. 4 (5): 371–80. doi:10.1038 / nrc1365. PMID 15122208.
- ^ Nielsen HS (2011). „Sekundární opakovaný potrat a imunita vůči HY“. Aktualizace lidské reprodukce. 17 (4): 558–74. doi:10.1093 / humupd / dmr005. PMID 21482560.
externí odkazy
- Menší + histokompatibilita + antigeny v americké národní lékařské knihovně Lékařské předměty (Pletivo)