Mutace posunu snímků - Frameshift mutation

A mutace posunu snímků (také nazývaný a chyba rámování nebo a posun čtecího rámečku) je genetická mutace zapříčiněno indels (vložení nebo vypuštění ) z řady nukleotidy v sekvenci DNA, která není dělitelná třemi. Vzhledem k tripletové povaze genová exprese podle kodony, vložení nebo smazání může změnit čtecí rámec (seskupení kodonů), což vedlo k úplně jinému překlad z originálu. Čím dříve v sekvenci dojde k deleci nebo inzerci, tím více se protein změní.[1] Mutace posunu snímků není stejná jako a jedno-nukleotidový polymorfismus ve kterém je nukleotid nahrazen, místo aby byl vložen nebo odstraněn. A shifthift mutace obecně způsobí, že čtení kodonů po mutaci bude kódovat různé aminokyseliny. Mutace posunu snímků také změní první stop kodon („UAA“, „UGA“ nebo „UAG“), který se v sekvenci vyskytne. Vytvářený polypeptid může být neobvykle krátký nebo neobvykle dlouhý a pravděpodobně nebude funkční.[2]
Mutace posunu rámu jsou patrné u závažných genetických onemocnění, jako je Tay – Sachsova choroba; zvyšují náchylnost k určitým druhům rakoviny a třídám familiární hypercholesterolemie; v roce 1997,[3] mutace posunu snímků byla spojena s odolností proti infekci HIV retrovirem. Mutace posunu snímků byly navrženy jako zdroj biologické novinky, stejně jako u údajného vytvoření nylonase tato interpretace je však kontroverzní. Studie Negoro et al (2006)[4] zjistili, že je nepravděpodobné, že by byla příčinou mutace posunu snímků, a že spíše substituce dvou aminokyselin v DNA Aktivní stránky předků esteráza vedlo k nylonase.
Pozadí
Informace obsažené v DNA určují funkci bílkovin v buňkách všech organismů. Transkripce a translace umožňují přenos těchto informací do tvorby proteinů. Chyba při čtení této komunikace však může způsobit nesprávnou funkci proteinu a nakonec způsobit onemocnění, i když buňka obsahuje řadu nápravných opatření.
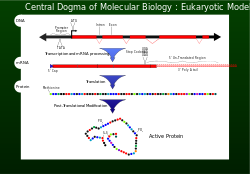
Centrální dogma
V roce 1956 Francis Crick popsal tok genetické informace z DNA na specifické uspořádání aminokyselin pro výrobu a protein jako ústřední dogma.[1] Aby buňka správně fungovala, musí být proteiny produkovány přesně pro strukturální a pro katalytické činnosti. Nesprávně vyrobený protein může mít škodlivé účinky na buňka životaschopnost a ve většině případů způsobí vyšší organismus stát se nezdravým abnormálními buněčnými funkcemi. Aby bylo zajištěno, že genom úspěšně předává informace o korektura mechanismy jako exonukleázy a oprava nesouladu systémy jsou začleněny do replikace DNA .[1]
Přepis a překlad
Po replikaci DNA je čtení vybrané části genetické informace dosaženo pomocí transkripce.[1]Nukleotidy obsahující genetickou informaci jsou nyní na jednovláknové messengerové šabloně zvané mRNA. MRNA je začleněna do podjednotky ribozom a komunikuje s rRNA. Genetická informace přenášená v kodonech mRNA je nyní čtena (dekódována) antikodony tRNA. Jak se čte každý kodon (triplet), aminokyseliny jsou spojeny dohromady až do stop kodon (UAG, UGA nebo UAA) je dosaženo. V tomto bodě polypeptid (protein) byl syntetizován a uvolněn.[1] Na každých 1 000 aminokyselin zabudovaných do proteinu je nesprávná maximálně jedna. Této věrnosti rozpoznávání kodonů, při zachování důležitosti správného čtecího rámce, je dosaženo správným párováním bází v místě ribozomu A, GTP aktivita hydrolýzy EF-Tu forma kinetické stability a korekturní mechanismus při uvolnění EF-Tu.[1]
K posunu snímků může dojít také během profáze translace, produkující různé proteiny z překrývajících se otevřených čtecích rámců, jako je gag-pol-env retrovir bílkoviny. To je docela běžné v viry a také se vyskytuje v bakterie a droždí (Farabaugh, 1996). Reverzní transkriptáza, naproti tomu RNA polymeráza II, se považuje za silnější příčinu výskytu mutací posunu snímků. V experimentech se vyskytlo pouze 3–13% všech mutací posunu snímků kvůli RNA Polymerase II. v prokaryoty chybovost vyvolávající mutace posunu snímků je pouze někde v rozsahu 0,0001 a 0,00001.[5]
Existuje několik biologických procesů, které pomáhají předcházet mutacím posunu snímků. Nastanou reverzní mutace, které změní mutovanou sekvenci zpět na původní divoký typ sekvence. Další možností pro korekci mutace je použití a potlačující mutace. To kompenzuje účinek původní mutace vytvořením sekundární mutace posunutím sekvence tak, aby bylo možné číst správné aminokyseliny. Průvodce RNA lze také použít k vložení nebo odstranění uridinu do mRNA po transkripci, což umožňuje správný čtecí rámec.[1]
Význam kodon-triplet

A kodon je sada tří nukleotidy, triplet, který kód pro určité aminokyselina. První kodon stanoví čtecí rámec, čímž začíná nový kodon. Páteř aminokyselin bílkoviny sekvence je definována souvislými trojčaty.[6] Kodony jsou klíčem k překladu genetické informace pro syntézu proteinů. Čtecí rámec je nastaven, když začíná překlad mRNA, a je udržován, když čte jednu triplet k další. Čtení genetického kódu podléhá třem pravidlům, která sledují kodony v mRNA. Nejprve se kodony čtou ve směru 5 'až 3'. Zadruhé, kodony se nepřekrývají a zpráva nemá žádné mezery. Poslední pravidlo, jak je uvedeno výše, že zpráva je přeložena do pevného čtecího rámce.[1]
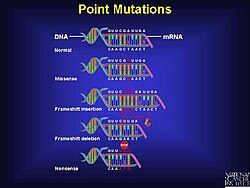
Mechanismus
Mutace posunu snímků mohou nastat náhodně nebo mohou být způsobeny vnějším podnětem. K detekci mutací posunu snímků může dojít několika různými metodami. Posunutí snímků je jen jedním typem mutace, která může vést k neúplným nebo nesprávným proteinům, ale představují významné procento chyb v DNA.
Genetické nebo environmentální
Jedná se o genetickou mutaci na úrovni nukleotidových bází. Proč a jak dochází k mutacím posunu snímků je neustále hledáno. Environmentální studie, konkrétně výroba UV Byly provedeny indukované mutace posunu rámců DNA polymerázami s deficitem 3 ′ → 5 ′ exonukleázové aktivity. Normální sekvence 5 'GTC GTT TTA CAA 3' byla změněna na GTC GTT T TTA CAA (MIDT) z GTC GTT C TTA CAA (MIDC), aby se studovaly posuny rámců. E-coli pol I mutant Kf a T7 DNA polymerázy enzymy postrádající 3 '→ 5' exonukleázovou aktivitu produkují UV-indukované revertanty s vyšší frekvencí než jejich exonukleáza zdatní protějšky. Data naznačují, že ztráta korektury zvyšuje frekvenci posunů rámů vyvolaných UV zářením.[7]
Detekce
Fluorescence
Účinky sousedních bází a sekundární struktury pro detekci frekvence mutací posunu snímků byly podrobně zkoumány pomocí fluorescence. Fluorescenčně značená DNA pomocí analogů bazí umožňuje studovat lokální změny sekvence DNA.[8] Studie účinků délky řetězce primeru odhalily, že byla pozorována rovnovážná směs čtyř hybridizačních konformací, když se báze templátu smyčkovitě vyklenuly, tj. Struktura lemovaná na obou stranách duplexní DNA. Naproti tomu byla struktura dvojité smyčky s neobvyklou neskládanou konformací DNA na jejím dolním okraji pozorována, když byly extrudované báze umístěny na spojení primer - templát, což ukazuje, že odchylky lze upravit sousední sekundární strukturou DNA.[9]
Sekvenování

Sangerovo sekvenování a pyrosekvenování jsou dvě metody, které byly použity k detekci mutací posunu snímků, je však pravděpodobné, že generovaná data nebudou mít nejvyšší kvalitu. I přesto 1,96 milionu indels byly identifikovány pomocí Sangerova sekvenování, které se nepřekrývají s jinými databázemi. Když je pozorována mutace posunu snímků, je porovnána s databází mutací lidského genomu (HGMD), aby se určilo, zda má mutace škodlivý účinek. To se děje při pohledu na čtyři funkce. Za prvé, poměr mezi ovlivněnou a konzervovanou DNA, za druhé umístění mutace vzhledem k transkriptu, za třetí poměr konzervovaných a ovlivněných aminokyselin a nakonec vzdálenost indelu ke konci exon.[10]
Masivně paralelní sekvenování je novější metoda, kterou lze použít k detekci mutací. Pomocí této metody lze sekvenovat až 17 gigabází najednou, na rozdíl od omezených rozsahů pro Sangerovo sekvenování jen asi 1 kilobase. K provedení tohoto testu je k dispozici několik technologií a zkoumá se jeho použití v klinických aplikacích.[11] Při testování různých karcinomů umožňují současné metody prohlížení pouze jednoho genu najednou. Masivně paralelní sekvenování může testovat různé mutace způsobující rakovinu najednou, na rozdíl od několika konkrétních testů.[12] Experiment ke stanovení přesnosti této nové metody sekvenování byl testován na 21 genech a neměl žádné falešně pozitivní výzvy k mutacím posunu snímků.[13]
Diagnóza
USA patent (5 958 684) v roce 1999 Leeuwen, podrobně popisuje metody a činidla pro diagnostiku nemocí způsobených nebo spojených s genem, který má somatickou mutaci vedoucí k mutaci rámcového posunu. Metody zahrnují poskytnutí vzorku tkáně nebo tekutiny a provedení genové analýzy pro mutaci posunu snímků nebo protein z tohoto typu mutace. Nukleotidová sekvence podezřelého genu je poskytována z publikovaných genových sekvencí nebo z klonování a sekvenování podezřelého genu. Potom se předpovídá aminokyselinová sekvence kódovaná genem.[14]
Frekvence
Navzdory pravidlům, kterými se řídí genetický kód a různé mechanismy přítomné v buňce k zajištění správného přenosu genetické informace během procesu replikace DNA i během translace, dochází k mutacím; mutace posunu snímků není jediným typem. Existují alespoň dva další typy rozpoznaných bodových mutací, konkrétně missense mutace a nesmyslná mutace.[1] Mutace posunu snímků může drasticky změnit kapacitu kódování (genetickou informaci) zprávy.[1] Malé inzerce nebo delece (ty, které mají méně než 20 párů bází) tvoří 24% mutací, které se projevují v současné době uznávaným genetickým onemocněním.[10]
Zjistilo se, že mutace posunu snímků jsou častější v opakovaných oblastech DNA. Důvodem je sklouznutí polymerázového enzymu v opakovaných oblastech, což umožňuje mutacím vstoupit do sekvence.[15] Experimenty lze spustit k určení frekvence mutace posunu snímků přidáním nebo odstraněním přednastaveného počtu nukleotidů. Experimenty byly spuštěny přidáním čtyř párů bází, nazývaných experimenty +4, ale tým z Emory University zkoumal rozdíl ve frekvenci mutace přidáním a odstraněním párů bází. Ukázalo se, že mezi přidáním a odstraněním páru bází nebyl žádný rozdíl ve frekvenci. Existuje však rozdíl v konečném výsledku proteinu.[15]
Huntingtonova choroba je jednou z devíti poruch kodonové reiterace způsobených polyglutaminovými expanzními mutacemi, které zahrnují spino-cerebelární ataxii (SCA) 1, 2, 6, 7 a 3, spinobulbární svalovou atrofii a dentatorubal-palidoluysianatrofii. Může existovat souvislost mezi chorobami způsobenými polyglutaminovými a polyalaninovými expanzními mutacemi, jako je posunutí původního produktu genu SCA3 kódujícího CAG / polyglutaminy na GCA / polyalaniny. Ribozomální sklouznutí během translace proteinu SCA3 bylo navrženo jako mechanismus vedoucí k přesunu z polyglutaminu do rámce kódujícího polyalanin. Dinukleotidová delece nebo vložení jediného nukleotidu do polyglutaminového traktu huntingtinového exonu 1 by posunuly CAG, polyglutaminen kódující rámec o +1 (+1 rámcový posun) na GCA, polyalanin kódující rámec a zavedly nový epitop na C konec Htt exon 1 (APAAAPAATRPGCG).[16]
Nemoci
Některé nemoci mají alespoň část příčiny mutace posunu snímků. Znalost převládajících mutací může také pomoci při diagnostice onemocnění. V současné době existují pokusy využít mutace posunu snímků příznivě při léčbě nemocí a změnit čtecí rámec aminokyselin.
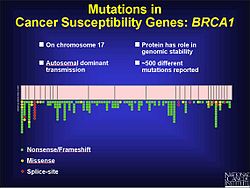

Rakovina
Je známo, že mutace posunu snímků jsou faktorem kolorektální rakovina i další rakoviny s nestabilita mikrosatelitu. Jak již bylo uvedeno, mutace posunu snímků se pravděpodobně vyskytují v oblasti opakující se sekvence. Když oprava nesouladu DNA neopraví přidání nebo deleci bází, jsou tyto mutace pravděpodobněji patogenní. Může to být částečně proto, že nádoru není řečeno, aby přestal růst. Experimenty s kvasinkami a bakteriemi pomáhají ukázat charakteristiky mikrosatelitů, které mohou přispět k opravě chybného párování DNA. Patří mezi ně délka mikrosatelit, složení genetického materiálu a čistota opakování. Na základě experimentálních výsledků mají delší mikrosatelity vyšší rychlost mutací posunu snímků. Lemující DNA může také přispět k mutacím posunu snímků.[17] U rakoviny prostaty mutace posunu snímků mění otevřený čtecí rámec (ORF) a brání apoptóza od výskytu. To vede k neregulovanému růstu nádor. I když existují faktory prostředí, které přispívají k progresi rakovina prostaty, existuje také genetická složka. Během testování kódujících oblastí k identifikaci mutací bylo objeveno 116 genetických variant, včetně 61 mutací posunu snímků.[18] Na chromozomu 17 existuje více než 500 mutací, které podle všeho hrají roli ve vývoji rakoviny prsu a vaječníků v genu BRCA1, z nichž mnohé jsou posunuty.[19]
Crohnova nemoc
Crohnova nemoc má asociaci s genem NOD2. Mutace je vložením a Cytosin v poloze 3020. To vede k předčasnému stop kodonu, což zkracuje protein, který má být transkribován. Když je protein schopen normálně se tvořit, reaguje na bakteriální liposacharidy, kde mutace 3020insC brání tomu, aby protein reagoval.[20]
Cystická fibróza
Cystická fibróza (CF) je onemocnění založené na mutacích v CF transmembránový gen regulátoru vodivosti (CFTR). Bylo identifikováno více než 1 500 mutací, ale ne všechny způsobují onemocnění.[21] Většina případů cystické fibrózy je výsledkem mutace ∆F508, která odstraní celou aminokyselinu. Při diagnostice CF, CF1213delT a CF1154-insTC jsou zajímavé dvě mutace posunu snímků. Obě tyto mutace se běžně vyskytují v tandemu s alespoň jednou další mutací. Oba vedou k malému snížení funkce plíce a vyskytují se asi u 1% testovaných pacientů. Tyto mutace byly identifikovány pomocí Sangerova sekvenování.[22]
HIV
CCR5 je jedním z kofaktorů vstupu buněk spojených s HIV, nejčastěji u kmenů indukujících nesyncytium, je nejvíce patrný u pacientů s HIV na rozdíl od pacientů s AIDS. Delece 32 párů bází v CCR5 byla identifikována jako mutace, která neguje pravděpodobnost infekce HIV. Tato oblast na otevřeném čtecím rámci ORF obsahuje mutaci posunu snímků vedoucí k předčasnému stop kodonu. To vede ke ztrátě funkce HIV-koreceptoru in vitro. CCR5-1 je považován za divoký typ a CCR5-2 je považován za mutantní alelu. Pacienti s heterozygotní mutací pro CCR5 byli méně náchylní k rozvoji HIV. Ve studii, navzdory vysoké expozici viru HIV, nebyl nikdo homozygotní pro mutaci CCR5, který by měl pozitivní test na HIV.[3]
Tay – Sachsova choroba
Tay – Sachsova choroba je smrtelné onemocnění postihující centrální nervový systém. Nejčastěji se vyskytuje u kojenců a malých dětí. Progrese nemoci začíná v lůno ale příznaky se neobjeví přibližně do 6 měsíců věku. Léčba nemoci neexistuje.[23] Je známo, že mutace v genu β-hexosaminidázy A (Hex A) ovlivňují nástup Tay-Sachs, přičemž je popsáno 78 mutací různých typů, o nichž je známo, že 67 způsobují onemocnění. Většina pozorovaných mutací (65/78) jsou substituce jedné báze nebo SNP, 11 delecí, 1 velká a 10 malých a 2 inzerce. 8 pozorovaných mutací je posun snímků, 6 delecí a 2 inzerce. Inzerce 4 párů bází v exonu 11 je pozorována u 80% přítomnosti Tay-Sachsovy choroby v Ashkenazi Židovská populace. Mutace posunu snímků vedou k časnému stop kodonu, o kterém je známo, že hraje roli v nemoci u kojenců. Zdá se, že onemocnění se zpožděným nástupem je způsobeno 4 různými mutacemi, z nichž jedna je delecí 3 bazických párů.[24]
Smith-Magenisův syndrom
Smith-Magenisův syndrom (SMS) je komplex syndrom zahrnující intelektuální postižení, poruchy spánku, problémy s chováním a různé kraniofaciální, skeletální a viscerální anomálie. Většina případů SMS obsahuje ~ 3,5 Mb společné delece, která zahrnuje gen indukovaný-1 (RAI1) kyselinou retinovou. Jiné případy ilustrují variabilitu SMS fenotyp dříve nebylo prokázáno pro mutaci RAI1, včetně ztráty sluchu, absence sebepoškozujícího chování a mírných globálních zpoždění. Sekvenování RAI1 odhalilo mutaci heptamerního C-traktu (CCCCCCC) v exonu 3, což mělo za následek mutace posunu snímků. Ze sedmi hlášených mutací posunu rámce vyskytujících se v poly C-traktech v RAI1 se v tomto heptamerním C-traktu vyskytují čtyři případy (~ 57%). Výsledky ukazují, že tento heptamerický C-trakt je preferenční rekombinační hotspot inzerce / delece (SNindels), a proto primární cíl pro analýzu u pacientů s podezřením na mutace v RAI1.[25]
Hypertrofické kardiomyopatie
Hypertrofické kardiomyopatie je nejčastější příčinou nenadálá smrt u mladých lidí, včetně trénovaných sportovců, a je způsobena mutacemi genů kódujících proteiny srdeční sarkomery. Mutace v genu pro troponin C (TNNC1) jsou vzácnou genetickou příčinou hypertrofické kardiomyopatie. Nedávná studie ukázala, že mutace posunu snímků (c.363dupG nebo p.Gln122AlafsX30) v troponinu C byla příčinou hypertrofické kardiomyopatie (a náhlé srdeční smrti) u 19letého muže.[26]
Léčí
Hledání léku na nemoci způsobené mutacemi posunu snímků je vzácné. Výzkum v této oblasti pokračuje. Jedním z příkladů je primární imunodeficience (PID), zděděný stav, který může vést ke zvýšení infekcí. V primární imunodeficienci hraje roli 120 genů a 150 mutací. Standardní léčba je v současné době genová terapie, ale jedná se o vysoce riskantní léčbu a může často vést k dalším chorobám, jako je leukémie. Postupy genové terapie zahrnují modifikaci fúzního proteinu nukleázy zinkového prstenu, štěpení obou konců mutace, což ji zase odstraní ze sekvence. Zprostředkován antisense-oligonukleotid přeskakování exonu je pro Duchenne další možnost svalová dystrofie. Tento proces umožňuje předání mutace tak, aby zbytek sekvence zůstal v rámci a funkce proteinu zůstala nedotčena. To však nemoci neléčí, pouze léčí příznaky a je praktické pouze u strukturních proteinů nebo jiných opakujících se genů. Třetí forma opravy je zvrácená mozaika, který se přirozeně vyskytuje vytvořením reverzní mutace nebo mutace na druhém místě, která opravuje čtecí rámec. K této reverzi může dojít intragenicky rekombinace, mitotický genová konverze, uklouznutí DNA z druhého místa nebo reverze specifická pro dané místo. To je možné u několika onemocnění, jako je X-vázaná těžká kombinovaná imunodeficience (SCID), Wiskott – Aldrichův syndrom a Bloomův syndrom. Neexistují žádné léky ani jiné farmakogenomické metody, které by pomohly s PID.[27]
Evropský patent (EP1369126A1) z roku 2003, který napsal Bork, zaznamenává metodu používanou pro prevenci rakoviny a pro kurativní léčbu rakoviny a prekancerózy, jako jsou sporadické nádory s nedostatečnou opravou DNA (MMR) a nádory spojené s HNPCC. Myšlenka je použít imunoterapie s kombinačními směsmi nádorově specifických peptidů odvozených od mutace, které vyvolávají cytotoxickou reakci T-buněk specificky namířenou proti nádorovým buňkám.[28]
Viz také
- Překladový posun snímků
- Mutace
- Přepis (genetika)
- Překlad (biologie)
- kodon
- protein
- čtecí rámec
- bodová mutace
- Crohnova nemoc
- Tay – Sachsova choroba
Reference
- ^ A b C d E F G h i j Losick, Richard; Watson, James D .; Baker, Tania A .; Bell, Stephen; Gann, Alexander; Levine, Michael W. (2008). Molekulární biologie genu (6. vydání). San Francisco: Pearson / Benjamin Cummings. ISBN 978-0-8053-9592-1.
- ^ „DNA se neustále mění procesem mutace“. Příroda. Citováno 17. května 2019.
- ^ A b Zimmerman PA, Buckler-White A, Alkhatib G, Spalding T, Kubofcik J, Combadiere C, Weissman D, Cohen O, Rubbert A, Lam G, Vaccarezza M, Kennedy PE, Kumaraswami V, Giorgi JV, Detels R, Hunter J, Chopek M, Berger EA, Fauci AS, Nutman TB, Murphy PM (leden 1997). „Zděděná rezistence na HIV-1 vyvolaná inaktivující mutací v CC chemokinovém receptoru 5: studie v populacích s kontrastními klinickými fenotypy, definovaným rasovým pozadím a kvantifikovaným rizikem“. Molekulární medicína (Cambridge, Massachusetts). 3 (1): 23–36. PMC 2230106. PMID 9132277.
- ^ Negoro S, Ohki T, Shibata N, Mizuno N, Wakitani Y, Tsurukame J, Matsumoto K, Kawamoto I, Takeo M, Higuchi Y (listopad 2005). „Rentgenová krystalografická analýza 6-aminohexanoát-dimer hydrolázy: molekulární základ pro zrození enzymu degradujícího nylonový oligomer“. J Biol Chem. 280 (47): 39644–52. doi:10,1074 / jbc.m505946200. PMID 16162506.
- ^ Zhang, J (srpen 2004). „Host RNA polymeráza II přispívá minimálně k retrovirovým mutacím s posunem rámce“. The Journal of General Virology. 85 (Pt 8): 2389–95. doi:10.1099 / vir.0.80081-0. PMID 15269381.
- ^ Cox, Michael; Nelson, David R .; Lehninger, Albert L (2008). Lehningerovy principy biochemie. San Francisco: W.H. Freemane. ISBN 978-0-7167-7108-1.
- ^ Sagher, Daphna; Turkington, Edith; Acharya, Sonia; Strauss, Bernard (červenec 1994). "Produkce UV-indukovaných Frameshift mutací in vitro DNA polymerázami s deficitem 3 '→ 5' exonukleázové aktivity". Journal of Molecular Biology. 240 (3): 226–242. doi:10.1006 / jmbi.1994.1437. PMID 8028006.
- ^ Johnson, Neil P .; Walter A. Baase; Peter H. von Hippel (březen 2004). „Nízkoenergetický cirkulární dichroismus 2-aminopurin-dinukleotidu jako sonda lokální konformace DNA a RNA“. Proc Natl Acad Sci U S A. 101 (10): 3426–31. doi:10.1073 / pnas.0400591101. PMC 373478. PMID 14993592.
- ^ Baase, Walter A .; Davis Jose; Benjamin C. Ponedel; Peter H. von Hippel; Neil P. Johnson (2009). „DNA modely trinukleotidových posunů rámových delecí: tvorba smyček a boulí na křižovatce primer-templát“. Výzkum nukleových kyselin. 37 (5): 1682–9. doi:10.1093 / nar / gkn1042. PMC 2655659. PMID 19155277.
- ^ A b Hu, J; Ng, PC (9. února 2012). „Predikce účinků indelů s posunem snímků“. Genome Biology. 13 (2): R9. doi:10.1186 / gb-2012-13-2-r9. PMC 3334572. PMID 22322200.
- ^ Tucker, Tracy; Marra, Marco; Friedman, Jan M. (2009). „Masivně paralelní sekvenování: další velká věc v genetické medicíně“. American Journal of Human Genetics. 85 (2): 142–154. doi:10.1016 / j.ajhg.2009.06.022. PMC 2725244. PMID 19679224.
- ^ Walsh, T .; Casadei, S .; Lee, M. K .; Pennil, C. C .; Nord, A. S .; Thornton, A. M .; Roeb, W .; Agnew, K. J .; Stray, S. M .; Wickramanayake, A .; Norquist, B .; Pennington, K. P .; Garcia, R. L .; King, M.-C .; Swisher, E. M. (2011). „Z obalu: Mutace ve 12 genech pro zděděný ovariální, vejcovod a peritoneální karcinom identifikované masivně paralelním sekvenováním“. Proc Natl Acad Sci U S A. 108 (44): 18032–7. doi:10.1073 / pnas.1115052108. PMC 3207658. PMID 22006311.
- ^ Walsh, T .; Lee, M. K .; Casadei, S .; Thornton, A. M .; Stray, S. M .; Pennil, C .; Nord, A. S .; Mandell, J. B .; Swisher, E. M .; King, M.-C. (2010). „Detekce zděděných mutací pro rakovinu prsu a vaječníků pomocí genomového záchytu a masivního paralelního sekvenování“. Proc Natl Acad Sci U S A. 107 (28): 12629–33. doi:10.1073 / pnas.1007983107. PMC 2906584. PMID 20616022.
- ^ Americký patent 5,958,684 (28. září 1999) „Diagnóza neurodegenerativních onemocnění“ od Leeuwena et al
- ^ A b Harfe, BD; Jinks-Robertson, S (červenec 1999). "Odstranění meziproduktů posunu rámců pomocí neshodných opravných proteinů v Saccharomyces cerevisiae". Molekulární a buněčná biologie. 19 (7): 4766–73. doi:10.1128 / MCB.19.7.4766. PMC 84275. PMID 10373526.
- ^ Davies, J. E.; Rubinsztein, D C (2006). „Produkty s polyalaninem a polyserinem při posunu při Huntingtonově chorobě“. Journal of Medical Genetics. 43 (11): 893–896. doi:10.1136 / jmg.2006.044222. PMC 2563184. PMID 16801344.
- ^ Schmoldt, A; Benthe, HF; Haberland, G (1. září 1975). "Metabolismus digitoxinu mikrosomy jater jater potkana". Biochemická farmakologie. 24 (17): 1639–41. doi:10.1016/0006-2952(75)90094-5. PMID 10.
- ^ Xu, XiaoLin; Zhu, KaiChang; Liu, Feng; Wang, Yue; Shen, JianGuo; Jin, Jizhong; Wang, Zhong; Chen, Lin; Li, Jiadong; Xu, Min (květen 2013). "Identifikace somatických mutací u lidské rakoviny prostaty pomocí RNA-Seq". Gen. 519 (2): 343–7. doi:10.1016 / j.gene.2013.01.046. PMID 23434521.
- ^ „Genomika rakoviny“. National Cancer Institute at the National Institute of Health. Citováno 24. března 2013.
- ^ Ogura Y, Bonen DK, Inohara N, Nicolae DL, Chen FF, Ramos R, Britton H, Moran T, Karaliuskas R, Duerr RH, Achkar JP, Brant SR, Bayless TM, Kirschner BS, Hanauer SB, Nuñez G, Cho JH (31. května 2001). „Mutace posunu snímků v NOD2 spojená s náchylností k Crohnově nemoci“ (PDF). Příroda. 411 (6837): 603–6. doi:10.1038/35079114. hdl:2027.42/62856. PMID 11385577. S2CID 205017657.
- ^ Farrell PM, Rosenstein BJ, White TB, Accurso FJ, Castellani C, Cutting GR, Durie PR, Legrys VA, Massie J, Parad RB, Rock MJ, Campbell PW (2008). „Pokyny pro diagnostiku cystické fibrózy u novorozenců u starších dospělých: Zpráva o shodě nadace pro cystickou fibrózu“. The Journal of Pediatrics. 153 (2): S4 – S14. doi:10.1016 / j.jpeds.2008.05.005. PMC 2810958. PMID 18639722.
- ^ Iannuzzi, MC; Stern, RC; Collins, FS; Hon, CT; Hidaka, N; Silný, T; Becker, L; Drumm, ML; Bílá, MB; Gerrard, B (únor 1991). „Dvě mutace posunu snímků v genu pro cystickou fibrózu“. American Journal of Human Genetics. 48 (2): 227–31. PMC 1683026. PMID 1990834.
- ^ „Učení o Tay-Sachsově nemoci“. Národní institut pro výzkum lidského genomu. Citováno 24. března 2013.
- ^ Myerowitz, R (1997). „Mutace způsobující Tay-Sachsovu chorobu a neutrální polymorfismy v genu Hex A“. Lidská mutace. 9 (3): 195–208. doi:10.1002 / (SICI) 1098-1004 (1997) 9: 3 <195 :: AID-HUMU1> 3.0.CO; 2-7. PMID 9090523.
- ^ Truong, Hoa T; Dudding, Tracy; Blanchard, Christopher L .; Elsea, Sarah H (2010). „Hotspot mutace Frameshift identifikovaný u Smith-Magenisova syndromu: kazuistika a přehled literatury“. Lékařská genetika BMC. 11 (1): 142. doi:10.1186/1471-2350-11-142. PMC 2964533. PMID 20932317.
- ^ Chung WK, Kitner C, Maron BJ (červen 2011). "Nová mutace posunu snímků v troponinu C (TNNC1) spojená s hypertrofickou kardiomyopatií a náhlou smrtí". Cardiol Young. 21 (3): 345–8. doi:10.1017 / S1047951110001927. PMID 21262074.
- ^ Hu, Hailiang; Gatti, Richard A (2008). „Nové přístupy k léčbě primárních imunodeficiencí: stanovení mutací chemickými látkami“. Aktuální názor na alergii a klinickou imunologii. 8 (6): 540–6. doi:10.1097 / ACI.0b013e328314b63b. PMC 2686128. PMID 18978469.
- ^ Evropský patent [1] (10. prosince 2003) „Použití kódujících mikrosatelitních regionů posune peptidy odvozené od mutací pro léčbu rakoviny“ od Bork et al
Další čtení
- Farabaugh PJ (1996). "Naprogramovaný překladový posun snímků". Annu. Genet. 30 (1): 507–28. doi:10.1146 / annurev.genet.30.1.507. PMC 239420. PMID 8982463.
- Lewis, Ricki (2005). Genetika člověka: koncepty a aplikace (6. vydání). Boston MA: McGraw Hill. s. 227–8. ISBN 978-0-07-111156-0.
- "Nylonase Enzymes". 20. dubna 2004. Citováno 2. června 2009.
externí odkazy
- Posun rámu + mutace v americké národní lékařské knihovně Lékařské předměty (Pletivo)
- Databáze NCBI dbSNP - "centrální úložiště jak pro nukleotidové substituce jedné báze, tak pro polymorfismy s krátkou delecí a inzercí"
- Wise2 - zarovná a protein proti umožnění sekvence DNA posuny rámů a introny
- FastY - porovnat sekvenci DNA s databází proteinových sekvencí, umožňující mezery a posuny rámů
- Cesta - nástroj, který porovnává dva shifthift bílkoviny (zadnípřeklad zásada)
- HGMD - Databáze mutací lidského genomu