RNA-sekv - RNA-Seq

RNA-sekv (pojmenovaný jako zkratka „sekvenování RNA“) je založen na konkrétní technologii sekvenování technika, která používá sekvenování nové generace (NGS) odhalit přítomnost a množství RNA v biologickém vzorku v daném okamžiku a analyzuje neustále se měnící buněčnou přepis.[2][3]
Konkrétně RNA-Seq usnadňuje schopnost dívat se na alternativní genové sestřihové transkripty, post-transkripční úpravy, genová fúze mutace /SNP a změny v genové expresi v průběhu času, nebo rozdíly v genové expresi v různých skupinách nebo léčbě.[4] Kromě transkriptů mRNA může RNA-Seq sledovat různé populace RNA, aby zahrnovala celkovou RNA, malou RNA, jako je miRNA, tRNA, a ribozomální profilování.[5] K určení lze také použít RNA-Seq exon /intron hranice a dříve ověřit nebo změnit anotováno 5' a 3' hranice genů. Mezi nedávné pokroky v RNA-Seq patří sekvenování jedné buňky a in situ sekvenování fixované tkáně.[6]
Před RNA-Seq byly studie genové exprese prováděny na základě hybridizace mikročipy. Problémy s mikročipy zahrnují artefakty křížové hybridizace, špatná kvantifikace nízko a vysoce exprimovaných genů a potřeba znát sekvenci a priori.[7] Kvůli těmto technickým problémům transkriptomika převedeny na metody založené na sekvenování. Ty postupovaly od Sangerovo sekvenování z Expresní značka sekvence knihovny, na metody založené na chemických značkách (např. sériová analýza genové exprese ) a konečně k současné technologii, sekvenování nové generace z cDNA (zejména RNA-Seq).
Metody
Příprava knihovny

Obecné kroky k přípravě a komplementární DNA (cDNA) knihovna pro sekvenování jsou popsány níže, ale často se liší mezi platformami.[8][3][9]
- Izolace RNA: RNA je izolována z tkáně a smíchané s deoxyribonukleáza (DNáza). DNáza snižuje množství genomové DNA. Míra degradace RNA se kontroluje pomocí gel a kapilární elektroforéza a slouží k přiřazení Číslo integrity RNA na vzorek. Tato kvalita RNA a celkové množství výchozí RNA se bere v úvahu během následujících kroků přípravy knihovny, sekvenování a analýzy.
- Výběr / vyčerpání RNA: K analýze sledovaných signálů může být izolovaná RNA buď uchována tak, jak je, zbavena ribozomální RNA (rRNA), filtrováno na RNA pomocí 3 'polyadenylovaný (poly (A)) ocasy pouze zahrnout mRNA a / nebo filtrovány na RNA, která váže specifické sekvence (Metody selekce a deplece RNA tabulka níže). U eukaryot je RNA s 3 'poly (A) ocasy zralé, zpracované, kódující sekvence. Poly (A) selekce se provádí smícháním eukaryotické RNA s poly (T) oligomery kovalentně připojenými k substrátu, typicky magnetickým kuličkám.[10][11] Výběr Poly (A) ignoruje nekódující RNA a zavádí zkreslení 3 ',[12] kterému se při strategii vyčerpání ribozomů vyhneme. RRNA je odstraněna, protože představuje více než 90% RNA v buňce, která, pokud by byla uchována, utopila by další data v transkriptomu.
- cDNA syntéza: RNA je zpětně přepsaný k cDNA, protože DNA je stabilnější a umožňuje amplifikaci (což využívá DNA polymerázy ) a využít vyspělejší technologii sekvenování DNA. Zesílení po reverzní transkripci vede ke ztrátě uvíznutí, kterému se lze vyhnout chemickým značením nebo sekvenováním jedné molekuly. Fragmentace a výběr velikosti se provádí za účelem čištění sekvencí, které jsou vhodné délky pro sekvenční stroj. RNA, cDNA nebo obě jsou fragmentovány enzymy, sonikace nebo nebulizéry. Fragmentace RNA snižuje zkreslení 5 'náhodně aktivované reverzní transkripce a vliv primer vazebná místa,[11] s nevýhodou, že 5 'a 3' konce jsou konvertovány na DNA méně efektivně. Po fragmentaci následuje výběr velikosti, kde jsou odstraněny buď malé sekvence, nebo je vybrán úzký rozsah délek sekvencí. Protože malé RNA mají rádi miRNA jsou ztraceny, jsou analyzovány samostatně. CDNA pro každý experiment může být indexována pomocí hexamerového nebo oktamerového čárového kódu, takže tyto experimenty mohou být sloučeny do jednoho pruhu pro multiplexované sekvenování.
| Strategie | Typ RNA | Obsah ribozomální RNA | Nezpracovaný obsah RNA | Obsah genomové DNA | Izolační metoda |
|---|---|---|---|---|---|
| Celková RNA | Všechno | Vysoký | Vysoký | Vysoký | Žádný |
| Výběr PolyA | Kódování | Nízký | Nízký | Nízký | Hybridizace s poly (dT) oligomery |
| Vyčerpání rRNA | Kódování, nekódování | Nízký | Vysoký | Vysoký | Odstranění oligomerů komplementárních k rRNA |
| Zachycení RNA | Cílené | Nízký | Mírný | Nízký | Hybridizace se sondami doplňujícími požadované transkripty |
Malé sekvenování RNA / nekódující RNA
Při sekvenování jiné RNA než mRNA se modifikuje příprava knihovny. Buněčná RNA je vybrána na základě požadovaného rozsahu velikosti. Pro malé cíle RNA, jako je miRNA, je RNA izolována výběrem velikosti. To lze provést pomocí gelu vylučujícího velikost, pomocí magnetických kuliček pro výběr velikosti nebo pomocí komerčně vyvinuté soupravy. Jakmile jsou izolovány, jsou linkery přidány na 3 'a 5' konec a poté purifikovány. Posledním krokem je cDNA generace prostřednictvím reverzní transkripce.
Přímé sekvenování RNA

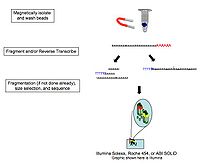
Protože přeměňuje RNA na cDNA Bylo prokázáno, že ligace, amplifikace a další manipulace se vzorky zavádějí předsudky a artefakty, které mohou interferovat se správnou charakterizací i kvantifikací transkriptů,[13] společnosti zahrnující přímé sekvenování jedné molekuly RNA Helicos (úpadce), Oxford Nanopore Technologies,[14] a další. Tato technologie sekvenuje molekuly RNA přímo masivně paralelně.
Sekvenování jednobuněčné RNA (scRNA-Seq)
Standardní metody jako např mikročipy a standardní hromadná analýza RNA-Seq analyzuje expresi RNA z velkých populací buněk. Ve smíšených populacích buněk mohou tato měření zakrýt kritické rozdíly mezi jednotlivými buňkami v těchto populacích.[15][16]
Sekvenování jednobuněčné RNA (scRNA-Seq) poskytuje profily výrazů jednotlivých buněk. Ačkoli není možné získat úplné informace o každé RNA exprimované každou buňkou, vzhledem k malému množství dostupného materiálu lze vzorce genové exprese identifikovat pomocí genu shlukové analýzy. To může odhalit existenci vzácných typů buněk v buněčné populaci, které nikdy předtím nebyly vidět. Například zvané vzácné specializované buňky v plicích plicní ionocyty které vyjadřují Regulátor transmembránové vodivosti cystické fibrózy byly identifikovány v roce 2018 dvěma skupinami provádějícími scRNA-Seq na epitelu dýchacích cest.[17][18]
Experimentální postupy

Současné protokoly scRNA-Seq zahrnují následující kroky: izolace jedné buňky a RNA, reverzní transkripce (RT), amplifikace, generování knihovny a sekvenování. První metody rozdělily jednotlivé buňky do samostatných jamek; novější metody zapouzdřují jednotlivé buňky v kapičkách v mikrofluidním zařízení, kde probíhá reverzní transkripční reakce, převádějící RNA na cDNA. Každá kapička nese DNA "čárový kód", který jedinečně označuje cDNA odvozené z jedné buňky. Jakmile je reverzní transkripce dokončena, mohou být cDNA z mnoha buněk smíchány dohromady pro sekvenování; přepisy z konkrétní buňky jsou identifikovány jedinečným čárovým kódem.[19][20]
Výzvy pro scRNA-Seq zahrnují zachování počátečního relativního množství mRNA v buňce a identifikaci vzácných transkriptů.[21] Krok reverzní transkripce je kritický, protože účinnost RT reakce určuje, kolik populace RNA buňky bude nakonec analyzováno sekvencerem. Procesivita reverzních transkriptáz a použité primární strategie mohou ovlivnit produkci cDNA v plné délce a generování knihoven předpojatých ke 3 'nebo 5' konci genů.
V kroku amplifikace buď PCR nebo in vitro transkripce (IVT) se v současné době používá k amplifikaci cDNA. Jednou z výhod metod založených na PCR je schopnost generovat cDNA plné délky. Různá účinnost PCR na konkrétních sekvencích (například obsah GC a struktura snapbacku) však může být také exponenciálně amplifikována, což vytváří knihovny s nerovnoměrným pokrytím. Na druhou stranu, zatímco knihovny generované IVT se mohou vyhnout zkreslení sekvencí vyvolaných PCR, specifické sekvence mohou být přepsány neefektivně, což způsobí výpadek sekvence nebo generování neúplných sekvencí.[22][15]Bylo publikováno několik protokolů scRNA-Seq: Tang et al.,[23]STRT,[24]SMART-seq,[25]CEL-seq,[26]RAGE-seq,[27], Quartz-seq.[28]a C1-CAGE.[29] Tyto protokoly se liší, pokud jde o strategie pro reverzní transkripci, syntézu a amplifikaci cDNA, a možnost přizpůsobit čárové kódy specifické pro sekvenci (tj. UMI ) nebo schopnost zpracovávat shromážděné vzorky.[30]
V roce 2017 byly zavedeny dva přístupy k současnému měření jednobuněčné exprese mRNA a proteinu prostřednictvím protilátek značených oligonukleotidy známých jako REAP-seq,[31] a CITE-seq.[32]
Aplikace
scRNA-Seq se stává široce používaným napříč biologickými disciplínami včetně vývoje, Neurologie,[33] Onkologie,[34][35][36] Autoimunitní onemocnění,[37] a Infekční nemoc.[38]
scRNA-Seq poskytl značný pohled na vývoj embryí a organismů, včetně červa Caenorhabditis elegans,[39] a regenerativní planar Schmidtea mediterranea.[40][41] První obratlovci, kteří byli takto mapováni, byli Zebrafish[42][43] a Xenopus laevis.[44] V každém případě bylo studováno několik stadií embrya, což umožnilo mapovat celý proces vývoje na bázi buňka po buňce.[8] Věda uznal tyto zálohy jako rok 2018 Průlom roku.[45]
Experimentální úvahy
Různé parametry jsou brány v úvahu při navrhování a provádění experimentů RNA-Seq:
- Specifičnost tkáně: Genová exprese se liší uvnitř a mezi tkáněmi a RNA-Seq měří tuto kombinaci buněčných typů. To může ztěžovat izolaci sledovaného biologického mechanismu. Sekvenování jednotlivých buněk lze použít ke studiu každé buňky samostatně, zmírnění tohoto problému.
- Časová závislost: Genová exprese se časem mění a RNA-Seq pořídí pouze snímek. Pro sledování změn v transkriptomu lze provádět experimenty s časovým průběhem.
- Pokrytí (také známé jako hloubka): RNA obsahuje stejné mutace pozorované v DNA a detekce vyžaduje hlubší pokrytí. S dostatečně vysokým pokrytím lze použít RNA-Seq k odhadu exprese každé alely. To může poskytnout vhled do jevů, jako jsou potisk nebo cis-regulační účinky. Hloubku sekvenování požadovanou pro konkrétní aplikace lze extrapolovat z pilotního experimentu.[46]
- Artefakty generování dat (známé také jako technická odchylka): Činidla (např. Souprava pro přípravu knihovny), zapojený personál a typ sekvenceru (např. Illumina, Pacific Biosciences ) může vést k technickým artefaktům, které mohou být nesprávně interpretovány jako smysluplné výsledky. Stejně jako u jiných vědeckých experimentů je rozumné provádět RNA-Seq v dobře kontrolovaném prostředí. Pokud to není možné nebo je studie a metaanalýza, dalším řešením je detekovat technické artefakty odvozením latentní proměnné (typicky analýza hlavních komponent nebo faktorová analýza ) a následnou korekci těchto proměnných.[47]
- Správa dat: Jeden experiment RNA-Seq u lidí je obvykle v řádu 1 GB.[48] Tento velký objem dat může představovat problémy s úložištěm. Jedno řešení je komprese data pomocí víceúčelových výpočetních schémat (např. gzip ) nebo schémata specifická pro genomiku. Ten může být založen na referenčních sekvencích nebo de novo. Dalším řešením je provádět experimenty s mikročipy, které mohou být dostatečné pro práci založenou na hypotézách nebo replikační studie (na rozdíl od průzkumného výzkumu).
Analýza
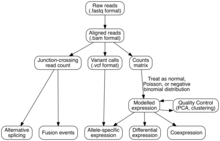
Sestava transkriptomu
K přiřazení nezpracovaných sekvenčních čtení genomickým znakům (tj. Sestavení transkriptomu) se používají dvě metody:
- De novo: Tento přístup nevyžaduje a referenční genom rekonstruovat transkriptom a obvykle se používá, pokud je genom ve srovnání s referencí neznámý, neúplný nebo podstatně pozměněný.[49] Výzvy při použití krátkých čtení pro sestavu de novo zahrnují 1) určení, která čtení by měla být spojena do souvislých sekvencí (kontigy ), 2) robustnost vůči sekvenčním chybám a dalším artefaktům a 3) výpočetní účinnost. Primární algoritmus použitý pro sestavu de novo přešel z překrývajících se grafů, které identifikují všechna párová překrytí mezi čteními, na de Bruijn grafy, který rozdělí čte do sekvencí délky k a sbalí všechny k-mery do hash tabulky.[50] Překrývající se grafy byly použity se Sangerovým sekvenováním, ale neomezují se dobře na miliony čtení generovaných pomocí RNA-Seq. Příklady assemblerů, které používají grafy de Bruijn, jsou Samet,[51] Trojice,[49] Oázy,[52] a Bridger.[53] Spárovaný konec a dlouhé sekvenční čtení stejného vzorku mohou zmírnit deficity v krátkém sekvenčním čtení tím, že slouží jako šablona nebo kostra. Metriky pro hodnocení kvality sestavy de novo zahrnují střední délku kontig, počet kontig a N50.[54]
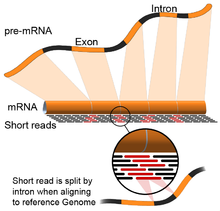
- Vedený genom: Tento přístup se opírá o stejné metody, které se používají pro zarovnání DNA, s další složitostí srovnávání čtení, která pokrývají nekontinuální části referenčního genomu.[55] Tato nekontinuální čtení jsou výsledkem sekvenování sestřižených přepisů (viz obrázek). Algoritmy zarovnání mají obvykle dva kroky: 1) zarovnání krátkých částí čtení (tj. Naočkování genomu) a 2) použití dynamické programování najít optimální zarovnání, někdy v kombinaci se známými anotacemi. Softwarové nástroje, které používají zarovnání vedené genomem, zahrnují Bowtie,[56] TopHat (který staví na výsledcích BowTie k zarovnání spojů spojení),[57][58] Subread,[59] HVĚZDA,[55] HISAT2,[60] Plachetník,[61] Kallisto,[62] a GMAP.[63] Kvalitu sestavy vedené genomem lze měřit jak 1) metrikami sestavy de novo (např. N50), tak 2) srovnáním se známými transkripty, spojovacími spoji, genomem a proteinovými sekvencemi pomocí přesnost, odvolání nebo jejich kombinace (např. skóre F1).[54] Navíc, in silico hodnocení lze provést pomocí simulovaných čtení.[64][65]
Poznámka ke kvalitě montáže: Současná shoda je v tom, že 1) kvalita sestavy se může lišit v závislosti na použité metrice, 2) sestavy, které dosáhly dobrého skóre u jednoho druhu, nemusí nutně dobře fungovat u ostatních druhů a 3) kombinace nejrůznějších přístupů může být nejspolehlivější.[66][67]
Kvantifikace genové exprese
Exprese je kvantifikována pro studium buněčných změn v reakci na vnější podněty, rozdíly mezi zdravými a nemocný státy a další výzkumné otázky. Genová exprese se často používá jako proxy pro hojnost proteinů, ale často nejsou ekvivalentní kvůli post transkripčním událostem, jako je Interference RNA a nesmysl zprostředkovaný úpadek.[68]
Výraz je kvantifikován počítáním počtu čtení, která byla namapována na každý lokus v transkriptomová sestava krok. Exprese může být kvantifikována pro exony nebo geny pomocí kontigů nebo anotací referenčních transkriptů.[8] Tyto pozorované počty čtení RNA-Seq byly důkladně ověřeny oproti starším technologiím, včetně expresních mikročipů a qPCR.[46][69] Příklady nástrojů, které kvantifikují počty, jsou HTSeq,[70] FeatureCounts,[71] Rcount,[72] maximální počet účtů,[73] FIXSEQ,[74] a Cuffquant. Počty čtení jsou poté převedeny do příslušných metrik pro testování hypotéz, regrese a další analýzy. Parametry pro tento převod jsou:
- Hloubka / pokrytí sekvence: I když je hloubka při provádění několika experimentů RNA-Seq předem specifikována, bude se mezi experimenty stále značně lišit.[75] Proto je celkový počet čtení generovaných v jednom experimentu obvykle normalizován převedením počtů na fragmenty, čtení nebo počty na milion mapovaných čtení (FPM, RPM nebo CPM). Hloubka sekvenování se někdy označuje jako velikost knihovny, počet zprostředkujících molekul cDNA v experimentu.
- Délka genu: Delší geny budou mít více fragmentů / čtení / počtů než kratší geny, pokud je transkripční exprese stejná. To je upraveno dělením FPM délkou genu, což vede k metrickým fragmentům na kilobázi přepisu na milion mapovaných čtení (FPKM).[76] Při pohledu na skupiny genů napříč vzorky se FPKM převádí na transkripty na milion (TPM) dělením každého FPKM součtem FPKM ve vzorku.[77][78][79]
- Celkový výstup RNA: Protože se z každého vzorku extrahuje stejné množství RNA, vzorky s více celkovou RNA budou mít méně RNA na gen. Zdá se, že tyto geny mají sníženou expresi, což vede k falešným pozitivům v následných analýzách.[75] Normalizační strategie zahrnující kvantil, DESeq2, TMM a Median Ratio se pokoušejí vysvětlit tento rozdíl porovnáním sady nediferenčně exprimovaných genů mezi vzorky a odpovídajícím měřítkem.[80]
- Rozptyl pro expresi každého genu: je vytvořen tak, aby odpovídal chyba vzorkování (důležité pro geny s nízkým počtem čtení), zvýšit sílu a snížit falešně pozitivní výsledky. Rozptyl lze odhadnout jako a normální, jed nebo negativní binomický rozdělení[81][82][83] a často se rozkládá na technickou a biologickou odchylku.
Absolutní kvantifikace
Absolutní kvantifikace genové exprese není možná u většiny experimentů RNA-Seq, které kvantifikují expresi vzhledem ke všem transkriptům. Je to možné provedením RNA-Seq se špičkami, vzorky RNA při známých koncentracích. Po sekvenování se počty načtení spike-in sekvencí použijí ke stanovení vztahu mezi počty načtení každého genu a absolutním množstvím biologických fragmentů.[11][84] V jednom příkladu byla tato technika použita v Xenopus tropicalis embrya k určení kinetiky transkripce.[85]
Diferenciální výraz
Nejjednodušším, ale často nejsilnějším využitím RNA-Seq je nalezení rozdílů v genové expresi mezi dvěma nebo více podmínkami (např., léčeno vs. neléčeno); tento proces se nazývá diferenciální výraz. Výstupy jsou často označovány jako odlišně exprimované geny (DEG) a tyto geny mohou být buď regulovány nahoru nebo dolů (tj., vyšší nebo nižší v podmínce zájmu). Je jich mnoho nástroje, které provádějí diferenciální výraz. Většina z nich je spuštěna R, Krajta, nebo Unix příkazový řádek. Mezi běžně používané nástroje patří DESeq,[82] edgeR,[83] a voom + limma,[81][86] všechny jsou k dispozici prostřednictvím R /Biovodič.[87][88] Při provádění rozdílového výrazu jsou běžné úvahy:
- Vstupy: Diferenciální vstupy exprese zahrnují (1) RNA-Seq expresní matici (M geny x N vzorky) a (2) a návrhová matice obsahující experimentální podmínky pro N vzorků. Nejjednodušší návrhová matice obsahuje jeden sloupec, který odpovídá štítkům testované podmínky. Mohou zahrnovat i další proměnné (označované také jako faktory, vlastnosti, štítky nebo parametry) dávkové efekty, známé artefakty a veškerá metadata, která by mohla zmást nebo zprostředkovat genovou expresi. Kromě známých kovariát je možné odhadnout také neznámé kovariáty strojové učení bez dozoru přístupy včetně hlavní složka, náhradní proměnná,[89] a PEER[47] analýzy. Skryté analýzy proměnných se často používají pro data RNA lidské sekvence tkáně, která obvykle obsahují další artefakty nezachycené v metadatech (např., ischemický čas, získávání z více institucí, základní klinické vlastnosti, shromažďování údajů po mnoho let s mnoha zaměstnanci).
- Metody: Většina nástrojů používá regrese nebo neparametrické statistiky identifikovat odlišně exprimované geny a jsou buď založené na počtu (DESeq2, limma, edgeR) nebo na základě sestavení (prostřednictvím kvantifikace bez zarovnání, sleuth,[90] Cuffdiff,[91] Plesové šaty[92]).[93] Po regresi většina nástrojů využívá buď rodinná chybovost (FWER) nebo míra falešných objevů (FDR) p-hodnota úpravy na účet více hypotéz (ve studiích na lidech ~ 20 000 genů kódujících proteiny nebo ~ 50 000 biotypů).
- Výstupy: Typický výstup se skládá z řádků odpovídajících počtu genů a alespoň tří sloupců, protokol každého genu složit změnu (log-transformace poměru ve vyjádření mezi podmínkami, míra velikost efektu ), p-hodnota a hodnota p upravená pro více srovnání. Geny jsou definovány jako biologicky smysluplné, pokud projdou mezními hodnotami pro velikost účinku (změna logaritmického záhybu) a statistická významnost. Tyto mezní hodnoty by měly být v ideálním případě specifikovány a priori, ale podstata experimentů RNA-Seq je často průzkumná, takže je obtížné předvídat velikosti účinku a příslušné mezní hodnoty předem.
- Úskalí: Raison d'etre těchto složitých metod je vyhnout se nesčetným nástrahám, které mohou vést statistické chyby a zavádějící interpretace. Mezi úskalí patří zvýšená míra falešně pozitivních výsledků (kvůli vícenásobnému srovnání), artefakty přípravy vzorků, heterogenita vzorků (jako smíšené genetické pozadí), vysoce korelované vzorky, nezohledněné víceúrovňové experimentální návrhy a chudý experimentální design. Jedním pozoruhodným úskalím je prohlížení výsledků v aplikaci Microsoft Excel bez použití funkce importu, která zajistí, že názvy genů zůstanou textem.[94] Ačkoli je to vhodné, Excel automaticky převádí některé názvy genů (SEPT1, DEC1, 2. BŘEZNA ) do dat nebo čísel s plovoucí desetinnou čárkou.
- Výběr nástrojů a benchmarking: Existuje mnoho snah, které srovnávají výsledky těchto nástrojů, přičemž DESeq2 má tendenci mírně překonávat jiné metody.[95][96][97][98][99][93][100] Stejně jako u jiných metod spočívá benchmarking ve vzájemném porovnání výstupů nástroje a známých zlaté standardy.
Následné analýzy seznamu odlišně exprimovaných genů přicházejí ve dvou příchutích, což potvrzuje pozorování a vytváří biologické závěry. Vzhledem k úskalím diferenciální exprese a RNA-Seq jsou důležitá pozorování replikována (1) ortogonální metodou ve stejných vzorcích (jako real-time PCR ) nebo (2) někdy jiný předregistrováno, experimentujte v nové kohortě. Ten pomáhá zajistit zobecnitelnost a lze jej obvykle sledovat metaanalýzou všech sdružených kohort. Nejběžnější metodou pro získání biologického porozumění výsledků na vyšší úrovni je analýza obohacení genové sady, i když se někdy používají přístupy kandidátských genů. Obohatení sady genů určuje, zda je překrytí mezi dvěma sadami genů statisticky významné, v tomto případě překrytí rozdílně exprimovaných genů a sad genů ze známých cest / databází (např., Genová ontologie, KEGG, Lidská fenotypová ontologie ) nebo z doplňkových analýz ve stejných datech (jako jsou sítě pro společné vyjádření). Mezi běžné nástroje pro obohacování genové sady patří webová rozhraní (např., ENRICHR, g: profiler) a softwarové balíčky. Při hodnocení výsledků obohacování je jednou heuristikou nejprve hledat obohacení známé biologie jako kontrolu zdravého rozumu a poté rozšířit rozsah tak, aby hledal novou biologii.

Alternativní sestřih
Sestřih RNA je nedílnou součástí eukaryot a významně přispívá k regulaci a rozmanitosti proteinů, vyskytující se u> 90% lidských genů.[101] Existuje několik alternativní režimy sestřihu: přeskočení exonu (nejběžnější způsob sestřihu u lidí a vyšších eukaryot), vzájemně se vylučující exony, alternativní donorová nebo akceptorová místa, retence intronu (nejběžnější způsob sestřihu u rostlin, hub a prvoků), alternativní místo zahájení transkripce (promotor) a alternativní polyadenylace.[101] Jedním z cílů RNA-Seq je identifikovat alternativní sestřihové události a otestovat, zda se liší mezi podmínkami. Sekvenování s dlouhým čtením zachycuje celý přepis a minimalizuje tak mnoho problémů při odhadování četnosti izoforem, jako je nejednoznačné mapování čtení. Pro krátké čtení RNA-Seq existuje několik metod k detekci alternativního sestřihu, které lze rozdělit do tří hlavních skupin:[102][103][104]
- Na základě počtu (také na základě událostí, diferenciální sestřih): odhad retence exonu. Příklady jsou DEXSeq,[105] KOBERCE,[106] a SeqGSEA.[107]
- Na základě izoformy (také vícečetné moduly, diferenciální výraz izoformy): nejprve odhadněte početnost izoformy a poté relativní hojnost mezi podmínkami. Příkladem jsou manžetové knoflíčky 2[108] a DiffSplice.[109]
- Intronová excize na základě: vypočítat alternativní sestřih pomocí split čtení. Příkladem je MAJIQ[110] a řezačka listí.[104]
Nástroje diferenciální genové exprese lze také použít pro diferenciální expresi izoformy, pokud jsou izoformy kvantifikovány předem s jinými nástroji, jako je RSEM.[111]
Koexpresní sítě
Koexpresní sítě jsou reprezentace dat odvozených od genů chujících se podobným způsobem napříč tkáněmi a experimentálními podmínkami.[112] Jejich hlavní účel spočívá v generování hypotéz a přístupech viny za asociací pro odvození funkcí dříve neznámých genů.[112] Data RNA-Seq byla použita k odvození genů zapojených do specifických drah na základě Pearsonova korelace, a to jak v rostlinách[113] a savci.[114] Hlavní výhodou dat RNA-Seq v tomto druhu analýzy přes platformy microarray je schopnost pokrýt celý transkriptom, což umožňuje možnost odhalit úplnější reprezentace genových regulačních sítí. Diferenciální regulace spojovacích izoforem stejného genu může být detekována a použita k předpovědi a jejich biologickým funkcím.[115][116] Vážená genová koexpresní síťová analýza byl úspěšně použit k identifikaci koexpresních modulů a genů intramodulárního rozbočovače na základě dat RNA seq. Moduly společné exprese mohou odpovídat buněčným typům nebo drahám. Vysoce propojené intramodulární rozbočovače lze interpretovat jako zástupce příslušného modulu. Eigengen je vážený součet exprese všech genů v modulu. Eigengeny jsou užitečné biomarkery (vlastnosti) pro diagnostiku a prognózu.[117] Byly navrženy přístupy transformace stabilizující rozptyl pro odhad korelačních koeficientů na základě dat RNA seq.[113]
Objev variant
RNA-Seq zachycuje variace DNA, včetně jednotlivé nukleotidové varianty, malá vložení / vymazání. a strukturální variace. Varianta volání v RNA-Seq je podobný volání varianty DNA a často používá stejné nástroje (včetně SAMtools mpileup[118] a GATK HaplotypeCaller[119]) s úpravami, které zohledňují spojování. Jedna jedinečná dimenze pro varianty RNA je alelově specifický výraz (ASE): varianty pouze z jednoho haplotypu mohou být přednostně vyjádřeny v důsledku regulačních účinků včetně potisk a exprese kvantitativní znak loci a nekódující vzácné varianty.[120][121] Omezení identifikace varianty RNA zahrnují, že odráží pouze exprimované oblasti (u lidí <5% genomu) a má nižší kvalitu ve srovnání s přímým sekvenováním DNA.
Editace RNA (post-transkripční změny)
Mít odpovídající genomové a transkriptomické sekvence jednotlivce může pomoci detekovat post-transkripční úpravy (Úpravy RNA ).[3] Událost post-transkripční modifikace je identifikována, pokud má transkript genu alelu / variantu, která nebyla v genomových datech pozorována.
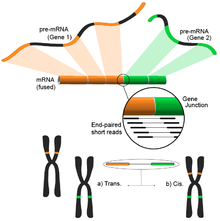
Detekce fúzního genu
Fúzní geny, způsobené různými strukturálními modifikacemi v genomu, získaly pozornost kvůli jejich vztahu k rakovině.[122] Schopnost RNA-Seq analyzovat celý transkriptom vzorku nezaujatým způsobem z něj činí atraktivní nástroj k nalezení těchto druhů běžných událostí u rakoviny.[4]
Myšlenka vyplývá z procesu srovnávání krátkých transkriptomických čtení s referenčním genomem. Většina krátkých čtení spadá do jednoho úplného exonu a očekává se, že menší, ale stále velká sada bude mapována na známé křižovatky exon-exon. Zbývající nezmapovaná krátká čtení by pak byla dále analyzována, aby se zjistilo, zda odpovídají spojení exon-exon, kde exony pocházejí z různých genů. To by byl důkaz možné události fúze, ale vzhledem k délce čtení by se to mohlo ukázat jako velmi hlučné. Alternativním přístupem je použití čtení na párových koncích, kdy by potenciálně velký počet spárovaných čtení mapoval každý konec na jiný exon, což by poskytlo lepší pokrytí těchto událostí (viz obrázek). Konečný výsledek nicméně sestává z mnoha a potenciálně nových kombinací genů poskytujících ideální výchozí bod pro další validaci.
Dějiny

RNA-Seq byl poprvé vyvinut v polovině 2000s s příchodem technologie sekvenování nové generace.[123] První rukopisy, které používaly RNA-Seq i bez použití tohoto termínu, zahrnují rukopisy rakovina prostaty buněčné linie[124] (ze dne 2006), Medicago truncatula[125] (2006), kukuřice[126] (2007) a Arabidopsis thaliana[127] (2007), zatímco samotný termín „RNA-Seq“ byl poprvé zmíněn v roce 2008.[128] Počet rukopisů odkazujících na RNA-Seq v názvu nebo abstraktu (obrázek, modrá čára) se neustále zvyšuje s 6754 rukopisy publikovanými v roce 2018 (odkaz na vyhledávání PubMed ). Průsečík RNA-Seq a medicíny (obrázek, zlatá čára, odkaz na vyhledávání PubMed ) má podobnou celeritu.[původní výzkum? ]
Aplikace v medicíně
RNA-Seq má potenciál identifikovat novou biologii nemocí, profilovat biomarkery pro klinické indikace, odvodit léčebné cesty a provádět genetické diagnózy. Tyto výsledky by mohly být dále přizpůsobeny pro podskupiny nebo dokonce pro jednotlivé pacienty, což by mohlo zdůraznit účinnější prevenci, diagnostiku a terapii. Proveditelnost tohoto přístupu je částečně dána náklady v penězích a čase; souvisejícím omezením je požadovaný tým specialistů (bioinformatici, lékaři / kliničtí lékaři, základní výzkumní pracovníci, technici), aby plně interpretovali obrovské množství dat generovaných touto analýzou.[129]
Velké úsilí v oblasti sekvenování
Velký důraz byl kladen na data RNA-Seq po Encyklopedie prvků DNA (ENCODE) a Atlas rakovinového genomu (TCGA) projekty tento přístup využily k charakterizaci desítek buněčných linií[130] a tisíce vzorků primárních nádorů,[131] resp. ENCODE zaměřený na identifikaci regulačních oblastí v celém genomu v různých kohortách buněčných linií a transkriptomické údaje jsou zásadní, aby bylo možné pochopit následný účinek těchto epigenetických a genetických regulačních vrstev. TCGA se místo toho zaměřila na shromáždění a analýzu tisíců vzorků pacientů od 30 různých typů nádorů, aby pochopila základní mechanismy maligní transformace a progrese. V této souvislosti poskytují data RNA-Seq jedinečný snímek transkriptomického stavu onemocnění a zkoumají nestrannou populaci transkriptů, která umožňuje identifikaci nových transkriptů, fúzních transkriptů a nekódujících RNA, které by nebylo možné detekovat různými technologiemi.
Viz také
Reference
- ^ Shafee T, Lowe R (2017). "Eukaryotická a prokaryotická genová struktura". WikiJournal of Medicine. 4 (1). doi:10.15347 / wjm / 2017.002.
- ^ Chu Y, Corey DR (srpen 2012). „Sekvenování RNA: výběr platformy, experimentální design a interpretace dat“. Nukleová kyselina Therapeutics. 22 (4): 271–4. doi:10.1089 / nat.2012.0367. PMC 3426205. PMID 22830413.
- ^ A b C Wang Z, Gerstein M, Snyder M (leden 2009). „RNA-Seq: revoluční nástroj pro transkriptomiku“. Recenze přírody. Genetika. 10 (1): 57–63. doi:10.1038 / nrg2484. PMC 2949280. PMID 19015660.
- ^ A b Maher CA, Kumar-Sinha C, Cao X, Kalyana-Sundaram S, Han B, Jing X a kol. (Březen 2009). "Transkriptomové sekvenování k detekci genových fúzí u rakoviny". Příroda. 458 (7234): 97–101. Bibcode:2009Natur.458 ... 97M. doi:10.1038 / nature07638. PMC 2725402. PMID 19136943.
- ^ Ingolia NT, Brar GA, Rouskin S, McGeachy AM, Weissman JS (červenec 2012). „Strategie profilování ribozomu pro monitorování translace in vivo hlubokým sekvenováním fragmentů mRNA chráněných ribozomy“. Přírodní protokoly. 7 (8): 1534–50. doi:10.1038 / nprot.2012.086. PMC 3535016. PMID 22836135.
- ^ Lee JH, Daugharthy ER, Scheiman J, Kalhor R, Yang JL, Ferrante TC a kol. (Březen 2014). "Vysoce multiplexované subcelulární RNA sekvenování in situ". Věda. 343 (6177): 1360–3. Bibcode:2014Sci ... 343.1360L. doi:10.1126 / science.1250212. PMC 4140943. PMID 24578530.
- ^ Kukurba KR, Montgomery SB (duben 2015). "Sekvenování a analýza RNA". Cold Spring Harbor Protocols. 2015 (11): 951–69. doi:10.1101 / pdb.top084970. PMC 4863231. PMID 25870306.
- ^ A b C d E Griffith M, Walker JR, Spies NC, Ainscough BJ, Griffith OL (srpen 2015). „Informatics for RNA Sequencing: a Web Resource for Analysis on the cloud“. PLOS výpočetní biologie. 11 (8): e1004393. Bibcode:2015PLSCB..11E4393G. doi:10.1371 / journal.pcbi.1004393. PMC 4527835. PMID 26248053.
- ^ "RNA-seqlopedia". rnaseq.uoregon.edu. Citováno 2017-02-08.
- ^ Morin R, Bainbridge M, Fejes A, Hirst M, Krzywinski M, Pugh T a kol. (Červenec 2008). „Profilování transkriptomu HeLa S3 pomocí náhodně aktivované cDNA a masivně paralelního sekvenování krátkého čtení“. Biotechniky. 45 (1): 81–94. doi:10.2144/000112900. PMID 18611170.
- ^ A b C Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B (červenec 2008). "Mapování a kvantifikace savčích transkriptomů pomocí RNA-Seq". Přírodní metody. 5 (7): 621–8. doi:10.1038 / nmeth.1226. PMID 18516045. S2CID 205418589.
- ^ Chen EA, Souaiaia T, Herstein JS, Evgrafov OV, Spitsyna VN, Rebolini DF, Knowles JA (říjen 2014). "Vliv integrity RNA na jedinečně mapovaná čtení v RNA-Seq". Poznámky k výzkumu BMC. 7 (1): 753. doi:10.1186/1756-0500-7-753. PMC 4213542. PMID 25339126.
- ^ Liu D, Graber JH (únor 2006). „Kvantitativní srovnání knihoven EST vyžaduje kompenzaci za systematické předsudky při generování cDNA“. BMC bioinformatika. 7: 77. doi:10.1186/1471-2105-7-77. PMC 1431573. PMID 16503995.
- ^ Garalde DR, Snell EA, Jachimowicz D, Sipos B, Lloyd JH, Bruce M a kol. (Březen 2018). "Vysoce paralelní přímé sekvenování RNA na poli nanopórů". Přírodní metody. 15 (3): 201–206. doi:10.1038 / nmeth.4577. PMID 29334379. S2CID 3589823.
- ^ A b "Shapiro E, Biezuner T, Linnarsson S (září 2013). „Technologie založené na sekvenování jednotlivých buněk způsobí revoluci ve vědě celého organismu.“ Recenze přírody. Genetika. 14 (9): 618–30. doi:10.1038 / nrg3542. PMID 23897237. S2CID 500845."
- ^ Kolodziejczyk AA, Kim JK, Svensson V, Marioni JC, Teichmann SA (květen 2015). „Technologie a biologie sekvenování jednobuněčné RNA“. Molekulární buňka. 58 (4): 610–20. doi:10.1016 / j.molcel.2015.04.005. PMID 26000846.
- ^ Montoro DT, Haber AL, Biton M, Vinarsky V, Lin B, Birket SE, et al. (Srpen 2018). "A revised airway epithelial hierarchy includes CFTR-expressing ionocytes". Příroda. 560 (7718): 319–324. Bibcode:2018Natur.560..319M. doi:10.1038/s41586-018-0393-7. PMC 6295155. PMID 30069044.
- ^ Plasschaert LW, Žilionis R, Choo-Wing R, Savova V, Knehr J, Roma G, et al. (Srpen 2018). "A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte". Příroda. 560 (7718): 377–381. Bibcode:2018Natur.560..377P. doi:10.1038/s41586-018-0394-6. PMC 6108322. PMID 30069046.
- ^ Klein AM, Mazutis L, Akartuna I, Tallapragada N, Veres A, Li V a kol. (Květen 2015). „Čárové kódy kapiček pro transkriptomika jedné buňky aplikované na embryonální kmenové buňky“. Buňka. 161 (5): 1187–1201. doi:10.1016 / j.cell.2015.04.044. PMC 4441768. PMID 26000487.
- ^ Macosko EZ, Basu A, Satija R, Nemesh J, Shekhar K, Goldman M, et al. (Květen 2015). "Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets". Buňka. 161 (5): 1202–1214. doi:10.1016/j.cell.2015.05.002. PMC 4481139. PMID 26000488.
- ^ "Hebenstreit D (November 2012). "Methods, Challenges and Potentials of Single Cell RNA-seq". Biologie. 1 (3): 658–67. doi:10.3390/biology1030658. PMC 4009822. PMID 24832513."
- ^ Eberwine J, Sul JY, Bartfai T, Kim J (January 2014). "The promise of single-cell sequencing". Přírodní metody. 11 (1): 25–7. doi:10.1038/nmeth.2769. PMID 24524134. S2CID 11575439.
- ^ Tang F, Barbacioru C, Wang Y, Nordman E, Lee C, Xu N a kol. (Květen 2009). msgstr "Analýza celého transkriptomu mRNA-Seq jedné buňky". Přírodní metody. 6 (5): 377–82. doi:10.1038/NMETH.1315. PMID 19349980. S2CID 16570747.
- ^ Islam S, Kjällquist U, Moliner A, Zajac P, Fan JB, Lönnerberg P, Linnarsson S (July 2011). "Characterization of the single-cell transcriptional landscape by highly multiplex RNA-seq". Výzkum genomu. 21 (7): 1160–7. doi:10.1101/gr.110882.110. PMC 3129258. PMID 21543516.
- ^ Ramsköld D, Luo S, Wang YC, Li R, Deng Q, Faridani OR, et al. (Srpen 2012). "Full-length mRNA-Seq from single-cell levels of RNA and individual circulating tumor cells". Přírodní biotechnologie. 30 (8): 777–82. doi:10.1038/nbt.2282. PMC 3467340. PMID 22820318.
- ^ Hashimshony T, Wagner F, Sher N, Yanai I (září 2012). „CEL-Seq: jednobuněčná RNA-Seq multiplexovanou lineární amplifikací“. Zprávy buněk. 2 (3): 666–73. doi:10.1016 / j.celrep.2012.08.003. PMID 22939981.
- ^ Singh M, Al-Eryani G, Carswell S, Ferguson JM, Blackburn J, Barton K, Roden D, Luciani F, Phan T, Junankar S, Jackson K, Goodnow CC, Smith MA, Swarbrick A (2018). „Vysoce výkonné cílené dlouho čitelné sekvenování jedné buňky odhaluje klonální a transkripční krajinu lymfocytů“. bioRxiv. doi:10.1101/424945. PMID 31311926.
- ^ Sasagawa Y, Nikaido I, Hayashi T, Danno H, Uno KD, Imai T, Ueda HR (duben 2013). „Quartz-Seq: vysoce reprodukovatelná a citlivá metoda jednobuněčného sekvenování RNA, odhaluje genetickou genetickou heterogenitu“. Genome Biology. 14 (4): R31. doi:10.1186 / gb-2013-14-4-r31. PMC 4054835. PMID 23594475.
- ^ Kouno T, Moody J, Kwon AT, Shibayama Y, Kato S, Huang Y a kol. (Leden 2019). „C1 CAGE detekuje počáteční místa transkripce a aktivitu zesilovače při rozlišení jedné buňky“. Příroda komunikace. 10 (1): 360. Bibcode:2019NatCo..10..360K. doi:10.1038 / s41467-018-08126-5. PMC 6341120. PMID 30664627.
- ^ Dal Molin A, Di Camillo B (2019). "How to design a single-cell RNA-sequencing experiment: pitfalls, challenges and perspectives". Briefings in Bioinformatics. 20 (4): 1384–1394. doi:10.1093/bib/bby007. PMID 29394315.
- ^ Peterson VM, Zhang KX, Kumar N, Wong J, Li L, Wilson DC, et al. (Říjen 2017). "Multiplexed quantification of proteins and transcripts in single cells". Přírodní biotechnologie. 35 (10): 936–939. doi:10.1038/nbt.3973. PMID 28854175. S2CID 205285357.
- ^ Stoeckius M, Hafemeister C, Stephenson W, Houck-Loomis B, Chattopadhyay PK, Swerdlow H a kol. (Září 2017). „Simultánní měření epitopu a transkriptomu v jednotlivých buňkách“. Přírodní metody. 14 (9): 865–868. doi:10.1038 / nmeth.4380. PMC 5669064. PMID 28759029.
- ^ Raj B, Wagner DE, McKenna A, Pandey S, Klein AM, Shendure J a kol. (Červen 2018). „Simultánní jednobuněčný profil linií a typů buněk v mozku obratlovců“. Přírodní biotechnologie. 36 (5): 442–450. doi:10.1038 / nbt.4103. PMC 5938111. PMID 29608178.
- ^ Olmos D, Arkenau HT, Ang JE, Ledaki I, Attard G, Carden CP, et al. (Leden 2009). "Circulating tumour cell (CTC) counts as intermediate end points in castration-resistant prostate cancer (CRPC): a single-centre experience". Annals of Oncology. 20 (1): 27–33. doi:10.1093/annonc/mdn544. PMID 18695026.
- ^ Levitin HM, Yuan J, Sims PA (April 2018). "Single-Cell Transcriptomic Analysis of Tumor Heterogeneity". Trendy v rakovině. 4 (4): 264–268. doi:10.1016/j.trecan.2018.02.003. PMC 5993208. PMID 29606308.
- ^ Jerby-Arnon L, Shah P, Cuoco MS, Rodman C, Su MJ, Melms JC, et al. (Listopad 2018). "A Cancer Cell Program Promotes T Cell Exclusion and Resistance to Checkpoint Blockade". Buňka. 175 (4): 984–997.e24. doi:10.1016/j.cell.2018.09.006. PMC 6410377. PMID 30388455.
- ^ Stephenson W, Donlin LT, Butler A, Rozo C, Bracken B, Rashidfarrokhi A, et al. (Únor 2018). "Single-cell RNA-seq of rheumatoid arthritis synovial tissue using low-cost microfluidic instrumentation". Příroda komunikace. 9 (1): 791. Bibcode:2018NatCo...9..791S. doi:10.1038/s41467-017-02659-x. PMC 5824814. PMID 29476078.
- ^ Avraham R, Haseley N, Brown D, Penaranda C, Jijon HB, Trombetta JJ, et al. (Září 2015). "Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses". Buňka. 162 (6): 1309–21. doi:10.1016/j.cell.2015.08.027. PMC 4578813. PMID 26343579.
- ^ Cao J, Packer JS, Ramani V, Cusanovich DA, Huynh C, Daza R a kol. (August 2017). „Komplexní transkripční profilování mnohobuněčného organismu v jedné buňce“. Věda. 357 (6352): 661–667. Bibcode:2017Sci ... 357..661C. doi:10.1126 / science.aam8940. PMC 5894354. PMID 28818938.
- ^ Plass M, Solana J, Wolf FA, Ayoub S, Misios A, Glažar P, et al. (Květen 2018). "Cell type atlas and lineage tree of a whole complex animal by single-cell transcriptomics". Věda. 360 (6391): eaaq1723. doi:10.1126/science.aaq1723. PMID 29674432.
- ^ Fincher CT, Wurtzel O, de Hoog T, Kravarik KM, Reddien PW (May 2018). "Schmidtea mediterranea". Věda. 360 (6391): eaaq1736. doi:10.1126/science.aaq1736. PMC 6563842. PMID 29674431.
- ^ Wagner DE, Weinreb C, Collins ZM, Briggs JA, Megason SG, Klein AM (červen 2018). „Jednobuněčné mapování krajiny a linie genové exprese v embryu zebrafish“. Věda. 360 (6392): 981–987. Bibcode:2018Sci ... 360..981W. doi:10.1126 / science.aar4362. PMC 6083445. PMID 29700229.
- ^ Farrell JA, Wang Y, Riesenfeld SJ, Shekhar K, Regev A, Schier AF (June 2018). "Single-cell reconstruction of developmental trajectories during zebrafish embryogenesis". Věda. 360 (6392): eaar3131. doi:10.1126/science.aar3131. PMC 6247916. PMID 29700225.
- ^ Briggs JA, Weinreb C, Wagner DE, Megason S, Peshkin L, Kirschner MW, Klein AM (June 2018). "The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution". Věda. 360 (6392): eaar5780. doi:10.1126/science.aar5780. PMC 6038144. PMID 29700227.
- ^ You J. "Science's 2018 Breakthrough of the Year: tracking development cell by cell". Vědecký časopis. Americká asociace pro rozvoj vědy.
- ^ A b Li H, Lovci MT, Kwon YS, Rosenfeld MG, Fu XD, Yeo GW (December 2008). "Determination of tag density required for digital transcriptome analysis: application to an androgen-sensitive prostate cancer model". Sborník Národní akademie věd Spojených států amerických. 105 (51): 20179–84. Bibcode:2008PNAS..10520179L. doi:10.1073/pnas.0807121105. PMC 2603435. PMID 19088194.
- ^ A b Stegle O, Parts L, Piipari M, Winn J, Durbin R (February 2012). "Using probabilistic estimation of expression residuals (PEER) to obtain increased power and interpretability of gene expression analyses". Přírodní protokoly. 7 (3): 500–7. doi:10.1038/nprot.2011.457. PMC 3398141. PMID 22343431.
- ^ Kingsford C, Patro R (June 2015). "Reference-based compression of short-read sequences using path encoding". Bioinformatika. 31 (12): 1920–8. doi:10.1093/bioinformatics/btv071. PMC 4481695. PMID 25649622.
- ^ A b Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, et al. (Květen 2011). „Kompletní transkriptomová sestava z dat RNA-Seq bez referenčního genomu“. Přírodní biotechnologie. 29 (7): 644–52. doi:10.1038 / nbt.1883. PMC 3571712. PMID 21572440.
- ^ "De Novo Assembly Using Illumina Reads" (PDF). Citováno 22. října 2016.
- ^ Zerbino DR, Birney E (May 2008). "Velvet: algoritmy pro de novo sestavení krátkého čtení pomocí grafů de Bruijn". Výzkum genomu. 18 (5): 821–9. doi:10.1101 / gr.074492.107. PMC 2336801. PMID 18349386.
- ^ Oases: a transcriptome assembler for very short reads
- ^ Chang Z, Li G, Liu J, Zhang Y, Ashby C, Liu D, et al. (Únor 2015). "Bridger: a new framework for de novo transcriptome assembly using RNA-seq data". Genome Biology. 16 (1): 30. doi:10.1186/s13059-015-0596-2. PMC 4342890. PMID 25723335.
- ^ A b Li B, Fillmore N, Bai Y, Collins M, Thomson JA, Stewart R, Dewey CN (December 2014). "Evaluation of de novo transcriptome assemblies from RNA-Seq data". Genome Biology. 15 (12): 553. doi:10.1186/s13059-014-0553-5. PMC 4298084. PMID 25608678.
- ^ A b Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, et al. (Leden 2013). "STAR: ultrafast universal RNA-seq aligner". Bioinformatika. 29 (1): 15–21. doi:10.1093/bioinformatics/bts635. PMC 3530905. PMID 23104886.
- ^ Langmead B, Trapnell C, Pop M, Salzberg SL (2009). "Ultrafast and memory-efficient alignment of short DNA sequences to the human genome". Genome Biology. 10 (3): R25. doi:10.1186/gb-2009-10-3-r25. PMC 2690996. PMID 19261174.
- ^ Trapnell C, Pachter L, Salzberg SL (May 2009). "TopHat: discovering splice junctions with RNA-Seq". Bioinformatika. 25 (9): 1105–11. doi:10.1093/bioinformatics/btp120. PMC 2672628. PMID 19289445.
- ^ Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, et al. (Březen 2012). "Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks". Přírodní protokoly. 7 (3): 562–78. doi:10.1038/nprot.2012.016. PMC 3334321. PMID 22383036.
- ^ Liao Y, Smyth GK, Shi W (May 2013). "The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote". Výzkum nukleových kyselin. 41 (10): e108. doi:10.1093/nar/gkt214. PMC 3664803. PMID 23558742.
- ^ Kim D, Langmead B, Salzberg SL (April 2015). "HISAT: a fast spliced aligner with low memory requirements". Přírodní metody. 12 (4): 357–60. doi:10.1038/nmeth.3317. PMC 4655817. PMID 25751142.
- ^ Patro R, Mount SM, Kingsford C (May 2014). „Sailfish umožňuje kvantifikaci izoformy bez zarovnání ze čtení RNA-sek pomocí lehkých algoritmů“. Přírodní biotechnologie. 32 (5): 462–4. arXiv:1308.3700. doi:10.1038 / nbt.2862. PMC 4077321. PMID 24752080.
- ^ Bray NL, Pimentel H, Melsted P, Pachter L (May 2016). "Near-optimal probabilistic RNA-seq quantification". Přírodní biotechnologie. 34 (5): 525–7. doi:10.1038/nbt.3519. PMID 27043002. S2CID 205282743.
- ^ Wu TD, Watanabe CK (May 2005). "GMAP: a genomic mapping and alignment program for mRNA and EST sequences". Bioinformatika. 21 (9): 1859–75. doi:10.1093/bioinformatics/bti310. PMID 15728110.
- ^ Baruzzo G, Hayer KE, Kim EJ, Di Camillo B, FitzGerald GA, Grant GR (February 2017). "Simulation-based comprehensive benchmarking of RNA-seq aligners". Přírodní metody. 14 (2): 135–139. doi:10.1038/nmeth.4106. PMC 5792058. PMID 27941783.
- ^ Engström PG, Steijger T, Sipos B, Grant GR, Kahles A, Rätsch G, et al. (December 2013). "Systematic evaluation of spliced alignment programs for RNA-seq data". Přírodní metody. 10 (12): 1185–91. doi:10.1038/nmeth.2722. PMC 4018468. PMID 24185836.
- ^ Lu B, Zeng Z, Shi T (February 2013). "Comparative study of de novo assembly and genome-guided assembly strategies for transcriptome reconstruction based on RNA-Seq". Science China Life Sciences. 56 (2): 143–55. doi:10.1007/s11427-013-4442-z. PMID 23393030.
- ^ Bradnam KR, Fass JN, Alexandrov A, Baranay P, Bechner M, Birol I, et al. (Červenec 2013). „Assemblathon 2: hodnocení de novo metod montáže genomu u tří druhů obratlovců“. GigaScience. 2 (1): 10. arXiv:1301.5406. Bibcode:2013arXiv1301.5406B. doi:10.1186 / 2047-217X-2-10. PMC 3844414. PMID 23870653.
- ^ Greenbaum D, Colangelo C, Williams K, Gerstein M (2003). "Comparing protein abundance and mRNA expression levels on a genomic scale". Genome Biology. 4 (9): 117. doi:10.1186/gb-2003-4-9-117. PMC 193646. PMID 12952525.
- ^ Zhang ZH, Jhaveri DJ, Marshall VM, Bauer DC, Edson J, Narayanan RK, et al. (Srpen 2014). "A comparative study of techniques for differential expression analysis on RNA-Seq data". PLOS ONE. 9 (8): e103207. Bibcode:2014PLoSO...9j3207Z. doi:10.1371/journal.pone.0103207. PMC 4132098. PMID 25119138.
- ^ Anders S, Pyl PT, Huber W (January 2015). "HTSeq--a Python framework to work with high-throughput sequencing data". Bioinformatika. 31 (2): 166–9. doi:10.1093/bioinformatics/btu638. PMC 4287950. PMID 25260700.
- ^ Liao Y, Smyth GK, Shi W (April 2014). "featureCounts: an efficient general purpose program for assigning sequence reads to genomic features". Bioinformatika. 30 (7): 923–30. arXiv:1305.3347. doi:10.1093/bioinformatics/btt656. PMID 24227677. S2CID 15960459.
- ^ Schmid MW, Grossniklaus U (February 2015). "Rcount: simple and flexible RNA-Seq read counting". Bioinformatika. 31 (3): 436–7. doi:10.1093/bioinformatics/btu680. PMID 25322836.
- ^ Finotello F, Lavezzo E, Bianco L, Barzon L, Mazzon P, Fontana P, et al. (2014). "Reducing bias in RNA sequencing data: a novel approach to compute counts". BMC bioinformatika. 15 Suppl 1 (Suppl 1): S7. doi:10.1186/1471-2105-15-s1-s7. PMC 4016203. PMID 24564404.
- ^ Hashimoto TB, Edwards MD, Gifford DK (March 2014). "Universal count correction for high-throughput sequencing". PLOS výpočetní biologie. 10 (3): e1003494. Bibcode:2014PLSCB..10E3494H. doi:10.1371/journal.pcbi.1003494. PMC 3945112. PMID 24603409.
- ^ A b Robinson MD, Oshlack A (2010). "A scaling normalization method for differential expression analysis of RNA-seq data". Genome Biology. 11 (3): R25. doi:10.1186/gb-2010-11-3-r25. PMC 2864565. PMID 20196867.
- ^ Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, et al. (Květen 2010). "Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation". Přírodní biotechnologie. 28 (5): 511–5. doi:10.1038/nbt.1621. PMC 3146043. PMID 20436464.
- ^ Pachter L (19 April 2011). "Models for transcript quantification from RNA-Seq". arXiv:1104.3889 [q-bio.GN ].
- ^ "What the FPKM? A review of RNA-Seq expression units". The farrago. 8. května 2014. Citováno 28. března 2018.
- ^ Wagner GP, Kin K, Lynch VJ (December 2012). "Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples". Theory in Biosciences = Theorie in den Biowissenschaften. 131 (4): 281–5. doi:10.1007/s12064-012-0162-3. PMID 22872506. S2CID 16752581.
- ^ Evans, Ciaran; Hardin, Johanna; Stoebel, Daniel M (28 September 2018). "Selecting between-sample RNA-Seq normalization methods from the perspective of their assumptions". Briefings in Bioinformatics. 19 (5): 776–792. doi:10.1093/bib/bbx008. PMC 6171491. PMID 28334202.
- ^ A b Law CW, Chen Y, Shi W, Smyth GK (February 2014). "voom: Precision weights unlock linear model analysis tools for RNA-seq read counts". Genome Biology. 15 (2): R29. doi:10.1186/gb-2014-15-2-r29. PMC 4053721. PMID 24485249.
- ^ A b Anders S, Huber W (2010). "Diferenciální analýza výrazu pro data o počtu sekvencí". Genome Biology. 11 (10): R106. doi:10.1186 / gb-2010-11-10-r106. PMC 3218662. PMID 20979621.
- ^ A b Robinson MD, McCarthy DJ, Smyth GK (January 2010). „edgeR: balíček Bioconductor pro diferenciální analýzu exprese dat digitální genové exprese“. Bioinformatika. 26 (1): 139–40. doi:10.1093 / bioinformatika / btp616. PMC 2796818. PMID 19910308.
- ^ Marguerat S, Schmidt A, Codlin S, Chen W, Aebersold R, Bähler J (October 2012). "Quantitative analysis of fission yeast transcriptomes and proteomes in proliferating and quiescent cells". Buňka. 151 (3): 671–83. doi:10.1016/j.cell.2012.09.019. PMC 3482660. PMID 23101633.
- ^ Owens ND, Blitz IL, Lane MA, Patrushev I, Overton JD, Gilchrist MJ, et al. (Leden 2016). "Measuring Absolute RNA Copy Numbers at High Temporal Resolution Reveals Transcriptome Kinetics in Development". Zprávy buněk. 14 (3): 632–647. doi:10.1016/j.celrep.2015.12.050. PMC 4731879. PMID 26774488.
- ^ Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, Smyth GK (April 2015). "limma powers differential expression analyses for RNA-sequencing and microarray studies". Výzkum nukleových kyselin. 43 (7): e47. doi:10.1093/nar/gkv007. PMC 4402510. PMID 25605792.
- ^ "Bioconductor - Open source software for bioinformatics".
- ^ Huber W, Carey VJ, Gentleman R, Anders S, Carlson M, Carvalho BS, et al. (Únor 2015). „Organizace vysoce výkonné genomické analýzy pomocí Bioconductor“. Přírodní metody. 12 (2): 115–21. doi:10.1038 / nmeth.3252. PMC 4509590. PMID 25633503.
- ^ Leek JT, Storey JD (September 2007). "Capturing heterogeneity in gene expression studies by surrogate variable analysis". Genetika PLOS. 3 (9): 1724–35. doi:10.1371/journal.pgen.0030161. PMC 1994707. PMID 17907809.
- ^ Pimentel H, Bray NL, Puente S, Melsted P, Pachter L (July 2017). "Differential analysis of RNA-seq incorporating quantification uncertainty". Přírodní metody. 14 (7): 687–690. doi:10.1038/nmeth.4324. PMID 28581496. S2CID 15063247.
- ^ Trapnell C, Hendrickson DG, Sauvageau M, Goff L, Rinn JL, Pachter L (January 2013). "Differential analysis of gene regulation at transcript resolution with RNA-seq" (PDF). Přírodní biotechnologie. 31 (1): 46–53. doi:10.1038/nbt.2450. PMC 3869392. PMID 23222703.
- ^ Frazee AC, Pertea G, Jaffe AE, Langmead B, Salzberg SL, Leek JT (March 2015). "Ballgown bridges the gap between transcriptome assembly and expression analysis". Přírodní biotechnologie. 33 (3): 243–6. doi:10.1038/nbt.3172. PMC 4792117. PMID 25748911.
- ^ A b Sahraeian SM, Mohiyuddin M, Sebra R, Tilgner H, Afshar PT, Au KF, et al. (Červenec 2017). "Gaining comprehensive biological insight into the transcriptome by performing a broad-spectrum RNA-seq analysis". Příroda komunikace. 8 (1): 59. Bibcode:2017NatCo...8...59S. doi:10.1038/s41467-017-00050-4. PMC 5498581. PMID 28680106.
- ^ Ziemann M, Eren Y, El-Osta A (srpen 2016). „Chyby ve jménech genů jsou ve vědecké literatuře rozšířené“. Genome Biology. 17 (1): 177. doi:10.1186 / s13059-016-1044-7. PMC 4994289. PMID 27552985.
- ^ Soneson C, Delorenzi M (March 2013). "A comparison of methods for differential expression analysis of RNA-seq data". BMC bioinformatika. 14: 91. doi:10.1186/1471-2105-14-91. PMC 3608160. PMID 23497356.
- ^ Fonseca NA, Marioni J, Brazma A (30 September 2014). "RNA-Seq gene profiling--a systematic empirical comparison". PLOS ONE. 9 (9): e107026. Bibcode:2014PLoSO...9j7026F. doi:10.1371/journal.pone.0107026. PMC 4182317. PMID 25268973.
- ^ Seyednasrollah F, Laiho A, Elo LL (January 2015). "Comparison of software packages for detecting differential expression in RNA-seq studies". Briefings in Bioinformatics. 16 (1): 59–70. doi:10.1093/bib/bbt086. PMC 4293378. PMID 24300110.
- ^ Rapaport F, Khanin R, Liang Y, Pirun M, Krek A, Zumbo P, et al. (2013). "Comprehensive evaluation of differential gene expression analysis methods for RNA-seq data". Genome Biology. 14 (9): R95. doi:10.1186/gb-2013-14-9-r95. PMC 4054597. PMID 24020486.
- ^ Conesa A, Madrigal P, Tarazona S, Gomez-Cabrero D, Cervera A, McPherson A, et al. (Leden 2016). „Průzkum osvědčených postupů pro analýzu dat RNA-seq“. Genome Biology. 17 (1): 13. doi:10.1186 / s13059-016-0881-8. PMC 4728800. PMID 26813401.
- ^ Costa-Silva J, Domingues D, Lopes FM (21 December 2017). "RNA-Seq differential expression analysis: An extended review and a software tool". PLOS ONE. 12 (12): e0190152. Bibcode:2017PLoSO..1290152C. doi:10.1371/journal.pone.0190152. PMC 5739479. PMID 29267363.
- ^ A b Keren H, Lev-Maor G, Ast G (May 2010). "Alternative splicing and evolution: diversification, exon definition and function". Recenze přírody. Genetika. 11 (5): 345–55. doi:10.1038/nrg2776. PMID 20376054. S2CID 5184582.
- ^ Liu R, Loraine AE, Dickerson JA (December 2014). "Comparisons of computational methods for differential alternative splicing detection using RNA-seq in plant systems". BMC bioinformatika. 15 (1): 364. doi:10.1186/s12859-014-0364-4. PMC 4271460. PMID 25511303.
- ^ Pachter, Lior (19 April 2011). "Models for transcript quantification from RNA-Seq". arXiv:1104.3889 [q-bio.GN ].
- ^ A b Li YI, Knowles DA, Humphrey J, Barbeira AN, Dickinson SP, Im HK, Pritchard JK (January 2018). "Annotation-free quantification of RNA splicing using LeafCutter". Genetika přírody. 50 (1): 151–158. doi:10.1038/s41588-017-0004-9. PMC 5742080. PMID 29229983.
- ^ Anders S, Reyes A, Huber W (October 2012). "Detecting differential usage of exons from RNA-seq data". Výzkum genomu. 22 (10): 2008–17. doi:10.1101/gr.133744.111. PMC 3460195. PMID 22722343.
- ^ Shen S, Park JW, Huang J, Dittmar KA, Lu ZX, Zhou Q, et al. (April 2012). "MATS: a Bayesian framework for flexible detection of differential alternative splicing from RNA-Seq data". Výzkum nukleových kyselin. 40 (8): e61. doi:10.1093/nar/gkr1291. PMC 3333886. PMID 22266656.
- ^ Wang X, Cairns MJ (June 2014). "SeqGSEA: a Bioconductor package for gene set enrichment analysis of RNA-Seq data integrating differential expression and splicing". Bioinformatika. 30 (12): 1777–9. doi:10.1093/bioinformatics/btu090. PMID 24535097.
- ^ Trapnell C, Hendrickson DG, Sauvageau M, Goff L, Rinn JL, Pachter L (January 2013). "Differential analysis of gene regulation at transcript resolution with RNA-seq". Přírodní biotechnologie. 31 (1): 46–53. doi:10.1038/nbt.2450. PMC 3869392. PMID 23222703.
- ^ Hu Y, Huang Y, Du Y, Orellana CF, Singh D, Johnson AR, et al. (Leden 2013). "DiffSplice: the genome-wide detection of differential splicing events with RNA-seq". Výzkum nukleových kyselin. 41 (2): e39. doi:10.1093/nar/gks1026. PMC 3553996. PMID 23155066.
- ^ Vaquero-Garcia J, Barrera A, Gazzara MR, González-Vallinas J, Lahens NF, Hogenesch JB, et al. (Únor 2016). "A new view of transcriptome complexity and regulation through the lens of local splicing variations". eLife. 5: e11752. doi:10.7554/eLife.11752. PMC 4801060. PMID 26829591.
- ^ Merino GA, Conesa A, Fernández EA (March 2019). "A benchmarking of workflows for detecting differential splicing and differential expression at isoform level in human RNA-seq studies". Briefings in Bioinformatics. 20 (2): 471–481. doi:10.1093/bib/bbx122. PMID 29040385. S2CID 22706028.
- ^ A b Marcotte EM, Pellegrini M, Thompson MJ, Yeates TO, Eisenberg D (November 1999). "A combined algorithm for genome-wide prediction of protein function". Příroda. 402 (6757): 83–6. Bibcode:1999Natur.402...83M. doi:10.1038/47048. PMID 10573421. S2CID 144447.
- ^ A b Giorgi FM, Del Fabbro C, Licausi F (March 2013). "Comparative study of RNA-seq- and microarray-derived coexpression networks in Arabidopsis thaliana". Bioinformatika. 29 (6): 717–24. doi:10.1093/bioinformatics/btt053. PMID 23376351.
- ^ Iancu OD, Kawane S, Bottomly D, Searles R, Hitzemann R, McWeeney S (June 2012). "Utilizing RNA-Seq data for de novo coexpression network inference". Bioinformatika. 28 (12): 1592–7. doi:10.1093/bioinformatics/bts245. PMC 3493127. PMID 22556371.
- ^ Eksi R, Li HD, Menon R, Wen Y, Omenn GS, Kretzler M, Guan Y (Nov 2013). "Systematically differentiating functions for alternatively spliced isoforms through integrating RNA-seq data". PLOS výpočetní biologie. 9 (11): e1003314. Bibcode:2013PLSCB...9E3314E. doi:10.1371/journal.pcbi.1003314. PMC 3820534. PMID 24244129.
- ^ Li HD, Menon R, Omenn GS, Guan Y (August 2014). "The emerging era of genomic data integration for analyzing splice isoform function". Trendy v genetice. 30 (8): 340–7. doi:10.1016/j.tig.2014.05.005. PMC 4112133. PMID 24951248.
- ^ Foroushani A, Agrahari R, Docking R, Chang L, Duns G, Hudoba M, et al. (Březen 2017). "Large-scale gene network analysis reveals the significance of extracellular matrix pathway and homeobox genes in acute myeloid leukemia: an introduction to the Pigengene package and its applications". BMC Medical Genomics. 10 (1): 16. doi:10.1186/s12920-017-0253-6. PMC 5353782. PMID 28298217.
- ^ Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. (August 2009). "The Sequence Alignment/Map format and SAMtools". Bioinformatika. 25 (16): 2078–9. doi:10.1093/bioinformatics/btp352. PMC 2723002. PMID 19505943.
- ^ DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C, et al. (Květen 2011). "A framework for variation discovery and genotyping using next-generation DNA sequencing data". Genetika přírody. 43 (5): 491–8. doi:10.1038/ng.806. PMC 3083463. PMID 21478889.
- ^ Battle A, Brown CD, Engelhardt BE, Montgomery SB (October 2017). "Genetic effects on gene expression across human tissues". Příroda. 550 (7675): 204–213. Bibcode:2017Natur.550..204A. doi:10.1038/nature24277. PMC 5776756. PMID 29022597.
- ^ Richter F, Hoffman GE, Manheimer KB, Patel N, Sharp AJ, McKean D, et al. (Březen 2019). "ORE Identifies Extreme Expression Effects Enriched for Rare Variants". Bioinformatika. 35 (20): 3906–3912. doi:10.1093/bioinformatics/btz202. PMC 6792115. PMID 30903145.
- ^ Teixeira MR (prosinec 2006). "Opakované fúzní onkogeny v karcinomech". Kritické recenze v onkogenezi. 12 (3–4): 257–71. doi:10.1615 / critrevoncog.v12.i3-4,40. PMID 17425505.
- ^ Weber AP (November 2015). "Discovering New Biology through Sequencing of RNA". Fyziologie rostlin. 169 (3): 1524–31. doi:10.1104/pp.15.01081. PMC 4634082. PMID 26353759.
- ^ Bainbridge MN, Warren RL, Hirst M, Romanuik T, Zeng T, Go A, et al. (Září 2006). "Analysis of the prostate cancer cell line LNCaP transcriptome using a sequencing-by-synthesis approach". BMC Genomics. 7: 246. doi:10.1186/1471-2164-7-246. PMC 1592491. PMID 17010196.
- ^ Cheung F, Haas BJ, Goldberg SM, May GD, Xiao Y, Town CD (October 2006). "Sequencing Medicago truncatula expressed sequenced tags using 454 Life Sciences technology". BMC Genomics. 7: 272. doi:10.1186/1471-2164-7-272. PMC 1635983. PMID 17062153.
- ^ Emrich SJ, Barbazuk WB, Li L, Schnable PS (January 2007). "Gene discovery and annotation using LCM-454 transcriptome sequencing". Výzkum genomu. 17 (1): 69–73. doi:10.1101/gr.5145806. PMC 1716268. PMID 17095711.
- ^ Weber AP, Weber KL, Carr K, Wilkerson C, Ohlrogge JB (May 2007). "Sampling the Arabidopsis transcriptome with massively parallel pyrosequencing". Fyziologie rostlin. 144 (1): 32–42. doi:10.1104/pp.107.096677. PMC 1913805. PMID 17351049.
- ^ Nagalakshmi U, Wang Z, Waern K, Shou C, Raha D, Gerstein M, Snyder M (June 2008). "The transcriptional landscape of the yeast genome defined by RNA sequencing". Věda. 320 (5881): 1344–9. Bibcode:2008Sci...320.1344N. doi:10.1126/science.1158441. PMC 2951732. PMID 18451266.
- ^ Sandberg, Rickard (2013-12-30). "Entering the era of single-cell transcriptomics in biology and medicine". Přírodní metody. 11 (1): 22–24. doi:10.1038/nmeth.2764. ISSN 1548-7091.
- ^ "ENCODE Data Matrix". Citováno 2013-07-28.
- ^ "The Cancer Genome Atlas - Data Portal". Citováno 2013-07-28.
externí odkazy
| Scholia má téma profil pro RNA-sekv. |
- RNA-Seq for Everyone: a high-level guide to designing and implementing an RNA-Seq experiment.
- Taguchi, Y.-h. (2019). "Comparative Transcriptomics Analysis". Encyklopedie bioinformatiky a výpočetní biologie. pp. 814–818. doi:10.1016/B978-0-12-809633-8.20163-5. ISBN 9780128114322.
- Referenční modul v biologických vědách