Oxoguanin glykosyláza - Oxoguanine glycosylase - Wikipedia







| 8-oxoguanin DNA glykosyláza, N-terminální doména | |||||||||
|---|---|---|---|---|---|---|---|---|---|
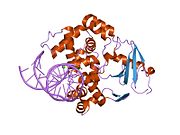
 struktura katalyticky neaktivní q315a lidské 8-oxoguanin glykosylázy v komplexu s 8-oxoguaninem dna | |||||||||
| Identifikátory | |||||||||
| Symbol | OGG_N | ||||||||
| Pfam | PF07934 | ||||||||
| Pfam klan | CL0407 | ||||||||
| InterPro | IPR012904 | ||||||||
| SCOP2 | 1ebm / Rozsah / SUPFAM | ||||||||
| |||||||||
8-oxoguanin glykosyláza také známý jako OGG1 je DNA glykosyláza enzym, který je u lidí kódován OGG1 gen. Je zapojen do oprava základní excize. Nachází se v bakteriální, archaeal a eukaryotický druh.
Funkce
OGG1 je primární enzym odpovědný za excizi 8-oxoguanin (8-oxoG), vedlejší produkt mutagenní báze, ke kterému dochází v důsledku expozice reaktivní formy kyslíku (ROS). OGG1 je bifunkční glykosyláza, protože je schopna štěpit glykosidickou vazbu mutagenní léze a způsobit zlom řetězce v páteři DNA. Alternativní sestřih C-koncové oblasti tohoto genu klasifikuje sestřihové varianty do dvou hlavních skupin, typu 1 a typu 2, v závislosti na posledním exonu sekvence. Alternativní varianty sestřihu typu 1 končí exonem 7 a typ 2 končí exonem 8. Jedna sada sestřižených forem je označena 1a, lb, 2a až 2e.[5] Všechny varianty mají společnou N-koncovou oblast. Bylo popsáno mnoho alternativních sestřihových variant pro tento gen, ale pro každou variantu nebyla stanovena povaha celé délky. U eukaryot obsahuje N-konec tohoto genu mitochondriální cílový signál, nezbytný pro mitochondriální lokalizaci.[6] OGG1-1a však také má na svém C-terminálním konci signál o jaderné poloze, který potlačuje mitochondriální cílení a způsobuje lokalizaci OGG1-1a do jádra.[5] Hlavní formou OGG1, která se lokalizuje do mitochondrií, je OGG1-2a.[5] A konzervovaný N-terminál doména přispívá zbytky k 8-oxoguaninu vazba kapsa. Tato doména je uspořádána do jedné kopie souboru TBP -jako složit.[7]
Navzdory předpokládané důležitosti tohoto enzymu byly generovány myši bez Ogg1 a bylo zjištěno, že mají normální životnost,[8] a Ogg1 knockout myši mají vyšší pravděpodobnost vzniku rakoviny, zatímco narušení genu Mth1 současně potlačuje vývoj rakoviny plic u Ogg1 - / - myší.[9] Ukázalo se, že myši postrádající Ogg1 jsou náchylné ke zvýšení tělesné hmotnosti a obezitě, stejně jako k inzulínové rezistenci vyvolané stravou s vysokým obsahem tuku.[10] Existuje diskuse o tom, zda delece Ogg1 ve skutečnosti vede ke zvýšeným hladinám 8-oxo-dG: analýza HPLC-EC navrhuje až 6krát vyšší hladiny 8-oxo-dG v jaderné DNA a 20krát vyšší v mitochondriální DNA zatímco stanovení Fapy-glykosylázy nenaznačuje žádnou změnu.[Citace je zapotřebí ]
OGG1 nedostatek a zvýšil 8-oxo-dG u myší

Myši bez funkčnosti OGG1 Gen má asi 5krát vyšší hladinu 8-oxo-dG v játrech ve srovnání s myší divokého typu OGG1.[9] Myši vadné OGG1 mají také zvýšené riziko rakoviny.[9] Kunisada a kol.[12] ozářené myši bez funkce OGG1 gen (myši OGG1 knock-out) a myši divokého typu třikrát týdně po dobu čtyřiceti týdnů s UVB světlem v relativně nízké dávce (nestačí k tomu, aby způsobily zarudnutí kůže). Oba typy myší měly vysokou hladinu 8-oxo-dG ve svých epidermálních buňkách tři hodiny po ozáření. Avšak o 24 hodin později chyběla většina 8-oxo-dG v epidermálních buňkách myší divokého typu, ale 8-oxo-dG zůstal zvýšený v epidermálních buňkách OGG1 knock-out myši. Ozářené knock-out myši OGG1 měly více než dvojnásobnou hladinu kožních nádorů ve srovnání s ozářenými myšmi divokého typu a míra malignity v nádorech byla vyšší u myší s knock-outem OGG1 (73%) než u divokých myší. myši typu (50%).
Jak přezkoumali Valavanidis et al.,[13] zvýšené hladiny 8-oxo-dG v tkáni mohou sloužit jako biomarker oxidačního stresu. Rovněž poznamenali, že během karcinogeneze se často vyskytují zvýšené hladiny 8-oxo-dG.
Na obrázku zobrazujícím příklady myšího epitelu tlustého střeva bylo zjištěno, že epitel tlustého střeva u myši na normální stravě má nízkou hladinu 8-oxo-dG ve svých kryptách tlustého střeva (panel A). Myš však pravděpodobně prochází tumorigenezí tlustého střeva (kvůli deoxycholát přidal do své stravy[11]) bylo zjištěno, že má vysokou hladinu 8-oxo-dG v epitelu tlustého střeva (panel B). Deoxycholát zvyšuje intracelulární produkci reaktivního kyslíku, což vede ke zvýšenému oxidačnímu stresu,[14]>[15] a to může vést k tumorigenezi a karcinogenezi.
Epigenetická kontrola
Ve studii rakoviny prsu byla úroveň methylace OGG1 Bylo zjištěno, že promotor antikoreluje s úrovní exprese OGG1 messenger RNA.[16] To znamená, že hypermethylace byla spojena s nízkou expresí OGG1 a hypomethylace korelovala s nadměrnou expresí OGG1. Tím pádem, OGG1 výraz je pod epigenetický řízení. Rakoviny prsu s úrovní methylace OGG1 promotor, které byly více než dvě standardní odchylky buď nad nebo pod normální hodnotou, byly spojeny se sníženým přežitím pacientů.[16]
U rakoviny
OGG1 je primární enzym odpovědný za excizi 8-oxo-2'-deoxyguanosin (8-oxo-dG). I když je exprese OGG1 normální, přítomnost 8-oxo-dG je mutagenní, protože OGG1 není 100% efektivní. Yasui a kol.[17] zkoumal osud 8-oxo-dG, když tento oxidovaný derivát deoxyguanosin byl vložen do specifického genu v 800 buňkách v kultuře. Po replikaci buněk byl 8-oxo-dG obnoven na G v 86% klonů, což pravděpodobně odráží přesný OGG1 oprava základní excize nebo syntéza překladů bez mutace. G: C až T: A transversions došlo v 5,9% klonů, jedna báze vypuštění v 2,1% a G: C až C: G transverze v 1,2%. Společně byly tyto mutace nejčastější, celkem 9,2% ze 14% mutací generovaných v místě inzerce 8-oxo-dG. Mezi dalšími mutacemi v 800 analyzovaných klonech byly také 3 větší delece o velikosti 6, 33 a 135 párů bází. 8-oxo-dG tedy může přímo způsobit mutace, k nimž některé mohou přispívat karcinogeneze.
Li OGG1 exprese je snížena v buňkách, zvýšená mutageneze, a proto zvýšená karcinogeneze by se dalo očekávat. V tabulce níže jsou uvedeny druhy rakoviny se sníženou expresí OGG1.
| Rakovina | Výraz | Forma OGG1 | 8-oxo-dG | Metoda hodnocení | Čj. |
|---|---|---|---|---|---|
| Rakovina hlavy a krku | Nedostatečný výraz | OGG1-2a | - | messenger RNA | [18] |
| Adenokarcinom žaludeční kardie | Nedostatečný výraz | cytoplazmatický | zvýšil | imunohistochemie | [19] |
| Astrocytom | Nedostatečný výraz | celková buňka OGG1 | - | messenger RNA | [20] |
| Rakovina jícnu | 48% nedostatečné vyjádření | jaderný | zvýšil | imunohistochemie | [21] |
| - | 40% nedostatečný výraz | cytoplazma | zvýšil | imunohistochemie | [21] |
Aktivita OGG1 nebo OGG v krvi a rakovina
OGG1 hladiny methylace v krevních buňkách byly měřeny v prospektivní studii 582 amerických veteránů, střední věk 72 let, a sledovány po dobu 13 let. Vysoký OGG1 methylace v konkrétní promotorové oblasti byla spojena se zvýšeným rizikem jakékoli rakoviny a zejména s rizikem rakoviny prostaty.[22]
Enzymatická aktivita excise 8-oxoguanin z DNA (Aktivita OGG) byl snížen v mononukleární buňky periferní krve (PBMC) a ve spárované plicní tkáni od pacientů s nemalobuněčný karcinom plic.[23] Aktivita OGG byla také snížena v PBMC pacientů s spinocelulární karcinom hlavy a krku (HNSCC).[24]
Interakce
Bylo prokázáno, že oxoguanin glykosyláza komunikovat s XRCC1[25] a PKC alfa.[26]
Patologie
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000114026 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000030271 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b C Nishioka K, Ohtsubo T, Oda H, Fujiwara T, Kang D, Sugimachi K, Nakabeppu Y (květen 1999). „Exprese a diferenciální intracelulární lokalizace dvou hlavních forem lidské 8-oxoguaninové DNA glykosylázy kódovaných alternativně sestřiženými OGG1 mRNA“. Molekulární biologie buňky. 10 (5): 1637–1652. doi:10,1091 / mbc. 10.5.1637. PMC 30487. PMID 10233168.
- ^ „Entrez Gene: OGG1 8-oxoguanin DNA glycosylase“.
- ^ Bjørås M, Seeberg E, Luna L, Pearl LH, Barrett TE (březen 2002). „Reciproční„ převrácení „je základem rozpoznání substrátu a katalytické aktivace lidskou 8-oxo-guaninovou DNA glykosylázou“. Journal of Molecular Biology. 317 (2): 171–177. doi:10.1006 / jmbi.2002.5400. PMID 11902834.
- ^ Klungland A, Rosewell I, Hollenbach S, Larsen E, Daly G, Epe B, Seeberg E, Lindahl T, Barnes DE (listopad 1999). "Akumulace premutagenních lézí DNA u myší vadná při odstraňování poškození oxidační bází". Sborník Národní akademie věd Spojených států amerických. 96 (23): 13300–13305. Bibcode:1999PNAS ... 9613300K. doi:10.1073 / pnas.96.23.13300. PMC 23942. PMID 10557315.
- ^ A b C Sakumi K, Tominaga Y, Furuichi M, Xu P, Tsuzuki T, Sekiguchi M, Nakabeppu Y (březen 2003). „Ogg1 knockout-spojená plicní tumorigeneze a její potlačení narušením genu Mth1“. Výzkum rakoviny. 63 (5): 902–905. PMID 12615700.
- ^ Sampath H, Vartanian V, Rollins MR, Sakumi K, Nakabeppu Y, Lloyd RS (prosinec 2012). „Nedostatek 8-oxoguanin DNA glykosylázy (OGG1) zvyšuje náchylnost k obezitě a metabolické dysfunkci“. PLOS ONE. 7 (12): e51697. Bibcode:2012PLoSO ... 751697S. doi:10.1371 / journal.pone.0051697. PMC 3524114. PMID 23284747.
- ^ A b Prasad AR, Prasad S, Nguyen H, Facista A, Lewis C, Zaitlin B, Bernstein H, Bernstein C (červenec 2014). „Nový myší model rakoviny tlustého střeva související se stravou je obdobou lidské rakoviny tlustého střeva“. World Journal of Gastrointestinal Oncology. 6 (7): 225–243. doi:10,4251 / wjgo.v6.i7.225. PMC 4092339. PMID 25024814.
- ^ Kunisada M, Sakumi K, Tominaga Y, Budiyanto A, Ueda M, Ichihashi M, Nakabeppu Y, Nishigori C (červenec 2005). „Tvorba 8-oxoguaninu vyvolaná chronickou expozicí UVB činí myši Ogg1 knockout náchylné na karcinogenezi kůže“. Výzkum rakoviny. 65 (14): 6006–6010. doi:10.1158 / 0008-5472.CAN-05-0724. PMID 16024598.
- ^ Valavanidis A, Vlachogianni T, Fiotakis K, Loridas S (srpen 2013). „Plicní oxidační stres, zánět a rakovina: dýchatelné částice, vláknité prachy a ozon jako hlavní příčiny karcinogeneze plic prostřednictvím mechanismů reaktivních forem kyslíku“. International Journal of Environmental Research and Public Health. 10 (9): 3886–3907. doi:10,3390 / ijerph10093886. PMC 3799517. PMID 23985773.
- ^ Tsuei J, Chau T, Mills D, Wan YJ (listopad 2014). „Dysregulace žlučových kyselin, dysbióza střev a rakovina zažívacího traktu“. Experimentální biologie a medicína. 239 (11): 1489–1504. doi:10.1177/1535370214538743. PMC 4357421. PMID 24951470.
- ^ Ajouz H, Mukherji D, Shamseddine A (květen 2014). „Sekundární žlučové kyseliny: nedostatečně uznávaná příčina rakoviny tlustého střeva“. World Journal of Surgical Oncology. 12: 164. doi:10.1186/1477-7819-12-164. PMC 4041630. PMID 24884764.
- ^ A b Fleischer T, Edvardsen H, Solvang HK, Daviaud C, Naume B, Børresen-Dale AL, Kristensen VN, Tost J (červen 2014). „Integrovaná analýza profilů methylace DNA s vysokým rozlišením, genové exprese, genotypů zárodečné linie a klinických koncových bodů u pacientů s rakovinou prsu“. International Journal of Cancer. 134 (11): 2615–2625. doi:10.1002 / ijc.28606. PMID 24395279. S2CID 32537522.
- ^ Yasui M, Kanemaru Y, Kamoshita N, Suzuki T, Arakawa T, Honma M (březen 2014). „Sledování osudů místně specificky zavedených DNA aduktů v lidském genomu“. Oprava DNA. 15: 11–20. doi:10.1016 / j.dnarep.2014.01.003. PMID 24559511.
- ^ Mahjabeen I, Kayani MA (2016). „Ztráta exprese genů potlačujících mitochondriální nádory je spojena s nepříznivými klinickými následky u karcinomu dlaždicových buněk hlavy a krku: data z retrospektivní studie“. PLOS ONE. 11 (1): e0146948. Bibcode:2016PLoSO..1146948M. doi:10.1371 / journal.pone.0146948. PMC 4718451. PMID 26785117.
- ^ Kohno Y, Yamamoto H, Hirahashi M, Kumagae Y, Nakamura M, Oki E, Oda Y (červen 2016). "Snížená exprese MUTYH, MTH1 a OGG1 a mutace TP53 v adenokarcinomu difúzního typu žaludeční kardie". Lidská patologie. 52: 145–152. doi:10.1016 / j.humpath.2016.01.006. PMID 26980051.
- ^ Jiang Z, Hu J, Li X, Jiang Y, Zhou W, Lu D (prosinec 2006). "Expresní analýzy 27 genů pro opravu DNA v astrocytomu pomocí pole TaqMan s nízkou hustotou". Neurovědy Dopisy. 409 (2): 112–117. doi:10.1016 / j.neulet.2006.09.038. PMID 17034947.
- ^ A b Kubo N, Morita M, Nakashima Y, Kitao H, Egashira A, Saeki H, Oki E, Kakeji Y, Oda Y, Maehara Y (duben 2014). „Oxidační poškození DNA u lidské rakoviny jícnu: klinicko-patologická analýza 8-hydroxydeoxyguanosinu a jeho opravného enzymu“. Nemoci jícnu. 27 (3): 285–293. doi:10,1111 / bod 12,107. hdl:2324/1441070. PMID 23902537.
- ^ Gao T, Joyce BT, Liu L, Zheng Y, Dai Q, Zhang Z, Zhang W, Shrubsole MJ, Tao MH, Schwartz J, Baccarelli A, Hou L (2016). „Methylace DNA genů pro oxidační stres a riziko rakoviny ve studii normativního stárnutí“. American Journal of Cancer Research. 6 (2): 553–561. PMC 4859680. PMID 27186424.
- ^ Paz-Elizur T, Krupsky M, Blumenstein S, Elinger D, Schechtman E, Livneh Z (září 2003). „Oprava DNA pro oxidační poškození a riziko rakoviny plic“. Journal of the National Cancer Institute. 95 (17): 1312–1319. doi:10.1093 / jnci / djg033. PMID 12953085.
- ^ Paz-Elizur T, Ben-Yosef R, Elinger D, Vexler A, Krupsky M, Berrebi A, Shani A, Schechtman E, Freedman L, Livneh Z (prosinec 2006). „Snížená oprava oxidačního poškození 8-oxoguaninu DNA a riziko rakoviny hlavy a krku“. Výzkum rakoviny. 66 (24): 11683–11689. doi:10.1158 / 0008-5472.CAN-06-2294. PMID 17178863.
- ^ Marsin S, Vidal AE, Sossou M, Ménissier-de Murcia J, Le Page F, Boiteux S, de Murcia G, Radicella JP (listopad 2003). „Role XRCC1 v koordinaci a stimulaci opravy poškození oxidační DNA iniciované DNA glykosylázou hOGG1“. The Journal of Biological Chemistry. 278 (45): 44068–44074. doi:10,1074 / jbc.M306160200. PMID 12933815.
- ^ Dantzer F, Luna L, Bjørås M, Seeberg E (červen 2002). „Lidský OGG1 prochází fosforylací serinu a asociuje se s jadernou matricí a mitotickým chromatinem in vivo“. Výzkum nukleových kyselin. 30 (11): 2349–2357. doi:10.1093 / nar / 30.11.2349. PMC 117190. PMID 12034821.
- ^ Osorio A, Milne RL, Kuchenbaecker K, Vaclová T, Pita G, Alonso R, et al. (Duben 2014). „DNA glykosylázy zapojené do opravy excize báze mohou být spojeny s rizikem rakoviny u nosičů mutací BRCA1 a BRCA2“. Genetika PLOS. 10 (4): e1004256. doi:10.1371 / journal.pgen.1004256. PMC 3974638. PMID 24698998.
Další čtení
- Boiteux S, Radicella JP (květen 2000). "Lidský gen OGG1: struktura, funkce a jeho implikace v procesu karcinogeneze". Archivy biochemie a biofyziky. 377 (1): 1–8. doi:10.1006 / abbi.2000.1773. PMID 10775435.
- Park J, Chen L, Tockman MS, Elahi A, Lazarus P (únor 2004). „Lidský 8-oxoguanin DNA N-glykosyláza 1 (hOGG1) DNA opravný enzym a jeho souvislost s rizikem rakoviny plic“. Farmakogenetika. 14 (2): 103–109. doi:10.1097/00008571-200402000-00004. PMID 15077011.
- Hung RJ, Hall J, Brennan P, Boffetta P (listopad 2005). „Genetické polymorfismy v dráze opravy základní excize a riziko rakoviny: recenze HuGE“. American Journal of Epidemiology. 162 (10): 925–942. doi:10.1093 / aje / kwi318. PMID 16221808.
- Mirbahai L, Kershaw RM, Green RM, Hayden RE, Meldrum RA, Hodges NJ (únor 2010). "Použití molekulárního majáku ke sledování aktivity základního excitačního opravného proteinu OGG1 v živých buňkách". Oprava DNA. 9 (2): 144–152. doi:10.1016 / j.dnarep.2009.11.009. PMID 20042377.
- Wang R, Hao W, Pan L, Boldogh I, Ba X (říjen 2018). "Role základního excitačního opravného enzymu OGG1 v genové expresi". Buněčné a molekulární biologické vědy. 75 (20): 3741–3750. doi:10.1007 / s00018-018-2887-8. PMC 6154017. PMID 30043138.
- Vlahopoulos S, Adamaki M, Khoury N, Zoumpourlis V, Boldogh I (2018). "Role DNA opravného enzymu OGG1 ve vrozené imunitě a jeho význam pro rakovinu plic". Farmakologie a terapeutika. 194: 59–72. doi:10.1016 / j.pharmthera.2018.09.004. PMC 6504182. PMID 30240635.
externí odkazy
- oxoguanin + glykosyláza + 1, + člověk v americké národní lékařské knihovně Lékařské předměty (Pletivo)
Galerie PDB | |
|---|---|
|