Nigrostriatální cesta - Nigrostriatal pathway
| Nigrostriatální cesta | |
|---|---|
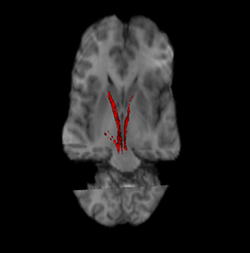 Nigrostriatální cesta (červená a levá a pravá). | |
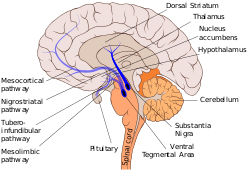 Nigrostriatální dráha je zde zobrazena modrou barvou a spojuje substantia nigra s dorzálním striatem. | |
| Anatomická terminologie |
The nigrostriatální dráha je bilaterální dopaminergní dráha v mozku, který spojuje substantia nigra pars compacta (SNc) ve středním mozku s hřbetní striatum (tj kádové jádro a putamen ) v předním mozku. Je to jeden ze čtyř hlavních dopaminové dráhy v mozku a je zásadní při produkci pohybu jako součásti systému zvaného motorická smyčka bazálních ganglií. Dopaminergní neurony této dráhy uvolňují dopamin z axonových terminálů, na které se synchronizuje GABAergický středně ostnaté neurony (MSN), také známé jako neurony s ostnatou projekcí (SPN),[1][2] umístěný ve striatu.
Degenerace dopaminergních látek neurony v SNc je jedním z hlavních patologických rysů Parkinsonova choroba,[3] což vede k výraznému snížení funkce dopaminu a symptomatickým motorickým deficitům Parkinsonova choroba počítaje v to hypokineze, třes, tuhost a posturální nerovnováha.
Anatomie
Spojení mezi substantia nigra pars compacta a dorzálním striatem je zprostředkováno dopaminergními axony.
Substantia nigra pars compacta (SNc)
The substantia nigra se nachází ve ventrální oblasti střední mozek každé hemisféry. Má dvě odlišné části, pars compacta (SNc) a pars reticulata (SNr). Pars compacta obsahuje dopaminergní neurony z A9 buněčná skupina, která tvoří nigrostriatální cestu, která dodáváním dopaminu do striatum, předává informace do bazální ganglia. Naproti tomu pars reticulata obsahuje většinou GABAergní neurony.
SNc se skládá z tenkého pásu buněk, který překrývá SNr a je umístěn bočně k A10 skupina dopaminergních neuronů v ventrální tegmentální oblast (VTA), která tvoří mezolimbická dopaminová cesta. SNc je snadno vizualizovatelný v sekcích lidského mozku, protože dopaminové neurony obsahují černý pigment zvaný neuromelanin o kterém je známo, že se hromadí s věkem.[4] Těla dopaminergních buněk v SNc jsou hustě zabalena přibližně 200 000 až 420 000 dopaminovými buňkami v lidském SNc a 8 000 až 12 000 dopaminovými buňkami v myším SNc.[5] Tato těla dopaminových buněk jsou lokalizována do jedné ze dvou chemicky definovaných vrstev.[6] Ti v horní vrstvě (nebo hřbetní vrstva) obsahují vazebný protein zvaný calbindin -D28K který může vyrovnávací paměť vápník hladiny uvnitř buňky, když je příliš vysoká nebo toxická. Dopaminové buňky ve spodní vrstvě (nebo ventrální vrstva) postrádá tento protein a je citlivější na účinky neurotoxinů (např. MPTP ), které mohou způsobit příznaky podobné Parkinsonově chorobě.[7][8] Dopaminové buňky dorzální úrovně mají dendrity které vyzařují horizontálně přes pars compacta, zatímco dopaminové buňky ventrální úrovně mají dendrity, které se ventrálně rozšiřují do pars reticulata.[6][9]
Dopaminergní axony
The axony z dopaminových neuronů vycházejí z primárního dendritu a projektu ipsilaterálně (na stejné straně) přes mediální svazek předního mozku do hřbetního striata. Existuje hrubá topografická korelace mezi anatomickou lokalizací těla dopaminových buněk v SNc a oblastí zakončení v dorzálním striatu. Dopaminergní buňky v postranních částech SNc projektu převážně do laterálních a kaudálních (zadních) částí striata, zatímco dopaminové buňky v mediálním SNc projektu do mediálního striata.[10][9] Navíc dopaminové buňky v dorzální vrstvě vyčnívají do ventromediálního striata, zatímco neurony ventrální vrstvy vyčnívají do dorzálního caudate jádra a putamenu.[6][9] Obecně existuje větší hustota dopaminergního vstupu do dorsolaterálního striata.[9]
Každý dopaminový neuron má extrémně velkou nemyelinizovanou axonální arborizaci, která může inervovat až 6% striatálního objemu u krysy.[11] Ačkoli všechny dopaminové buňky SNc promítají do obou striosome (nebo patch) a matice neurochemické oddíly striata, většina z axonálního území neuronu dorzální vrstvy je v maticovém kompartmentu, zatímco většina axonálního pole neuronů ventrální vrstvy je ve striosomech.[6][10][11] Nigrostriatální dopaminové axony mohou také vést k axonovým kolaterálům, které se promítají do dalších oblastí mozku. Například některé SNc nigrostriatální dopaminové axony posílají axonové kolaterály do pedunkulopontinové jádro, ventrální palladium, subtalamické jádro, globus pallidus, amygdala a thalamus.[6][9][12]
Malý počet dorcálních neuronů dopaminového neuronu SNc také vyčnívá přímo do kůry, ačkoli většina dopaminergní inervace kůry pochází ze sousedních dopaminových neuronů VTA.[9]
Hřbetní striatum
The hřbetní striatum se nachází v subkortikální oblasti přední mozek. U primátů a jiných savců se dělí přední končetinou a bílá hmota trakt zvaný vnitřní kapsle,[13] na dvě části: kádové jádro a putamen.[14] U hlodavců je vnitřní tobolka špatně vyvinutá, takže kaudát a putamen nejsou odděleny, ale tvoří jednu velkou entitu zvanou caudate putamen (CPu).[15][16] Většina (asi 95%) buněk v dorzálním striatu jsou GABAergní středně ostnaté neurony (MSN), známé také jako neurony s ostnatou projekcí (SPN). Přibližně polovina těchto MSN obsahuje dopaminové receptory D1 a promítnout přímo do substantia nigra za vzniku přímá cesta bazálních ganglií, zatímco druhá polovina vyjadřuje receptory dopaminu D2 že projekt nepřímo do substantia nigra přes globus pallidus a subthalamic jádro tvořit nepřímá cesta bazálních ganglií.[17] Zbývajících 5% buněk je interneurony to jsou buď cholinergní neurony,[18] nebo jeden z několika typů GABAergních neuronů.[19] Axony a dendrity těchto interneuronů zůstávají ve striatu.
Kaudátové jádro a putamen dostávají excitační informace ze všech oblastí mozková kůra.[20] Tyto glutamátergní vstupy jsou obecně topograficky uspořádány tak, že putamen bere informace převážně ze senzorimotorické kůry, zatímco jádro caudate získává informace převážně z asociační kůry.[20] Kromě toho dorzální striatum přijímá excitační vstupy z jiných mozkových struktur, jako je thalamus,[21] a drobné excitační vstupy z hipokampus a amygdala.
Hřbetní striatum obsahuje neurochemicky definované kompartmenty zvané striosomy (také známé jako náplasti), které vykazují husté zabarvení μ-opioidního receptoru zabudované do matricového kompartmentu, který acetylcholinesteráza a kalbindin-D28K.[22]
Dopaminergní axonové terminály nigrostriatální dráhy se synchronizují na GABAergický MSN v dorzálním striatu. Tvoří synapse na těle buňky a dendritických oblastech hřídele, ale většinou na krku dendritické trny které také přijímají glutamátergní vstup do hlav stejných dendritických trnů.[1]
Funkce
Hlavní funkcí nigrostriatální dráhy je ovlivňování dobrovolné hnutí přes motorické smyčky bazálních ganglií. Spolu s mezolimbické a mezokortikální dopaminergní dráhy nigrostriatální dopaminová cesta může také ovlivňovat další mozkové funkce včetně poznání,[23] odměna a závislost.[24] Nigrostriatální dopaminergní neurony vykazují tonické a fázické vzorce aktivity vypalování neuronů. To může vést k různým vzorcům uvolňování dopaminu z terminálních axonů v dorzálním striatu a také z buněčného těla (soma) a dendritů v SNc a SNr.[25][26] Kromě uvolňování dopaminu mohou některé axony v nigrostriatální dráze také společně uvolňovat GABA.[27][28]

Nigrostriatální dráha ovlivňuje pohyb dvěma cestami, a přímá dráha pohybu a nepřímá dráha pohybu.[29][30]
Přímá dráha pohybu
The přímá cesta je zapojen do usnadnění hledaných pohybů. Projekce z dopaminový D1 receptor obsahující středně ostnaté neurony v jádru caudate a putamen synapse na tonicky aktivní buňky GABAergic v substantia nigra pars reticulata a vnitřním segmentu globus pallidus (GPi), které pak vyčnívají do thalamu. Protože striatonigral / striatoentopedunkulární a nigrothalamické dráhy jsou inhibiční, aktivace přímé dráhy vytváří celkovou síťovou excitaci na thalamu a na pohyb generovaný motorickou kůrou.
Nepřímá cesta pohybu
The nepřímá cesta je zapojen do potlačení nežádoucího pohybu. Projekce z dopaminový D2 receptor obsahující středně ostnaté neurony v jádru caudate a putamen synapse na tonicky aktivní buňky GABAergic ve vnějším segmentu globus pallidus (GPe), které pak přes excitační projektují na substantia nigra pars reticulata subthalmické jádro (STN). Protože striatopallidní a nigrothalamické dráhy jsou inhibiční, ale dráha subthalamická k nigře je excitační, aktivace nepřímé dráhy vytváří celkový čistý inhibiční účinek na thalamus a na pohyb motorickou kůrou.
Klinický význam
Parkinsonova choroba
Parkinsonova choroba je charakterizován hlavně závažnými problémy s motorem hypokineze tuhost, třes a posturální nerovnováha.[31] Ztráta dopaminových neuronů v nigrostriatální dráze je jedním z hlavních patologických rysů Parkinsonovy choroby.[32] Degenerace neuronů produkujících dopamin v substantia nigra pars compacta a komplexu putamen-kaudátu vede ke snížené koncentraci dopaminu v nigrostriatální dráze, což vede ke snížené funkci a charakteristickým příznakům.[33] Příznaky onemocnění se obvykle projeví až poté, co dojde ke ztrátě 80-90% dopaminové funkce.
Dyskineze vyvolaná levodopou
Dyskineze vyvolané levodopou (LID) je komplikace spojená s dlouhodobým užíváním Parkinsonovy léčby L-DOPA charakterizované mimovolním pohybem a kontrakcemi svalů. Tato porucha se vyskytuje až u 90% pacientů po 9 letech léčby. Použití L-DOPA u pacientů může vést k přerušení nigrostriatálních dopaminových projekcí a ke změnám v postsynaptických neuronech v bazálních gangliích.[34]
Schizofrenie
Je změněn presynaptický metabolismus dopaminu schizofrenie.[35][36]
Další cesty dopaminu
Mezi další hlavní cesty dopaminu patří:
Viz také
Reference
- ^ A b David Smith, A .; Paul Bolam, J. (01.07.1990). „Neuronová síť bazálních ganglií, jak odhalila studie synaptických spojení identifikovaných neuronů“. Trendy v neurovědách. 13 (7): 259–265. doi:10.1016 / 0166-2236 (90) 90106-K.. ISSN 0166-2236. PMID 1695400.
- ^ Tritsch, NX; Ding, JB; Sabatini, BL (říjen 2012). „Dopaminergní neurony inhibují striatální výdej nekanonickým uvolňováním GABA“. Příroda. 490 (7419): 262–6. Bibcode:2012Natur.490..262T. doi:10.1038 / příroda11466. PMC 3944587. PMID 23034651.
- ^ Diaz, Jaime. Jak drogy ovlivňují chování. Englewood Cliffs: Prentice Hall, 1996.
- ^ Zucca, Fabio A .; Basso, Emy; Cupaioli, Francesca A .; Ferrari, Emanuele; Sulzer, David; Casella, Luigi; Zecca, Luigi (leden 2014). „Neuromelanin z lidské substantia nigra: aktualizace“. Výzkum neurotoxicity. 25 (1): 13–23. doi:10.1007 / s12640-013-9435-r. ISSN 1476-3524. PMID 24155156.
- ^ Brichta, Lars; Greengard, Paul (2014). „Molekulární determinanty selektivní dopaminergní zranitelnosti u Parkinsonovy choroby: aktualizace“. Hranice v neuroanatomii. 8: 152. doi:10.3389 / fnana.2014.00152. ISSN 1662-5129. PMC 4266033. PMID 25565977.
- ^ A b C d E Prensa, L .; Giménez-Amaya, J. M .; Parent, A .; Bernácer, J .; Cebrián, C. (2009). "Nigrostriatální dráha: axonální kolateralizace a kompartmentová specificita". Journal of Neural Transmission. Supplementum (73): 49–58. doi:10.1007/978-3-211-92660-4_4. ISBN 978-3-211-92659-8. ISSN 0303-6995. PMID 20411767.
- ^ Nemoto, C .; Hida, T .; Arai, R. (1999-10-30). „Kalretinin a kalbindin-D28k v dopaminergních neuronech středního mozku potkana: trojnásobně značená imunohistochemická studie“. Výzkum mozku. 846 (1): 129–136. doi:10.1016 / s0006-8993 (99) 01950-2. ISSN 0006-8993. PMID 10536220.
- ^ Dopeso-Reyes, Iria G .; Rico, Alberto J .; Roda, Elvira; Sierra, Salvador; Pignataro, Diego; Lanz, Maria; Sucunza, Diego; Chang-Azancot, Luis; Lanciego, Jose L. (2014). „Obsah kalbindinu a různá zranitelnost dopaminergních neuronů vylučujících střední mozek u makaků“. Hranice v neuroanatomii. 8: 146. doi:10.3389 / fnana.2014.00146. ISSN 1662-5129. PMC 4253956. PMID 25520629.
- ^ A b C d E F Haber, S. N. (12. 12. 2014). „Místo dopaminu v okruhu kortiko-bazálních ganglií“. Neurovědy. 282: 248–257. doi:10.1016 / j.neuroscience.2014.10.008. ISSN 1873-7544. PMC 5484174. PMID 25445194.
- ^ A b Gerfen, C. R .; Herkenham, M .; Thibault, J. (prosinec 1987). „Neostriatální mozaika: II. Mezostriatální dopaminergní a nedopaminergní systémy zaměřené na patch a matici“. The Journal of Neuroscience. 7 (12): 3915–3934. doi:10.1523 / JNEUROSCI.07-12-03915.1987. ISSN 0270-6474. PMC 6569093. PMID 2891799.
- ^ A b Matsuda, Wakoto; Furuta, Takahiro; Nakamura, Kouichi C .; Hioki, Hiroyuki; Fujiyama, Fumino; Arai, Ryohachi; Kaneko, Takeshi (14.01.2009). „Jednotlivé nigrostriatální dopaminergní neurony tvoří široce rozšířené a vysoce husté axonální arborizace v neostriatu“. The Journal of Neuroscience. 29 (2): 444–453. doi:10.1523 / JNEUROSCI.4029-08.2009. ISSN 1529-2401. PMC 6664950. PMID 19144844.
- ^ Prensa, L .; Parent, A. (2001-09-15). „Nigrostriatální dráha u potkana: Jednoosá studie vztahu mezi dorzálními a ventrálními nigrálními neurony a striosomovými / matricovými striatálními oddíly“. The Journal of Neuroscience. 21 (18): 7247–7260. doi:10.1523 / JNEUROSCI.21-18-07247.2001. ISSN 1529-2401. PMC 6762986. PMID 11549735.
- ^ Emos, Marc Christopher; Agarwal, Sanjeev (2019), „Neuroanatomy, Internal Capsule“, StatPearls, StatPearls Publishing, PMID 31194338, vyvoláno 2019-10-06
- ^ Mai, Jürgen K. Atlas lidského mozku. Majtanik, Milan, Paxinos, George, 1944- (4. vydání). Amsterdam. ISBN 9780128028001. OCLC 934406284.
- ^ Coizet, Veronique; Heilbronner, Sarah R .; Carcenac, Carole; Mailly, Philippe; Lehman, Julia F .; Savasta, Marc; David, Oivier; Deniau, Jean-Michel; Groenewegen, Henk J .; Haber, Suzanne N. (8. března 2017). „Organizace přední končetiny vnitřní tobolky u krysy“. The Journal of Neuroscience. 37 (10): 2539–2554. doi:10.1523 / JNEUROSCI.3304-16.2017. ISSN 1529-2401. PMC 5354315. PMID 28159909.
- ^ Paxinos, George, 1944- (07.11.2013). Mozek krysy ve stereotaxických souřadnicích. Watson, Charles, 1943- (sedmé vydání). Amsterdam. ISBN 9780123919496. OCLC 859555862.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Gerfen, Charles R .; Surmeier, D. James (2011). "Modulace striatálních projekčních systémů dopaminem". Roční přehled neurovědy. 34: 441–466. doi:10.1146 / annurev-neuro-061010-113641. ISSN 1545-4126. PMC 3487690. PMID 21469956.
- ^ Gonzales, Kalynda K .; Smith, Yoland (září 2015). „Cholinergní interneurony v dorzálním a ventrálním striatu: anatomické a funkční aspekty za normálních a nemocných podmínek“. Annals of the New York Academy of Sciences. 1349 (1): 1–45. Bibcode:2015NYASA1349 .... 1G. doi:10.1111 / nyas.12762. ISSN 1749-6632. PMC 4564338. PMID 25876458.
- ^ Tepper, James M .; Koós, Tibor; Ibanez-Sandoval, Osvaldo; Tecuapetla, Fatuel; Faust, Thomas W .; Assous, Maxime (2018). „Heterogenita a rozmanitost striatálních GABAergních interneuronů: aktualizace 2018“. Hranice v neuroanatomii. 12: 91. doi:10.3389 / fnana.2018.00091. ISSN 1662-5129. PMC 6235948. PMID 30467465.
- ^ A b Haber, Suzanne N. (březen 2016). „Kortikostriatální obvody“. Dialogy v klinické neurovědě. 18 (1): 7–21. ISSN 1958-5969. PMC 4826773. PMID 27069376.
- ^ Smith, Yoland; Galvan, Adriana; Ellender, Tommas J .; Doig, Natalie; Villalba, Rosa M .; Huerta-Ocampo, Icnelia; Wichmann, Thomas; Bolam, J. Paul (2014). „Thalamostriatální systém v normálním a nemocném stavu“. Frontiers in Systems Neuroscience. 8: 5. doi:10.3389 / fnsys.2014.00005. ISSN 1662-5137. PMC 3906602. PMID 24523677.
- ^ Brimblecombe, Katherine R .; Cragg, Stephanie J. (15. února 2017). „Striosomové a maticové kompartmenty striata: Cesta labyrintem od neurochemie k funkci“. ACS Chemical Neuroscience. 8 (2): 235–242. doi:10.1021 / acschemneuro.6b00333. ISSN 1948-7193. PMID 27977131.
- ^ Boot, Nathalie; Baas, Matthijs; van Gaal, Simon; Cools, Roshan; De Dreu, Carsten K. W. (červenec 2017). „Kreativní poznání a dopaminergní modulace fronto-striatálních sítí: Integrativní revize a výzkumná agenda“. Neurovědy a biobehaviorální recenze. 78: 13–23. doi:10.1016 / j.neubiorev.2017.04.007. ISSN 1873-7528. PMID 28419830.
- ^ Wise, RA (říjen 2009). „Role pro nigrostriatální - nejen mezokortikolimbické - dopamin jako odměna a závislost“. Trendy v neurovědách. 32 (10): 517–524. doi:10.1016 / j.tins.2009.06.004. PMC 2755633. PMID 19758714.
- ^ Rice, M. E.; Patel, J. C .; Cragg, S. J. (2011-12-15). „Uvolňování dopaminu v bazálních gangliích“. Neurovědy. 198: 112–137. doi:10.1016 / j.neuroscience.2011.08.066. ISSN 1873-7544. PMC 3357127. PMID 21939738.
- ^ Rice, Margaret E .; Patel, Jyoti C. (2015-07-05). „Uvolňování somatodendritického dopaminu: nedávné mechanistické poznatky“. Filozofické transakce Královské společnosti v Londýně. Série B, Biologické vědy. 370 (1672): 20140185. doi:10.1098 / rstb.2014.0185. ISSN 1471-2970. PMC 4455754. PMID 26009764.
- ^ Tritsch, Nicolas X .; Granger, Adam J .; Sabatini, Bernardo L. (březen 2016). "Mechanismy a funkce společného vydání GABA". Recenze přírody. Neurovědy. 17 (3): 139–145. doi:10.1038 / nrn.2015.21. ISSN 1471-0048. PMC 6980171. PMID 26865019.
- ^ Trudeau, Louis-Eric; Hnasko, Thomas S .; Wallén-Mackenzie, Asa; Morales, Marisela; Rayport, Steven; Sulzer, David (2014). „Mnohojazyčná povaha dopaminových neuronů“. Pokrok ve výzkumu mozku. 211: 141–164. doi:10.1016 / B978-0-444-63425-2.00006-4. ISBN 9780444634252. ISSN 1875-7855. PMC 4565795. PMID 24968779.
- ^ Kravitz, Alexxai V .; Kreitzer, Anatol C. (červen 2012). „Striatální mechanismy, které jsou základem pohybu, posilování a trestání“. Fyziologie. 27 (3): 167–177. doi:10.1152 / physiol.00004.2012. ISSN 1548-9221. PMC 3880226. PMID 22689792.
- ^ Kravitz, Alexxai V .; Freeze, Benjamin S .; Parker, Philip R. L .; Kay, Kenneth; Thwin, Myo T .; Deisseroth, Karl; Kreitzer, Anatol C. (2010-07-29). „Regulace parkinsonského motorického chování pomocí optogenetické kontroly obvodů bazálních ganglií“. Příroda. 466 (7306): 622–626. Bibcode:2010Natur.466..622K. doi:10.1038 / nature09159. ISSN 1476-4687. PMC 3552484. PMID 20613723.
- ^ Cenci, Angela M (2006). „Post-versus presynaptická plastika u dyskineze vyvolané L-DOPA“ (PDF). Journal of Neurochemistry. 99 (2): 381–92. doi:10.1111 / j.1471-4159.2006.04124.x. PMID 16942598.
- ^ Deumens, Ronald (21. června 2002). „Modelování Parkinsonovy nemoci u potkanů: vyhodnocení lézí 6-OHDA na Nigrostriatal Pathway“. Experimentální neurologie. 175 (2): 303–17. doi:10.1006 / ex.2002,7891. PMID 12061862.
- ^ Groger, Adraine (8. ledna 2014). „Snížení dopaminu u pacientů s Parkinsonovou chorobou u společnosti Conststantia Nigra potvrzeno spektroskopickým zobrazením magnetickou rezonancí in vivo“. PLOS ONE. 9 (1): e84081. Bibcode:2014PLoSO ... 984081G. doi:10.1371 / journal.pone.0084081. PMC 3885536. PMID 24416192.
- ^ Niethammer, Martin (květen 2012). „Funkční neuroimaging u Parkinsonovy choroby“. Perspektivy Cold Spring Harbor v medicíně. 2 (5): a009274. doi:10.1101 / cshperspect.a009274. PMC 3331691. PMID 22553499.
- ^ Fusar-Poli, Paolo; Meyer-Lindenberg, Andreas (1. ledna 2013). „Striatální presynaptický dopamin u schizofrenie, část II: metaanalýza studií [(18) F / (11) C] -DOPA PET“. Bulletin schizofrenie. 39 (1): 33–42. doi:10.1093 / schbul / sbr180. ISSN 1745-1701. PMC 3523905. PMID 22282454.
- ^ Weinstein, Jodi J .; Chohan, Muhammad O .; Slifstein, Mark; Kegeles, Lawrence S .; Moore, Holly; Abi-Dargham, Anissa (1. ledna 2017). „Abnormality dopaminu specifické pro cestu u schizofrenie“. Biologická psychiatrie. 81 (1): 31–42. doi:10.1016 / j.biopsych.2016.03.2104. ISSN 1873-2402. PMC 5177794. PMID 27206569.