Folikulogeneze - Folliculogenesis
Tento článek je věcná přesnost je sporný. (Březen 2012) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
- Ačkoli je proces u mnoha zvířat podobný, bude se tento článek zabývat výhradně člověk folikulogeneze.

1 - Menstruace
2 - Vývoj folikulu
3 - Zralý folikul
4 - Ovulace
5 - Žluté tělo
6 - Zhoršení žlutého tělíska
v biologie, folikulogeneze je zrání ovariální folikul, hustě zabalená skořápka somatické buňky který obsahuje nezralé oocyt. Folliculogenesis popisuje postup řady malých prvotní folikuly do velkého preovulační folikuly která se vyskytuje částečně během menstruační cyklus.
Na rozdíl od muže spermatogeneze, která může trvat neurčitě, folikulogeneze končí, když zbývající folikuly v vaječníky nejsou schopni reagovat na hormonální podněty, které dříve nabíraly některé folikuly k dozrávání. Toto vyčerpání v zásobování folikulem signalizuje začátek menopauza.
Přehled
Primární role folikulu je oocyt Podpěra, podpora. Od narození vaječníky lidské ženy obsahují řadu nezralých, prvotní folikuly. Každý z těchto folikulů obsahuje podobně nezralé primární oocyt. Na puberta spojky folikulů začínají folikulogenezí, vstupují do růstového vzorce, který končí smrtí (apoptóza) nebo ovulací (proces, kdy oocyt opouští folikul).
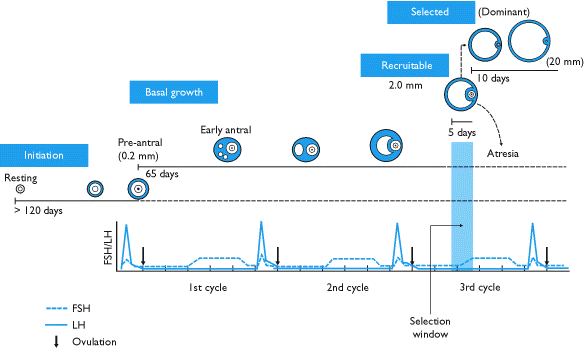
Během vývoje folikulů prochází primordiální folikuly řadou kritických změn v charakteru, histologicky i hormonálně. Nejprve se změní na primární folikuly a později na sekundární folikuly. Folikuly pak přecházejí do terciární nebo antrální folikuly. V této fázi vývoje se stávají závislými na hormonech, zejména FSH, což způsobuje podstatné zvýšení jejich rychlosti růstu. Pozdní terciární nebo preovulační folikul praskne a vybije oocyt (který se stal a sekundární oocyt ), končící folikulogenezí.
Fáze vývoje
Folikulogeneze je kontinuální, což znamená, že vaječník kdykoli obsahuje folikuly v mnoha stádiích vývoje. Většina folikulů umírá a nikdy nedokončí vývoj. Několik se plně vyvine a vytvoří sekundární oocyt, který se uvolní prasknutím folikulu v procesu zvaném ovulace.
Rostoucí folikul prochází následujícími odlišnými stádii, která jsou definována určitými strukturálními charakteristikami:
Z větší perspektivy celá folikulogeneze, od prvotního po preovulační folikul, patří do stadia ootidogeneze z oogeneze.
| Etapa | Popis | Velikost |
| Prvotní | Spící, malé, pouze jedna vrstva bytu granulosa buňky | Primordiální folikuly mají průměr asi 0,03-0,05 mm. |
| Hlavní | Mitotické buňky, kvádrové granulózní buňky | Průměr téměř 0,1 mm |
| Sekundární | Přítomnost buňky theca, více vrstev granulózových buněk | Folikul má nyní průměr 0,2 mm |
| Brzy terciární | Časný terciární folikul je libovolně rozdělen do pěti tříd. Folikuly třídy 1 mají průměr 0,2 mm, třída 2 asi 0,4 mm, třída 3 asi 0,9 mm, třída 4 asi 2 mm a třída 5 asi 5 mm. | |
| Pozdní terciární | Plně formovaný antrum, žádná další cytodiferenciace, žádný nový pokrok | Folikuly třídy 6 mají průměr asi 10 mm, třída 7 asi 16 mm a třída 8 asi 20 mm. Je běžné, že nedominantní folikuly rostou za třídu 5, ale zřídka je více než jeden folikul třídy 8. |
| Předovulační | Budování růstu koncentrace estrogenu, všech ostatních folikulů atretický nebo mrtvý |
Kromě toho folikuly, které vytvořily antrum jsou nazývány antrální folikuly nebo Graafiánské folikuly. Definice se liší v tom, kde k tomuto posunu dochází ve stagingu uvedeném výše, přičemž některé uvádějí, že k němu dochází při vstupu do sekundární etapa,[1] a další uvádějící, že k němu dochází při vstupu do terciární etapa.[2]
Až do preovulačního stadia obsahuje folikul primární oocyt, který je zastaven v profázi meióza I. Během pozdního preovulačního stadia oocyt pokračuje v meióze a stává se sekundárním oocytem, který je zadržen metafáze II.

Prvotní
Ve 18–22 týdnech po početí byla mozková kůra ženská vaječník (plodový ženský vaječník) obsahuje svůj maximální počet folikulů (v průměru asi 4 až 5 milionů, ale jednotlivé maximální populace se pohybují od 6 do 7 milionů).[3] Tyto prvotní folikuly obsahují nezralé oocyty obklopené plochými, plochými plochami granulózní buňky (podpůrné buňky), které jsou odděleny od prostředí oocytů bazální laminou. Jsou v klidu, ukazují málo k ne biologická aktivita. Protože primordiální folikuly mohou být u člověka spící až 50 let, délka ovariálního cyklu tuto dobu nezahrnuje.
Zásoba folikulů před narozením mírně klesá a v průměrném případě na pubertu na 180 000 (populace v pubertě se pohybuje od 25 000 do 1,5 milionu).[3] Na základě „neefektivní“ povahy folikulogeneze (o které se pojednává dále), pouze 400 z těchto folikulů dosáhne preovulační fáze. Na menopauza, zůstává pouze 1 000 folikulů. Zdá se pravděpodobné, že časná menopauza nastává u žen s nízkou populací při narození a pozdní menopauza nastává u žen s vysokou populací při narození, ale zatím neexistují žádné klinické důkazy.[3]
Proces, kterým se prvotní buňky „probouzí“, je znám jako počáteční nábor. Výzkum ukázal, že počáteční nábor je zprostředkován protiváhou různých stimulačních a inhibičních hormonů a lokálně produkovaných růstových faktorů.[4]
Hlavní
V době aktivace vaječníkových folikulů se granulózní buňky primordiálních folikulů mění z ploché na kvádrovitou strukturu, což znamená začátek primární folikul. Oocytový genom je aktivován a geny se stávají přepsal. Základní parakrinní signalizace jsou vytvořeny dráhy, které jsou životně důležité pro komunikaci mezi folikulem a oocytem. Oocyt i folikul dramaticky rostou a jejich průměr se zvětšuje na téměř 0,1 mm.
Primární folikuly vyvíjejí receptory hormon stimulující folikuly (FSH) v tomto okamžiku, ale jsou nezávislé na gonadotropinu až do antrální fáze. Výzkum však ukázal, že přítomnost FSH urychluje růst folikulů in vitro.
Glykoproteinová polymerní kapsle s názvem zona pellucida se tvoří kolem oocytu a odděluje jej od okolních buněk granulózy. Zona pellucida, která po ovulaci zůstává s oocytem, obsahuje enzymy, které katalyzují spermie umožnit průnik.
Sekundární
Buňky podobné stromům podobné stromům jsou přijímány signály vylučovanými oocyty. Obklopují nejvzdálenější vrstvu folikulu, bazální lamina, a podstoupit cytodiferenciaci, aby se stal theca externa a theca interna. Mezi těmito dvěma vrstvami teku se tvoří složitá síť kapilárních cév, která začíná cirkulovat krev do a z folikulu.
Pozdější sekundární folikul je histologicky a strukturně poznamenán plně dospělým oocytem obklopeným zona pellucida, přibližně devíti vrstvami buněk granulózy, bazální lamina, theca interna, kapilární síť a theca externa. Vývoj antra také začíná probíhat ve stadiu sekundárního folikulu
Tvorba Antrum
Tvoření tekutiny naplněné dutiny sousedící s oocytem zvané antrum označuje folikul jako antrální folikul, na rozdíl od tzv preantrální folikul stále mu chybí antrum. Antrální folikul se také nazývá a Graafiánský folikul.
Definice se liší v tom, ve které fázi tento posun nastává, přičemž některé označují folikuly v sekundární fáze jako antral,[1] a další je označují jako předpremiéra.[2]
Brzy terciární
V terciární folikulse vytvořila základní struktura zralého folikulu a nejsou detekovatelné žádné nové buňky. Buňky granulózy a theky nadále procházejí mitotis souběžně se zvýšením objemu antra. Terciární folikuly mohou dosáhnout obrovské velikosti, která je omezena pouze dostupností FSH, na které je nyní závislá.
Působením morfogenního gradientu vylučovaného oocyty podléhají granulózní buňky terciárního folikulu diferenciaci do čtyř odlišných podtypů: corona radiata, obklopující zona pellucida; membrana, vnitřek k bazální vrstvě; periantral, přiléhající k antrum a kupa oophorous, který spojuje buňky membrány a corona radiata granulosa dohromady. Každý typ buňky se chová odlišně v reakci na FSH.
Buňky theca interna exprimují receptory pro luteinizační hormon (LH ). LH indukuje produkci androgeny zejména buňkami theky androstendion, které jsou aromatizovány buňkami granulózy estrogeny, především estradiol. V důsledku toho hladiny estrogenu začínají stoupat.
Pozdní terciární a preovulační (folikulární fáze menstruačního cyklu)
V tomto okamžiku zemřela většina skupiny folikulů, které zahájily růst. Tento proces smrti folikulů je známý jako atrézie a vyznačuje se radikálem apoptóza všech základních buněk a oocytů. I když není známo, co způsobuje atrézii, bylo prokázáno, že přítomnost vysokých koncentrací FSH tomu brání.
Nárůst FSH hypofýzy způsobený rozpadem žlutého tělíska na konci menstruačního cyklu urychluje nábor pěti až sedmi folikulů třídy 5 k účasti v dalším cyklu. Tyto folikuly vstupují na konec předchozího menstruačního cyklu a přecházejí do folikulární fáze toho dalšího. Vybrané folikuly, nazývané antrální folikuly, mezi sebou soutěží o FSH indukující růst.
Je diskutován vzor tohoto vývoje kohorty pěti až sedmi antrálních folikulů. Existují teorie nepřetržitého náboru antrálních folikulů, teorie jediné náborové epizody na konci luteální fáze a v poslední době existují důkazy o náborovém modelu vyznačeném 2 - 3 vlnami náboru a vývoje folikulů během menstruačního cyklu (pouze jedna z nich je ve skutečnosti ovulační vlna).[5]
V reakci na vzestup FSH začínají antrální folikuly vylučovat estrogen a inhibin, které mají negativní vliv zpětné vazby na FSH.[6] Folikuly, které mají méně FSH receptory nebude se moci dále rozvíjet; projeví se zpomalením jejich rychlosti růstu a stanou se atretickými. Nakonec bude životaschopný pouze jeden folikul. Tento zbývající folikul, nazývaný dominantní folikul, poroste rychle a dramaticky - až do průměru 20 mm - a stane se preovulační folikul.
Poznámka: Mnoho zdrojů zkresluje tempo růstu folikulů, některé dokonce naznačují, že trvá jen čtrnáct dní, než se primordiální folikul stane preovulačním. Folikulární fáze menstruačního cyklu ve skutečnosti znamená dobu mezi výběrem terciárního folikulu a jeho následným růstem do preovulačního folikulu. Skutečná doba pro vývoj folikulu se liší.
Růst dominantního folikulu během folikulární fáze je asi 1,5 mm za den (± 0,1 mm), a to jak v přirozených cyklech, tak pro jakýkoli dominantní folikul vyvíjející se při užívání kombinovaná perorální antikoncepční pilulka.[7] Předvádění kontrolovaná ovariální hyperstimulace vede k většímu náboru folikulů a roste přibližně o 1,6 mm denně.[7]
Ovulace a žluté tělísko
Na konci folikulární (nebo proliferativní) fáze třináctého dne menstruačního cyklu se cumulus oophorus vrstva preovulačního folikulu vyvine otvor, nebo stigma, a vylučovat oocyt doplňkem kumulativních buněk v procesu zvaném ovulace. V přirozených cyklech může dojít k ovulaci ve folikulích, které jsou alespoň 14 mm.[8]
Oocyt je technicky stále sekundární oocyt, suspendovaný v metafáze II meiózy. Bude se vyvíjet v ootid a rychle poté na vajíčko (po dokončení meiózy II) až po oplodnění. Oocyt nyní bude cestovat dolů jedním z vejcovody být nakonec vypuštěn menstruací v případě, že je neoplodněný nebo pokud není úspěšně implantován do děloha (pokud dříve oplodněno ).
Prasklý folikul projde dramatickou transformací na corpus luteum, steroidiogenní shluk buněk, který udržuje endometrium dělohy vylučováním velkého množství progesteron a menší množství estrogen.
Tyto dva kroky, i když nejsou součástí folikulogeneze, jsou pro úplnost zahrnuty. Jsou o nich diskutovány v celém rozsahu jejich příslušnými články a jsou uvedeny do perspektivy prostřednictvím menstruační cyklus článek. Doporučuje se, aby byla tato tři témata přezkoumána.
Funkce hormonů
Stejně jako u většiny věcí souvisejících s reprodukčním systémem je folikulogeneze řízena endokrinní systém. Pět hormonů se účastní složitého procesu pozitivní a negativní zpětné vazby k regulaci folikulogeneze. Oni jsou:
- hormon uvolňující gonadotropin (GnRH) vylučovaný hypotalamus
- dva gonadotropiny:
- folikuly stimulující hormon (FSH)
- luteinizační hormon (Levý)
- estrogen
- progesteron
GnRH stimuluje uvolňování FSH a LH z přední hypofýzy, což bude mít později stimulační účinek na růst folikulů (ne však okamžitě, protože na FSH a LH jsou závislé pouze antrální folikuly). Když se buňky theca vytvoří v terciárním folikulu, množství estrogenu prudce vzroste (androgen odvozený od theca je aromatizován do estrogenu buňkami granulózy).
Při nízké koncentraci estrogen inhibuje gonadotropiny, ale vysoká koncentrace estrogenu je stimuluje. Kromě toho, jak se vylučuje více estrogenu, produkují buňky theca více receptorů LH, které podněcují buňky theca a vytvářejí více androgenu, který se po proudu stane estrogenem. Tato smyčka pozitivní zpětné vazby způsobí, že LH prudce stoupá, a právě tento hrot způsobuje ovulaci.
Po ovulaci stimuluje LH tvorbu žlutého tělíska. Estrogen od té doby klesl na negativní stimulační hladiny po ovulaci, a proto slouží k udržení koncentrace FSH a LH. Inhibin, který je také vylučován žlutým tělem, přispívá k inhibici FSH. Progesteron, vylučovaný žlutým tělem, inhibuje růst folikulů a udržuje těhotenství.
Endokrinní systém se shoduje s menstruačním cyklem a v průběhu normální folikulogeneze prochází třinácti cykly (a tedy třinácti špičkami LH). Koordinovaná enzymová signalizace a časově specifická exprese hormonálních receptorů však zajišťuje, že růst folikulů nebude během těchto předčasných špiček neregulován.
Počet folikulů

Nedávno dvě publikace zpochybnily myšlenku, že kolem doby narození je stanoven konečný počet folikulů.[10][11] V postnatálním myším vaječníku bylo hlášeno obnovení ovariálních folikulů ze zárodečných kmenových buněk (pocházejících z kostní dřeně a periferní krve). Probíhají studie, které se pokoušejí replikovat tyto výsledky, ale studie populací ve 325 lidských vaječnících nenalezla žádné podpůrné důkazy o doplňování folikulů.[3]
V roce 2010 vědci z University of Edinburgh zjistili, že v době, kdy je ženám 30 let, zůstává pouze 10% jejich nerostoucích folikulů (NGF).[9] Při narození mají ženy všechny folikuly pro folikulogenezi a neustále klesají až do menopauza.
Vyčerpání ovariální rezervy
Jak ženy (a myši) stárnou, dvojvláknové zlomy se hromadí v jejich prvotní rezervě folikulů. Tyto folikuly obsahují primární oocyty, které jsou zastaveny v profázi prvního buněčného dělení meiózy. Přerušení dvouřetězcových vláken jsou během meiózy přesně opravena hledáním a budováním shodného řetězce (nazývaného „homologní rekombinační oprava“). Titus a kol.[12] (2013) zjistili, že s věkem lidí (a myší) exprese čtyř klíčových genů pro opravu DNA nezbytných pro homologní rekombinační opravu klesá v oocytech. Předpokládali, že oprava dvouřetězcového zlomu DNA je nezbytná pro udržení rezervy oocytů a že pokles účinnosti opravy s věkem hraje klíčovou roli při vyčerpání rezervy vaječníků (stárnutí vaječníků).
Viz také
Další obrázky

Sekce vaječníku. (# 5 až # 9 představují stadia folikulogeneze)

přechodný primární folikul.

Reference
- ^ A b Stránka 769, část „tvorba antra“ v: Sherwood, Lauralee. (2010). Fyziologie člověka: od buněk k systému. Austrálie; Spojené státy: Brooks / Cole. ISBN 978-0-495-39184-5.
- ^ A b Stránka 76 v: Vandenhurk, R .; Bevers, M .; Beckers, J. (1997). "In-vivo a in-vitro vývoj preantrálních folikulů". Theriogenologie. 47: 73–82. doi:10.1016 / S0093-691X (96) 00341-X.
- ^ A b C d Wallace, WHB; Kelsey, TW (2010). "Lidská ovariální rezerva od početí do menopauzy". PLOS ONE. 5 (1): e8772. doi:10,1371 / journal.pone.0008772. PMC 2811725. PMID 20111701.
- ^ Fortune J, Cushman R, Wahl C, Kito S (2000). "Přechod mezi prvotním a primárním folikulem". Mol Cell Endocrinol. 163 (1–2): 53–60. doi:10.1016 / S0303-7207 (99) 00240-3. PMID 10963874. S2CID 8746207.
- ^ „Ovariální antrální folikulogeneze během menstruačního cyklu člověka: přehled“. ResearchGate. Citováno 2019-01-23.
- ^ de Ziegler D (2007), „Role FSH a LH během folikulární fáze: vhled do přirozeného cyklu IVF“, RBM Online svazek 15 č. 5, strana 508
- ^ A b Baerwald, Angela R .; Walker, Randy A .; Pierson, Roger A. (2009). „Tempo růstu ovariálních folikulů během přirozených menstruačních cyklů, cyklů orální antikoncepce a cyklů stimulace vaječníků“. Plodnost a sterilita. 91 (2): 440–449. doi:10.1016 / j.fertnstert.2007.11.054. ISSN 0015-0282. PMID 18249401.
- ^ Stránka 34 v: Michael K. Skinner (2018). Encyklopedie reprodukce (2. vyd.). Akademický tisk. ISBN 9780128151457.
- ^ A b Wallace, W. Hamish B .; Thomas W. Kelsey (2010-01-27). "Lidská ovariální rezerva od početí do menopauzy". PLOS ONE. 5 (1): e8772. doi:10,1371 / journal.pone.0008772. PMC 2811725. PMID 20111701.
- ^ Johnson J, Bagley J, Skaznik-Wikiel M, Lee H, Adams G, Niikura Y, Tschudy K, Tilly J, Cortes M, Forkert R, Spitzer T, Iacomini J, Scadden D, Tilly J (2005). „Generování oocytů ve vaječnících dospělých savců předpokládanými zárodečnými buňkami v kostní dřeni a periferní krvi“. Buňka. 122 (2): 303–15. doi:10.1016 / j.cell.2005.06.031. PMID 16051153. S2CID 19006732.
- ^ Johnson J, Canning J, Kaneko T, Pru J, Tilly J (2004). "Germline kmenové buňky a folikulární obnova v postnatálním savčím vaječníku". Příroda. 428 (6979): 145–50. doi:10.1038 / nature02316. PMID 15014492. S2CID 1124530.
- ^ Titus, S; Li, F; Stobezki, R; Akula, K; Unsal, E; Jeong, K; Dickler, M; Robson, M; Moy, F; Goswami, S; Oktay, K (2013). „Porucha opravy dvouvláknového zlomu DNA související s BRCA1 vede ke stárnutí vaječníků u myší a lidí“. Sci Transl Med. 5 (172): 172. doi:10.1126 / scitranslmed.3004925. PMC 5130338. PMID 23408054.
- Caglar G, Asimakopoulos B, Nikolettos N, Diedrich K, Al-Hasani S (2005). "Rekombinantní LH při stimulaci vaječníků". Reprod Biomed Online. 10 (6): 774–85. doi:10.1016 / S1472-6483 (10) 61123-6. PMID 15970010.
- Fortune, JE; Yang, MY; Muruvi, W (2010). Lucy, MC; Pate, JL; Smith, MF; Spencer, TE (eds.). „Nejranější stadia vývoje folikulů: tvorba a aktivace folikulů“. Soc Reprod hnojivo Suppl. 67: 203–16. doi:10,7313 / upo9781907284991,018. ISBN 9781907284991. PMC 5316466. PMID 21755674.
- Gougeon A (1996). „Regulace ovariálního folikulárního vývoje u primátů: fakta a hypotézy“. Endocr Rev. 17 (2): 121–55. doi:10.1210 / er.17.2.121. PMID 8706629.
- Gougeon A (1986). "Dynamika folikulárního růstu u člověka: model z předběžných výsledků". Hum Reprod. 1 (2): 81–7. doi:10.1093 / oxfordjournals.humrep.a 136365. PMID 3558758.
- van den Hurk R, Zhao J (2005). "Tvorba savčích oocytů a jejich růst, diferenciace a zrání ve vaječníkových folikulech". Theriogenologie. 63 (6): 1717–51. doi:10.1016 / j.theriogenology.2004.08.005. PMID 15763114.
- Uzumcu, Mehmet; Zachow, Rob (2007). „Vývojová expozice endokrinním disruptorům v prostředí: Důsledky ve vaječníku a na reprodukční funkci žen“. Reprodukční toxikologie. 23 (3): 337–352. doi:10.1016 / j.reprotox.2006.10.006. PMC 1950429. PMID 17140764.