Chorismát mutáza - Chorismate mutase
| Chorismát mutáza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Krystalová struktura chorismát mutázy s analogovým vázáním přechodového stavu | |||||||||
| Identifikátory | |||||||||
| EC číslo | 5.4.99.5 | ||||||||
| Číslo CAS | 9068-30-8 | ||||||||
| Databáze | |||||||||
| IntEnz | IntEnz pohled | ||||||||
| BRENDA | Vstup BRENDA | ||||||||
| EXPASY | Pohled NiceZyme | ||||||||
| KEGG | Vstup KEGG | ||||||||
| MetaCyc | metabolická cesta | ||||||||
| PRIAM | profil | ||||||||
| PDB struktur | RCSB PDB PDBe PDBsum | ||||||||
| Genová ontologie | AmiGO / QuickGO | ||||||||
| |||||||||
v enzymologie, chorismát mutáza (ES 5.4.99.5 ) je enzym že katalyzuje the chemická reakce pro převod chorismát na prefenovat v cesta k výrobě fenylalanin a tyrosin, také známý jako shikimate Tento enzym tedy má jeden Podklad, chorismát a jeden produkt, prefenovat. Chorismát mutáza se nachází v bodě větve v cestě. Enzym směruje substrát, chorismát na biosyntézu tyrosinu a fenylalaninu a dále od tryptofan.[1] Jeho role v udržování rovnováhy těchto aromatických aminokyselin v buňce je zásadní.[2] Toto je jediný známý příklad přirozeně se vyskytujícího enzymu katalyzujícího a pericyklická reakce.[2][poznámka 1] Chorismát mutáza se vyskytuje pouze u hub, bakterií a vyšších rostlin. Některé odrůdy tohoto proteinu mohou používat morpheein model alosterická regulace.[4]
Rodina proteinů
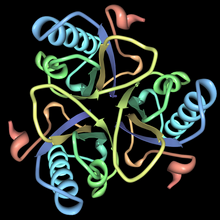
Tento enzym patří do rodiny izomerázy, konkrétně intramolekulární transferázy které přenášejí funkční skupiny. The systematické jméno této třídy enzymů je chorismát pyruvatemutáza. Chorismát mutáza, také známý jako hydroxyfenylpyruvát syntáza, podílí se na biosyntéze fenylalaninu, tyrosinu a tryptofanu.[1] Struktury chorismát mutáz se liší v různých organismech, ale většina patří do rodiny AroQ a je charakterizována propleteným homodimerem 3-šroubovicových podjednotek. Většina chorismátových mutáz v této rodině vypadá podobně jako u mutací Escherichia coli. Například sekundární struktura chorismát mutázy z droždí je velmi podobný tomu z E-coli. Chorimate mutase v rodině AroQ jsou v přírodě běžnější a jsou široce distribuovány mezi prokaryoty.[1] Pro optimální funkci je obvykle nutné je doprovázet jiným enzymem, jako je prefanátdehydrogenáza. Tyto chorismátové mutázy jsou obvykle bifunkční enzymy, což znamená, že obsahují dvě katalytické kapacity ve stejném polypeptidovém řetězci.[1] Avšak chorismát mutáza eukaryotických organismů je častěji monofunkční. Existují organismy jako např Bacillus subtilis jejichž chorismát mutáza má úplně jinou strukturu a je monofunkční. Tyto enzymy patří do rodiny AroH a jsou charakterizovány trimerní topologií α / β barelu.[5]
Mechanismus katalýzy
Konverze chorismátu na prefenát je první odhodlaný krok na cestě k výrobě aromatické aminokyseliny: tyrosin a fenylalanin. Přítomnost chorismát mutázy zvyšuje rychlost reakce milionkrát.[6] Při absenci enzymové katalýzy tento mechanismus probíhá jako koordinovaný, ale asynchronní krok a je exergonický proces. Mechanismus této transformace je formálně a Claisenův přesmyk, podporované kinetickými a izotopovými údaji uváděnými Knowlesem a kol[7]

E-coli a kvasinková chorismát mutáza mají omezenou sekvenční homologii, ale jejich aktivní místa obsahují podobné zbytky. Aktivní místo kvasinkové chorismát mutázy obsahuje Arg16, Arg157, Thr242, Glu246, Glu198, Asn194 a Lys168. The E-coli aktivní místo obsahuje stejné zbytky s výjimkou těchto známých výměn: Asp48 pro Asn194, Gln88 pro Glu248 a Ser84 pro Thr242. V aktivním místě enzymu interakce mezi těmito specifickými zbytky a substrátem omezují konformační stupně volnosti, takže entropie aktivace je účinně snížena na nulu, a tím podporuje katalýzu. Výsledkem je, že neexistuje žádný formální meziprodukt, ale spíše pseudo-diaxiální křeslo přechodový stav. Důkaz pro tuto konformaci poskytuje inverzní sekundární kinetický izotopový efekt na uhlíku přímo navázaném na hydroxylovou skupinu.[6] Tohoto zdánlivě nepříznivého uspořádání je dosaženo pomocí řady elektrostatických interakcí, které rotují prodloužený řetězec chorismátu do konformace požadované pro tento koordinovaný mechanismus.

Dalším stabilizujícím faktorem v tomto komplexu enzym-substrát je vodíková vazba mezi osamělým párem kyslíku ve vinyletherovém systému a zbytky donoru vodíkové vazby. To nejen stabilizuje komplex, ale narušení rezonance ve vinyletheru destabilizuje základní stav a snižuje energetickou bariéru pro tuto transformaci. Alternativní pohled je, že elektrostatická stabilizace polarizovaného přechodového stavu má v této reakci velký význam. V aktivním místě chorismát mutázy je analog přechodového stavu stabilizován 12 elektrostatickými a vodíkovými vazbami.[8] To je ukázáno u mutantů nativního enzymu, ve kterém je Arg90 nahrazen citrulin demonstrovat důležitost vodíkových vazeb ke stabilizaci přechodového stavu.[9] Další práce využívající chorismát mutázu z Bacillus subtilis prokázal, že když a kation byl vhodně umístěn do aktivního místa, elektrostatické interakce mezi ním a negativně nabitým přechodovým stavem podporovaly katalýzu.[2]
Byly provedeny další studie, aby se podpořila relevance konformátoru blízkého útoku (NAC) v reakci katalyzované chorismát mutázou. Tento NAC je reaktivní konformace základního stavu, která se v enzymu přímo převádí na přechodný stav. Použitím termodynamická integrace (TI), standardní volné energie (ΔGN°) pro tvorbu NAC byly vypočítány v šesti různých prostředích. Získaná data naznačují, že účinná katalýza je odvozena od stabilizace jak NAC, tak přechodového stavu.[10] Jiné experimentální důkazy však podporují, že pozorovaný efekt NAC je jednoduše výsledkem stabilizace elektrostatického přechodového stavu.[11][12]
Celkově byly provedeny rozsáhlé studie týkající se přesného mechanismu této reakce. Relativní příspěvek konformačního omezení pružného substrátu, specifická vodíková vazba k přechodnému stavu a elektrostatické interakce k pozorovanému zvýšení rychlosti je však stále předmětem diskuse.
Poznámky
- ^ Bylo navrženo, aby dimetylallyltryptofan syntáza katalyzovala a Vyrovnejte přeskupení, ale toto ještě musí být definitivně prokázáno[3]
Reference
- ^ A b C d Qamra R, Prakash P, Aruna B, Hasnain SE, Mande SC (červen 2006). „2.15 Krystalová struktura chorismát mutázy Mycobacterium tuberculosis chorismate mutase odhaluje neočekávané duplikace genů a naznačuje roli v interakcích hostitel-patogen“. Biochemie. 45 (23): 6997–7005. doi:10.1021 / bi0606445. PMID 16752890.
- ^ A b C Kast P, Grisostomi C, Chen IA, Li S, Krengel U, Xue Y, Hilvert D (listopad 2000). „Strategicky umístěný kation je zásadní pro účinnou katalýzu chorismát mutázou“. The Journal of Biological Chemistry. 275 (47): 36832–8. doi:10,1074 / jbc.M006351200. PMID 10960481.
- ^ Luk LY, Qian Q, Tanner ME (srpen 2011). „Vyrovnání reakce v reakci katalyzované dimethylallyltryptofan syntázou?“. Journal of the American Chemical Society. 133 (32): 12342–5. doi:10.1021 / ja2034969. PMID 21766851.
- ^ Selwood T, Jaffe EK (březen 2012). „Dynamické disociační homo-oligomery a řízení funkce proteinu“. Archivy biochemie a biofyziky. 519 (2): 131–43. doi:10.1016 / j.abb.2011.11.020. PMC 3298769. PMID 22182754.
- ^ Babu M (1999). „Anotace chorismát mutázy z Mycobacterium tuberculosis a Mycobacterium leprae genomu“ (PDF). Bakalářská práce pro Centrum biotechnologie.
- ^ A b Lee AY, Stewart JD, Clardy J, Ganem B (duben 1995). „Nový pohled na katalytický mechanismus chorismát mutáz ze strukturálních studií“. Chemie a biologie. 2 (4): 195–203. doi:10.1016/1074-5521(95)90269-4. PMID 9383421.
- ^ Gray JV, Knowles JR (srpen 1994). „Monofunkční chorismát mutáza z Bacillus subtilis: studie FTIR a mechanismus působení enzymu“. Biochemie. 33 (33): 9953–9. doi:10.1021 / bi00199a018. PMID 8061004.
- ^ Grisham C (2017). 6. vydání Biochemistry. Spojené státy americké: Brooks / Cole - Cengage Learning. p. 505. ISBN 978-1133106296.
- ^ Kienhöfer A, Kast P, Hilvert D (březen 2003). „Selektivní stabilizace přechodného stavu chorismát mutázy pozitivně nabitým dárcem vodíkové vazby“. Journal of the American Chemical Society. 125 (11): 3206–7. doi:10.1021 / ja0341992. PMID 12630863.
- ^ Hur S, Bruice TC (říjen 2003). „Konformační přístup blízký útoku ke studiu reakce chorismátu na prephenate“. Sborník Národní akademie věd Spojených států amerických. 100 (21): 12015–20. doi:10.1073 / pnas.1534873100. PMC 218705. PMID 14523243.
- ^ Strajbl M, Shurki A, Kato M, Warshel A (srpen 2003). „Zdánlivý efekt NAC v chorismát mutase odráží stabilizaci elektrostatického přechodového stavu“. Journal of the American Chemical Society. 125 (34): 10228–37. doi:10.1021 / ja0356481. PMID 12926945.
- ^ Burschowsky D, van Eerde A, Ökvist M, Kienhöfer A, Kast P, Hilvert D, Krengel U (prosinec 2014). „Stabilizace elektrostatickým přechodovým stavem spíše než destabilizace reaktantů poskytuje chemický základ pro účinnou katalýzu chorismát mutázy“. Sborník Národní akademie věd Spojených států amerických. 111 (49): 17516–21. Bibcode:2014PNAS..11117516B. doi:10.1073 / pnas.1408512111. PMC 4267393. PMID 25422475.